Cloning and molecular
characterization of heat shock
cognate 70 from tiger shrimp
(Penaeus monodon)
Wan-Yu Lo,
1Kuan-Fu Liu,
2I-Chiu Liao,
3Yen-Ling Song
4 1Institute of Zoology, National Taiwan University, Taipei, Taiwan2Tungkang Marine Laboratory, Taiwan Fisheries Research Institute, Tungkang, Taiwan 3Taiwan Fisheries Research Institute, Tungkang, Taiwan
4Department of Life Science, Institute of Zoology, National Taiwan University, Taipei, Taiwan
Abstract We cloned the complementary deoxyribonucleic acid (cDNA) of the heat shock cognate 70 (hsc70) gene of
tiger shrimp (Penaeus monodon). It was 2207 bp long and included a 1959-bp coding region, a 40-bp flanking region
at the 59end, and a 208-bp flanking region at the 39end. The deduced, 652–amino acid sequence had a molecular
mass of 71 481 Da and an estimated isoelectric point (pI) of 5.2. Based on phylogenetic analysis, the gene is clustered
with the hsc70 proteins of invertebrates and vertebrates. In native gel electrophoresis, recombinantP monodon hsc70
expressed in an Escherichia coli system is tightly associated with carboxymethylated a-lactalbumin (CMLA), which
indicates that hsc70 probably functions as a chaperone. In an in vitro adenosine triphosphatase assay, recombinant
hsc70 hydrolyzed adenosine triphosphate to adenosine-59-diphosphate and increased hydrolysis activity by binding to
unfolded peptide, CMLA. In situ hybridization using an antisense riboprobe revealed that the hsc70 gene was active
in most tissues of unstressed shrimp. The expression of hsc70 messenger ribonucleic acid (mRNA) in hemocytes
increased 2- to 3-fold at the first hour after shrimp experienced heat shock and 0.5-hour recovery. Hsc70 mRNA
decreased gradually to the background level. Cloning and characterizing theP monodon hsc70 gene is the first, crucial
step in studying the relationship of heat shock proteins with the stress or immune responses of shrimp.
INTRODUCTION
A sudden increase in temperature and other types of en-vironmental stresses induce the synthesis of a specific set of proteins called heat shock proteins (Hsps). The first description of a subset of cellular proteins in Drosophila induced by heat shock (Ritossa 1962) triggered extensive research on their function in stress tolerance in a variety of organisms. Hsps are grouped into 5 major families on the basis of their molecular size in kilodaltons (kDa): Hsp110, Hsp90, Hsp70, Hsp60, and the low–molecular weight Hsp family. The Hsp70 family occurs in diverse organisms, and it is highly conserved among eukaryotes. Received 16 April 2004; Revised 17 June 2004; Accepted 17 June 2004. Correspondence to: Yen-Ling Song, Tel: 33662455; Fax: 886-2-23660243; E-mail: [email protected].
Present address of I-C Liao: National Taiwan Ocean University, Keelung 202, Taiwan
Members of the Hsp70 family play an essential role in protein metabolism in both stressed and unstressed or-ganisms. Hsp70 proteins are involved with de novo pro-tein folding, membrane translocation, formation and dis-assembly of protein complexes, and degradation of mis-folded proteins (Hartl 1996; Bukau and Horwich 1998; Kregel 2002).
Hsp70 family members are composed of 3 domains. The 44-kDa, N-terminal adenosine triphosphatase (ATPase) domain binds and hydrolyses adenosine tri-phosphate (ATP). The variable 18-kDa, peptide-binding domain interacts with unfolded polypeptides, and the 10-kDa, C-terminal domain bears the highly conserved, EEVD terminal sequence present in all eukaryotic Hsp70 families (Flaherty et al 1990; Kiang and Tsokos 1998). Hsp70 family members also interact with a number of other proteins, promoting specific chaperoning functions (Santacruz et al 1997). Some hsp70 proteins are weakly
expressed, at best, under normal conditions but are in-duced by heat and other stresses allowing cells to cope with acute stressor insults. The genes for these bona fide hsp70 proteins lack introns. The heat shock cognate 70 (hsc70) proteins are expressed constitutively. They play essential roles in protein metabolism under normal con-ditions. The genes for these proteins contain introns and are only slightly inducible, at best. Physiologically, hsc70 functions as a ‘‘housekeeping protein’’ or ‘‘professional chaperone.’’ It is involved with many cellular physiolog-ical processes. They include the folding of newly synthe-sized polypeptides, membrane translocation, formation and disassembly of protein complexes, and degradation of misfolded proteins (Hartl 1996; Bukau and Horwich 1998; Kiang and Tsokos 1998; Hartl and Hayar-Hartl 2002; Kregel 2002). Hsc70 even increases cell survival by restoring cellular homeostasis (Mallouk et al 1999). Ad-ditional members of the Hsp70 family include a mito-chondrial form, hsp75, and a glucose-regulated protein known as Grp78, or heavy-chain binding proteins (Bip), that is located in the endoplasmic reticulum (Lindquist and Craig 1988; Kiang and Tsokos 1998).
The Hsc70 protein always associates with nascent poly-peptide chains. It also forms a stable complex with un-folded proteins, such as reduced carboxymethylated a-lactalbumin (CMLA), a thermally unstable mutant of staphylococcal nuclease and apocytochrome c (Beckmann et al 1990; Palleros et al 1991; Sadis and Hightower 1992; Frydman et al 1994). Hsc70 has a high affinity for ATP but relatively weak intrinsic ATPase activity. The ATP hy-drolysis activity of hsc70 can be stimulated by the other peptides, suggesting that the released energy is used to facilitate folding of the substrates. Thus, CMLA is used as a model protein to form a complex with hsc70 to iden-tify the binding capacity in basal chaperone functions (Wang and Lee 1993; Hu and Wang 1996).
Penaeus monodon is native to the Western Pacific region and is a major species in Asian aquaculture. As previ-ously reported, 2 expressed sequence tag (EST) homo-logues for the stress-related hsp70 gene were found in tiger shrimp (P monodon) and Pacific white shrimp (Li-popenaeus vannamei) (Tassanakajorn A, GenBank: BI78445; Gross et al 2001) and an EST homologue for the stress-related hsp60 gene was found in P monodon (Lehnert et al 1999). However, they were not characterized further, and their full-length complementary deoxyribonucleic acids (cDNA) were not obtained. In this study, we molecularly cloned and characterized the hsc70 gene in P monodon. The recombinant protein expressed in an Escherichia coli system is the first member of the Hsp70 family in shrimp. Using in situ hybridization, we compared the expression patterns of hsc70 in a variety of tissues from unstressed shrimp. Finally, we demonstrated that regulation of hsc70 expression in P monodon hemocytes was heat dependent.
MATERIALS AND METHODS
Hemocyte collection and ribonucleic acid extraction Shrimp hemolymph was withdrawn from the ventral si-nus in the first abdominal segment using a 26-gauge hy-podermic needle on a 1-mL syringe. The syringe was pre-filled with anticoagulant solution (100 mM sodium cit-rate, 250 mM sucrose, 10 mM Tris-HCl, pH 7.6, 7806 20 mOsm/kg). Extra anticoagulant was added to make a 1: 4 volume ratio of hemolymph to anticoagulant. The mix-ture was centrifuged at 5003 g for 10 minutes to separate hemocytes from plasma. Hemocytes (106 cells) were
im-mediately homogenized in 1 mL TRIzol reagent (Gibco BRL, Carlsbad, CA, USA). The total ribonucleic acid (RNA) was extracted and further purified by using an RNA isolation kit according to the manufacturer’s instruc-tions (Gibco BRL).
Reverse transcription–polymerase chain reaction First-strand cDNA synthesis was carried out using 2 mg of deoxyribonuclease-treated total RNA as template and incubating at 378C for 30 minutes. Reverse transcription was performed using Moloney leukemia virus reverse transcriptase (Super-Script Preamplification System, Gib-co BRL). Degenerate oligonucleotide primers, 59 GGGAT GGTCG GCAA(A/G) TGGCA (T/C)TTGG 39 and 59 AAGTG AGCCT TGTAC TT(C/G)CC 39, were synthe-sized on the basis of the polynucleotide sequences of hsc70 from oyster (AF14466), coral (AF152004), and sea urchin (X61379). A 330-bp fragment was cloned using a set of these primers and 1 mg of hemocyte cDNA as a template. To this mixture we added 5 mL polymerase chain reaction (PCR) 103 buffer 1 Mg21, 4 mL
deox-ynucleoside triphosphate MIX, and 1mL Taq DNA poly-merase (500 U/mL) KIT (Viogene, Taipei, Taiwan) ac-cording to the manufacturer’s instructions, yielding a fi-nal volume of 50mL. Thirty cycles of PCR amplification were performed. Each cycle consisted of denaturation for 40 seconds at 948C, 45 seconds of annealing at 568C, and 45 seconds of extension at 728C. The last extension step was extended to 10 minutes at 728C, followed by cooling to 48C. Amplified DNA fragments were ligated to the pGEM-T Easy vector (Promega, Madison, WI, USA) and transfected into E coli DH5a–competent cells. DNA se-quencing, from both ends of 3 clones, was carried out using an ABI PRISM BigDye Terminator Cycle Sequenc-ing Ready Reaction kit (Applied Biosystems, Foster City, CA, USA) with a DNA sequencer (ABI PRISM 377-96, Applied Biosystems).
Rapid amplification of cDNA ends
Full-length cDNA was obtained by using both 59–rapid amplification of cDNA ends (RACE) and 39-RACE. The
first-strand cDNA used in 59-RACE was synthesized as described above. The 59 end of the cDNA was tailed using a terminal deoxynucleotidyl transferase (Boehringer Mannheim) and deoxyguanosine triphosphate. The re-sulting poly G–tailed cDNA was used to generate a dou-ble-stranded cDNA in the first PCR amplification in the presence of a universal primer, RAAPC (59 GGCCA CGCGT CGACT AGTAC T(C)939), and a P
monodon–spe-cific primer, R1 (59 GACAA GGACG ATGTC GTGGA TC 39). PCR was carried out as described above, except that the annealing temperature was 568C. The first PCR prod-uct was diluted 50-fold with sterile, nanopure water. Then, 1mL diluted PCR product was used as the template for the second PCR amplification (at 608C), which used another universal primer, RAUAP (59 GGCCA CGCGT CGACT AGTAC 39) and a specific R2 (59 CATCA CGGAG TGACT TCTCC 39). Procedures for 39-RACE were the same as for 59-RACE, except that the first-strand cDNA was synthesized using a specific F1 (59 GATCC ACGAC ATCGT CCTTG TC 39) and an universal RAAPT (59 GGCCA CGCGT CGACT AGTAC (T)1839) to generate a
double-stranded cDNA, and the temperature was 328C. For the nested PCR (at 568C), 1 mL of the first PCR prod-uct (diluted 100-fold) was used as a template in the pres-ence of a pair of primers: F2 (59 GCCGG CGGTG TGATG ACTGC G 39) and RAUAP. Amplified DNA fragments were subcloned, and 3 clones from each end were se-quenced.
Sequence analysis
Nucleotides and deduced amino acid sequences were submitted to the National Center of Biotechnology Inter-national (NCBI, Bethesda, MD, USA) database for BLASTN and BLASTP searches of matches to known GenBank sequences. Multiple alignments were made with the ClustalW of Data Analysis in Molecular Biology and Evolution (DAMBE, version 4.0.75) software package (Xia 2000; Xia and Xie 2001). Using the alignment of ami-no acid sequences from different species, phylogenetic and molecular evolutionary analyses were conducted us-ing MEGA version 2.1 (Kumar et al 2001). The phyloge-netic tree was constructed using the neighbor-joining method (Nei and Kumar 2000).
Production of recombinant hsc70
To clone the hsc70 gene into the transfer vector, flanking NdeI and HindIII restriction sites (in bold) were intro-duced at the 59 and 39 ends by PCR mutagenesis with a sense primer (59-CATAT GGCAA AGCCT-39) and an an-tisense primer (59-AAGCT TTAAT CGACT TCCTC-39). The new hsc70 cDNA fragment was cloned into the pET-22b (1) transfer vector (Novagen, Darmstadt, Germany),
and then transformed into E coli BL21-CodonPlus (DE3)-RP-X–competent cells (Stratagene, La Jolla, CA, USA) ac-cording to the manufacturer’s instructions. The transfor-mation mixture was incubated with 600 mL Luria-Bertani medium containing 100 mg/mL ampicillin and 50 mg/ mL chloramphenicol for 16 to 20 hours at 378C. After the culture reached an optical density of OD600 5 0.4, re-combinant protein expression was induced with isopro-pylb-thiogalactoside, at a final concentration of 0.4 mM. Incubation was continued for four more hours.
Purification of recombinantP monodon hsc70 protein and the N-terminus sequence
The induced bacterial culture was centrifuged at 50003 g for 10 minutes, and the pellet was collected and stored overnight at2208C. The E coli pellet was resuspended in 20 mL B-PER Bacterial Protein Extraction Reagent (Pierce, Rockford, IL, USA) by vortexing until the cell suspension was homogeneous. Purified inclusion bodies were dis-solved with the same reagent according to the manufac-turer’s instructions. The inclusion bodies were resuspend-ed in 10 mL of 2 M urea in buffer A (5 mM MgCl, 125 mM NaCl, and 25 mM Tris base, pH 7.0) and incubated on ice for 10 minutes. After centrifugation at 12 0003 g for 10 minutes, the supernatant was stored at 48C. These procedures were repeated using 4 and 6 M urea, and each supernatant was collected and pooled together. We added 1 M L-arginine (Sigma) to the supernatant and kept the
mixture at 48C for 10–15 minutes to renature the unfolded or misfolded recombinant protein. After centrifugation at 12 0003 g for 20 minutes, the supernatant was dialyzed in 5 L 0.01 M phosphate-buffered saline (PBS) overnight to remove unnecessary urea. The dialyzed mixture was centrifuged at 12 000 3 g for 20 minutes. Then, 10 mM MgCl2was added to the supernatant before it was
sepa-rated with a 2-mL adenosine-triphosphate agarose col-umn (Sigma). Elution of the recombinant hsc70 was per-formed as previously described (Wang and Lee 1993). Af-ter collection, the fractions were dialyzed in 0.01 M PBS. The protein solution was concentrated with an Amicon Ultra-4 filter (Millipore, Billerica, MA, USA) and stored at2808C. Finally, the N-terminal amino acid sequence of the recombinant hsc70 was determined by Edman deg-radation using a Procisey 490 protein sequencing system (Applied Biosystems).
Formation of the recombinant hsc70–CMLA complex
To determine whether a hsc70-CMLA complex formed, 3 mM purified recombinant P monodon hsc70 was mixed with 25 mM CMLA (excess) and 0.2 mM adenosine-59-diphosphate in the buffer (75 mM KCl, 40 mM N-2-hy-droxyethylpiperazine-N9-2-ethanesulfonic acid [HEPES],
10 mM dithiothreitol, and 4.5 mM Mg(CH3COO)2, pH
7.0) to achieve a final volume of 20mL. The mixture was incubated at 378C for 45 minutes (Hu and Wang 1996). At the end of incubation, 5 mL of sample buffer (200 mM Tris, 25 mM ethylenediaminetetraacetic acid [EDTA], 0.05% bromophenol blue, 50% glycerol, pH 7.0) were add-ed. Finally, 12.5% native gel electrophoresis was conduct-ed as describconduct-ed by Laemmli (1970), except that sodium dodecyl sulfate (SDS) was omitted from the gels and the running buffer. The gels were stained with Coomassie Brilliant Blue.
ATPase assay
An ATP-hydrolysis assay was conducted according to the procedures of Tsai and Wang (1994). Briefly, 3mg recom-binant P monodon hsc70 protein was incubated at 378C in 10 mL of buffer B (75 mM KCl, 40 mM HEPES, 4.5 mM Mg[CH3COO]2, pH 7.0) with, or without, 50mM CMLA
(Sigma). Then, 1mCi [g-32P]-ATP (3000 Ci/mmol; NEN/
Dupont, Boston, MA, USA) was added. At 0, 20, 40, and 60 minutes, 0.25 mL of the reaction mixture was with-drawn and spotted onto PEI-cellulose sheets (Millipore). The chromatograms were developed in 1.0 M formic acid–0.5 M LiCl. The amount of ATP hydrolyzed was quantified by PhosphoImager Analysis (Molecular Dy-namics, Piscataway, NJ, USA).
Tissue preparation for histology
Juvenile shrimp (3–5.8 g) were fixed in an RNA-friendly fixative containing 34.9% formalin, 40.7% ethanol, and 2.2% ammonium hydroxide (pH 6.5) (Hasson et al 1997). After dehydration, the samples were embedded in Para-plast, and 4-mm sections were cut and mounted on slides and stored at 48C.
In situ nucleic acid hybridization of hsc70 messenger RNA in tissue sections
Riboprobes
A GEM-T Easy plasmid containing a region (1856–2175 nt) of P monodon hsc70 cDNA was used as a template for the preparation of the probes. Digoxigenin (DIG)-uridine triphosphate–labeled sense and antisense riboprobes were generated from linearized cDNA plasmids (5mg) by in vitro transcription using the RNA-labeling kits T7 RNA polymerase and SP6 RNA polymerase (Boehringer Mannheim), respectively.
Hybridization
After deparaffinization and hydration, sections were treated with proteinase K (20 mg/mL) for 30 minutes at
378C. The sections then were acetylated for 10 minutes at room temperature (using 0.25% acetic acid and 0.1 M tri-ethanolamine, pH 8.0) and dehydrated in ethanol. Sec-tions were incubated overnight at 438C with hybridization solution containing 2.5 ng/slide of DIG-labeled hsc70 an-tisense probe. The hybridization solution consisted of 43 standard saline citrate (SSC), 50% formamide, 13 Den-hardt’s solution, 5% dextran sulfate, 0.5 mg/mL of salm-on sperm DNA, and 0.25 mg/mL of yeast transfer RNA. The sections were washed at 438C with 13 SSC for 30 minutes and then with 0.13 SSC for 15 minutes. The fol-lowing steps were performed according to instructions for using Boehringer’s DIG kit in alkaline phosphatase (AP) substrate for 1.5 hours at room temperature. The enzyme reaction was stopped by adding 100% methanol for 10 minutes. Then, the sections were counterstained with Bismarck Brown Y (Sigma) for 30 seconds. Finally, the sections were dehydrated, made transparent with graded ethanol and xylene, and mounted in GEL/ MOUNTy (Biømeda Corp, Foster City, CA, USA). The sections were examined with a light microscope to de-termine the density of fixed, AP enzymatic reaction prod-ucts (blue-black), and the signal density of each tissue was recorded as faintly positive (1), moderately positive (11), or very strongly positive (111).
Northern blot analysis of hsc70 messenger RNA in P monodon hemocytes
Experimental animals
Healthy looking tiger shrimp (body length, 17.86 1.5 cm; weight, 28.46 1.64 g) were collected from the Tungkang Marine Laboratory located in southern Taiwan. Shrimp were kept in a plastic tank supplied with 1000 L of 266 18C, constantly flowing, recirculating seawater with 2% salinity. Shrimp were stocked at a density of 20–25 indi-viduals per cubic meter. Twice a day, shrimp were fed commercial feed pellets equivalent to 5% of their body weight. Shrimp were conditioned for more than 1 month before receiving heat treatments.
Shrimp were transferred from water maintained at a controlled temperature of 266 18C to a tank containing seawater preheated to 376 18C. After 1 hour at 37 6 18C, the temperature of the water was rapidly decreased to 26 6 18C for 0.5 hours of recovery by draining and adding cool water (266 18C, flow rate: 30 L/min). Shrimp in the control group were transferred to another tank of water maintained at 268C and kept at the same temperature by draining and adding water as in the test group. It was noticed that the water volume always was kept the same, both in heat stress and recovery treatment in the 2 groups. Hemolymph was withdrawn 0, 1, 2, 4, and 6 hours after treatment. For each sample, 10 shrimp were
randomly selected from the experimental and control groups. With 5 people, hemolymph was withdrawn si-multaneously from the first 5 shrimp of each sample and then, immediately afterward, from the remaining 5 shrimp.
DNA probes
A shrimp-specific hsc70 cDNA probe was prepared using PCR. Two primers were synthesized, F5 (59-ATGGC AAAGG CACCT GCTGT CGG-39) and R1. The primers corresponded to both ends of a 1012-bp fragment (nt 41– 1052). This fragment contained 2 signature motifs, the adenosine triphosphate-guanosine triphosphate (ATP-GTP)–binding site, potential bipartite nuclear localization signals, and the nonorganellar eukaryotic consensus re-gion. A PCR DIG-labeled DNA probe synthesis kit (Boeh-ringer Mannheim) was used to produce the DNA probe. The unincorporated DIG-labeled nucleotides were re-moved using a Nick column (Pharmacia, New York, NY, USA). The resulting 537-bp hemocytic actin DIG probe was synthesized as the hsc70 probe. Briefly, an oligo(dT)-primed cDNA corresponding to P monodon hemocytic ac-tin was used as a template. The primers 59ACGAG GGCTA CGCCC TGCCC C39 and 59GAGGC CAGGA TGGAG CCGCC G39 were synthesized on the basis of the EST of tiger shrimp hemocytic actin (AW600757).
Northern blot
From each sample, 10 mg of total RNA was run electro-phoretically on a denaturing formaldehyde-agarose gel (1.2%) in 13 formaldehyde gel-running buffer (20 mM 3[N-morpholino] propanesulfonic acid, 8 mM sodium ac-etate, and 1 mM EDTA, pH 7.0) at 50 V for 1 hour. Sam-ples were compared with a RNA Millennium Marker (Ambion, Austin, TX, USA), which served as the control. Overnight, separated RNA was transferred with 203 SSC onto a nylon membrane (Millipore) by capillary blotting. The blotted membrane was cross-linked with 20 000 mJ by an UV-linker (Stratagene) before being subjected to hybridization. Each membrane was prehybridized for 2 hours at 608C in hybridization solution (50% formamide, 53 SSC, 2% blocking reagent, 0.1% N-lauroylsarcosine, and 0.02% SDS). Hybridization was performed overnight at 608C in buffer containing the hsc70-specific probe. Af-ter hybridization, the membranes were stripped with pre-warmed dimethylformamide and reprobed with the actin cDNA probe. A total of 4 washes were performed at room temperature for 5 minutes. The first 2 washes were in 80 mL 23 SSC–0.1% SDS, and the final 2 washes were in 100 mL 13 SSC–0.1% SDS at 508C for 15 minutes each. Detection was performed with a DIG Nucleic Acid
De-tection Kit (Boehringer Mannheim). Blots were quantified using a Personal Densitometer S1 (Molecular Dynamics), and the hsc70 transcript was normalized relative to the actin transcript.
RESULTS
Isolation of cDNA clones, sequencing and analysis of the deduced amino acid sequences
The full-length cDNA of P monodon heat shock hsc70 pro-tein (GenBank accession number AF474375) was cloned. It is composed of 2207 bp, including a 1959-bp coding region and 40- and 208-bp flanking regions at the 59 and 39 ends, respectively. The most probable translation ini-tiation codon, ‘‘ATG,’’ is located at nt 41 from the 59 end, and the stop codon, ‘‘TAA,’’ is at nt 1997. A single poly-adenylation site (AATAAA) was found at nt 2158–2163 (Fig 1). The guanine-cytosine content of the P monodon hsc70 gene was 0.566. The deduced amino acid sequence encoded a 652–amino acid protein with a calculated mo-lecular mass of 71 481 Da and an isoelectric point (pI) of 5.21. Several general eukaryotic Hsp70 family motifs were identified on the deduced amino acid sequence: 2 signa-tures (IDLGTTYS, aa 9–16; DLGGGTFD, aa 199–206) and a putative ATP-GTP–binding site (AEAYLGST; aa 131– 138). Two additional specific motifs were found: the non-organellar consensus motif (RARFEEL; aa 299–305) and the cytoplasmic Hsp70 carboxyl terminal region (GPTIEEVD; aa 649–652) (Fig 1).
Relationship with other members of the Hsp70 family
A phylogenetic tree was constructed by analyzing the amino acid sequences of P monodon hsc70 and the hsc70, hsp70, and Bip from a variety of eukaryotic species. Twenty-three Hsp70 family members from 10 diverse genera were selected for analysis. P monodon hsc70 clus-tered with Artemia hsp70 on a unique branch of the tree. They clustered with other hsc70 from vertebrates, mol-luscs, and insects. The amino acid sequences of human, rat, Xenopus, trout, zebrafish, oyster, fly, chironomus, and P monodon hsc70 shared 83–99% identity. Human, rat, mouse, bovine, tilapia, and flounder hsp70 amino acid sequences shared 88–99% identity. The Bip and prokary-otic E coli DnaK sequences form their own unique branch-es (Fig 2).
RecombinantP monodon hsc70
In this study, recombinant hsc70 was overproduced as shown in the whole-cell lysate (Fig 3, lane 1). Almost all overexpressed proteins are insoluble and form inclusion bodies (Fig 3, lane 2). After the ATP-agarose column step,
Fig 1. Nucleotide and deduced amino acid sequence ofPenaeus monodon hsc70 complementary deoxyribonucleic acid (cDNA). The nucleotide sequence of the gene encoding the hsc70 protein and the predicted amino acid sequence deduced from the cDNA inserts are shown. Two signatures are underlined with dots, and the putative adenosine triphosphate-guanosine triphosphate (ATP-GTP)–binding site is boxed. The potential nonorganellar eukaryotic consensus motif is underlined, and the cytoplasmic motif carboxyl terminal region (GPTIEEVD) is in bold. The polyadenylation signal (AATAAA) is shown in a gray box, and the stop codon is marked with an asterisk.
essentially homogeneous hsc70 was collected from frac-tions 1–8 (Fig 3, lanes 3–10). Recombinant hsc70 migrates in the gel as a 71.5-kDa protein. Amino acid sequence analysis found that the N-terminus of recombinant hsc70 is AKAPA. This matches the corresponding, deduced P monodon hsc70 N-terminal amino acid sequence and is different from that of the E coli, 70-kDa hsp, DnaK.
Formation of recombinantP monodon hsc70–CMLA complex
To assay for a tightly associated, recombinant P monodon hsc70–CMLA complex, the protein was incubated with CMLA and then resolved on native gel. Polyacrylamide gel electrophoresis (PAGE) shows that the top ‘‘two
Fig 2. Phylogenetic tree showing the relationship ofPenaeus monodon hsc70 amino acid sequence to other Hsp70 family members from a variety of spe-cies. Sequences used in the tree, fol-lowed by their GenBank accession num-ber, wereXenopus hsc70I (AAB97092), Xenopus hsc70II (AAB41583), zebra-fish hsc70 (AAB03704), trout hsc70 (AAB21658), human hsc70 (CAA68445), rat hsc70 (CAA68265), scallop hsc70 (AAS17724), oyster Cghsp70 (CAC83009), oyster Cghsc70 (CAC-83683),P monodon hsc70 (AAQ05768), Artemia hsp70 (AAL27404), fly (AAC-23392), chironomus Cthsc70 14525), chironomus Cyhsc70 (AAN-14426), flounder hsp70 (BAA31697), ti-lapia hsp70 (CAA04673), mouse hsp70 (AAA59362), rat hsp70 (AAA17441), human hsp70B (NP005337), rat heavy-chain binding proteins (Bip) (AAA40817), yeast Bip (AAA08536), and Escherich-ia coli DnaK (BAB96589). The distance is the proportion of amino acid sites at which 2 sequences are different. Boot-strap values are given in percent.
Fig 3. Purification of recombinant hsc70. In sodium dodecyl sul-fate–polyacrylamide gel electrophoresis (SDS-PAGE) (12.5% acryl-amide), each lane corresponds to a different purification step. M, molecular mass markers (in kDa, on left); lane 1, whole-cell lysate; lane 2, inclusion body lysate; lanes 3 to 10, fractions 1 to 8 were eluted from an adenosine triphosphate (ATP)–agarose column. Re-combinantPenaeus monodon hsc70 migrates in the gel as a 71.5-kDa protein (arrow).
bands’’ in Figure 4, lane 1 emerge when hsc70 was mixed with CMLA (upper band was free hsc70 and the lower band was hsc70-CMLA complex). The other excess CMLA with its degraded proteins that cannot complex with recombinant hsc70 will be observed on the bottom band in lane 1 (Fig 4). Recombinant hsc70 only is in lane 2. ATPase assay
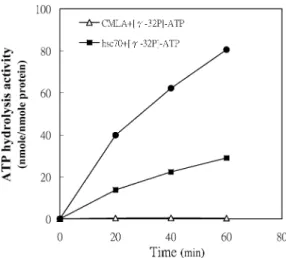
The hydrolysis activity of ATP by recombinant P monodon hsc70 increased with incubation time and then formed a complex with CMLA. CMLA increased recombinant hsc70 ATP hydrolysis activity (Fig 5, top). The activity of recombinant hsc70, alone, was low and that of CMLA alone was very low (Fig 5, middle and bottom). After incubation for 60 minutes with 50mM CMLA, hsc70 ATP hydrolysis activity was about 3.2-fold higher than that of hsc70 alone.
In situ nucleic acid hybridization
To determine the distribution of the hsc70 gene expression in different tissues, we used in situ nucleic acid
hybrid-Fig 4. Identification of the hsc70–carboxymethylateda-lactalbumin (CMLA) complex. Recombinant Penaeus monodon hsc70 (3mM) was incubated with 50mM CMLA and then separated by native gel electrophoresis. In the resolving gels, the concentration of acryl-amide is 12.5%. Lane 1 has a mixture of CMLA and hsc70. The arrow points at the upper one of the top 2 bands is free hsc70 not yet bound to CMLA. The lower band is the tightly bound hsc70-CMLA complex (arrow with dots). The bottom band is excess hsc70-CMLA and it’s degraded products. Lane 2 has recombinant hsc70 only.
Fig 5. The effect of hsc70 and CMLA on adenosine triphosphate (ATP) hydrolysis. Recombinant hsc70 (3mg) was incubated with or without 50mM CMLA. To initiate the reaction, 0.2 mM [g-32P]-ATP was added to the mixtures, in a total volume of 10mL. Every 20 minutes, ATP hydrolysis was measured. CMLA alone was incapable of ATP hydrolysis (open triangles).
ization to assess the basal levels of expression in un-stressed tiger shrimp. A blue-black, positive signal was observed in nearly all shrimp tissues and organs, except for the epithelium (Fig 6B, bold arrow). Unexpectedly, the largest shrimp organ, the hepatopancreas, exhibited only a faint signal (Fig 6A, arrow). The strongest positive sig-nals for hsc70 messenger RNA (mRNA) occurred in the gills, heart, hematopoietic tissue, hemocytes, and the lym-phoid organ. Moderately positive signals for hsc70 mRNA were observed in the eyestalk, hindgut, midgut, and stomach (Table 1).
Northern blot analysis
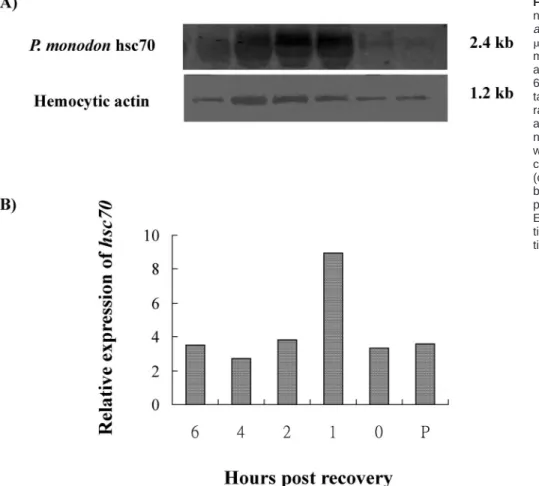
To determine whether P monodon constitutive hsc70 was inducible, shrimp were also subjected to heat shock treat-ment. Hemocytes, the cells in which the gene was isolat-ed, were used in this analysis. In tiger shrimp, heat shock treatment increased the amount of hsc70 mRNA tran-script. A 2.4-kb band was found in shrimp hemocytes before treatment as well as 0, 1, 2, 4, and 6 hours after recovery. The signal was weak before heat treatment and after 24 hours (Fig 7, lane P and 0). Expression was great-est during the first hour after heat treatment and recov-ery. It gradually decreased to the pretreatment level at the next time point. Hsc70 was expressed constitutively and until the termination of the experiment (Fig 3). In the control group, no notable difference was found at each sampling time (data not shown).
DISCUSSION
A full-length cDNA, containing a 1959-bp coding region with 40- and 208-bp flanking regions at each end, was cloned. The GPKH motif characteristic of the prokaryotic Hsp70 family was not present between amino acids 295– 296, excluding the possibility of false prokaryotic cloning (Karlin and Brocchieri 1998). The deduced amino acid se-quence of shrimp hsc70 was highly homologous with the amino acid sequences of Artemia, rainbow trout, humans, rats, and zebrafish hsc70. Two signature (I/V-D-L-G-T-T-x-S and D-L/F-G-G-G-T-F-D) sequences and the ATP-GTP–binding motif functional domain (A-E-A-Y-L-G-K/ R-T) characteristic of the eukaryotic Hsp70 family were found in the shrimp hsc70 sequence. Thus, the gene in this study clearly belongs to the eukaryotic Hsp70 family. Putative bipartite nuclear localization signals (K-K and R-R-L-R-T) were identified at aa 250–251 and aa 261–265 of the P monodon hsc70 primary sequence. The signal, char-acterized by an abundance of the basic amino acids lysine and arginine, is needed for the selective translocation of hsc70 protein into the nucleus (Knowlton and Salfity 1996). In addition, P monodon hsc70 included a nonorga-nellar stress protein motif (R-A-R-F-E-E-L; aa 299–305),
Fig 6. In situ hybridization of shrimp hsc70 with antisense riboprobe. (A) Lymphoid organ and hepatopancreas (2003). (B) Heart (2003) ofPenaeus monodon. The sections are slightly counterstained with Bismarck Brown Y. In (A), the red arrow points at the he-patopancreas, and in (B), the black ar-row points at the epithelium. No posi-tive signals were observed when a sense hsc70 riboprobe was used (data not shown).
Table 1 Abundance ofhsc70 mRNA in Penaeus monodon tissues as determined by in situ nucleic acid hybridizationa
Tissue Signal intensity
Eyestalk Gill Heart Hematopoietic tissue Hemocytes Hepatopancreas 11 111 111 111 111 1 Hindgut Lymphoid organ Midgut Muscle Stomach 11 111 11 1 11
amRNA, messenger ribonucleic acid; 111, strong positive sig-nal;11, moderate positive signal; and1, faint positive signal.
tetrapeptide GGXP repeats (where X is any aliphatic res-idue; aa 615–638), and the cytosolic-cytoplasmic Hsp70 family carboxyl terminal region (G-P-T-I/V-E-E-V-D-stop codon; aa 595–652), which affects binding of certain co-factors of the hsc70 protein (Demand et al 1998). These motifs account for the localization of our hsc70 in the cell cytosol and cytoplasm (Boorstein et al 1994; Gupta and Singh 1994; Rensing and Maier 1994). Based on the se-quence analyses presented above, the P monodon hsc70 is a member of the eukaryotic cytosolic-cytoplasmic Hsp70 family (Demand et al 1998; Vayssier et al 1999).
P monodon hsc70 clustered with the hsc70 amino acid sequences of vertebrates, arthopods, and molluscs to form a distinct clade. P monodon hsc70 is not as closely related to other eukaryotic hsp70 (whose genes lack introns), Bip in the Hsp70 family, and prokaryotic hsp70-equivalent DnaK. Phylogenetically, P monodon hsc70 is most closely related to brine shrimp hsp70 (Artemia franciscana). Both belong to a group of eukaryotic hsc70 whose genes
con-tain introns (Fig 2). Unfortunately, the brine shrimp hsp70 amino acid sequence in GenBank is incomplete in the C-terminal region. Therefore, our comparison of P monodon and A franciscana sequences is preliminary and inconclu-sive. However, the N-terminal 644 amino acids of P mon-odon hsc70 and Artemia hsp70 exhibited 86% identity (data not shown). Typically, eukaryotic Hsp70 family members localized in the cytoplasm share approximately 50% identity with prokaryotic hsp70. Among eukaryotic subgroups, cytoplasmic members share at least 71% iden-tity (Boorstein 1994). A detailed comparison of the amino acid sequence of P monodon hsc70 with the sequences of the other Hsp70 family members currently available in the database found highly conserved regions in the N-terminal part of the protein. Conservation of the amino acid sequence is expected decrease toward the C-terminal end of the protein.
Eukaryotic recombinant hsc70 produced by the E coli expression system usually is in the soluble form (Wang and Lee 1993; Boutet et al 2003). However, recombinant P monodon hsc70 was present in an insoluble form (inclu-sion bodies). Hsc70 protein (0.4 to 0.6 mg) was obtained from 0.6 L of cell culture. Insoluble recombinant hsc70 required a longer extraction time than the soluble hsc70, so that it sometimes was degraded from 71.5-kDa to 44-and 27.5-kDa fragments when separated by SDS-PAGE gel (Fig 3, lanes 5–7). Therefore, for biochemical assays and antibody production, we always purified the com-plete recombinant hsc70 by dialyzing with a membrane tube (MWCO: 5000). The five N-terminal amino acids were sequenced. They were A-K-A-P-A. They matched the deduced P monodon hsc70 sequence and proved that the sequence was eukaryotic. The N-terminal amino acids for the E coli sequence are G-K-I-I-G. The recombinant P mon-odon hsc70 was produced by a prokaryotic (E coli)
ex-Fig 7. Northern blot analysis of ribo-nucleic acid (RNA) extracted from Pen-aeus monodon hemocytes. RNA (10
mg per lane) was isolated from the he-mocytes of pretreated shrimp (lane P) and heat shocked shrimp 0, 1, 2, 4, and 6 hours after 0.5 hours of recovery. To-tal RNA was electrophoretically sepa-rated on a denaturing formaldehyde agarose gel (1.2%) and blotted onto a nylon membrane. (A) The upper blot was hybridized with thehsc70-specific complementary deoxyribonucleic acid (cDNA) probe. The lower blot was hy-bridized with a hemocytic actin–specific probe, providing an internal control. (B) Expression ofP monodon hsc70 rela-tive to expression of the hemocytic ac-tin gene.
pression system. Thus, the first amino acid (methionine) of the protein is not translated (Wang and Lee 1993).
Using native gel electrophoresis, we verified that re-combinant P monodon hsc70 formed a tight complex with CMLA. The two closest bands were unbound hsc70 (up-per) and hsc70-CMLA complex (lower) (Fig 4, top 2 bands in lane 1). The data demonstrated that the mobility of the tightly associated hsc70-CMLA complex is greater than that of hsc70 alone as given in a previous report (Hu and Wang 1996). In addition, we showed that recombi-nant hsc70 ATP hydrolytic activity was stimulated by the unfolded peptide, CMLA. This provides further evidence that recombinant P monodon hsc70 protein functions as a chaperone (Gething and Sambrook 1992; Benaroudj et al 1996; Hu and Wang 1996; Takeda and McKay 1996; Leng et al 1998; Kampinga et al 2003).
Using an in situ hybridization assay, we found that hsc70 mRNA transcripts were present in all P monodon tissues, except the epithelium. The level of hsc70 expres-sion varied in different tissues of P monodon (Table 1). Differential expression of hsc70 also has been reported in the tissues of rats, Xenopus, and carp (O’Malley et al 1985; Ali et al 1996, 2003). The exact reasons and mechanisms for this differential expression are not well understood.
In this study, we demonstrated that hsc70 increased
significantly in the hemocytes of P monodon after heat shock treatment (Fig 7). In previous studies, heat shock treatment did not increase hsc70 expression in most spe-cies (Kiang and Tsokos 1998). However, an increasing number of studies have demonstrated that heat shock treatment does increase mRNA levels severalfold ( ,20-fold) in a variety of cell types, including oyster hemocytes (Gourdon et al 2000), human peripheral blood monocytes (Jacquier-Sarlin et al 1995), human lymphocytes (Hansen et al 1991), and human HeLa cells (O’Malley et al 1985). Similarly, expressions of several members of the multi-genic Hsp70 family that are expressed constitutively in unstressed animals increase severalfold in stressed ani-mals, indicating that they are inducible (Taviara et al 1996). Based on our results and those from other studies, shrimp hemocytes, which are similar to immune cells, provide vital cellular protection against stress.
Cloning and characterization of the hsc70 and other stress protein genes in P monodon provides a tool for in-vestigating the immune and stress-related responses in shrimp. We are now in the process of cloning and se-quencing the hsp70 (the major inducible form). Based on differences between hsp70 and hsc70, we can develop the transgenic techniques in marine crustaceans. In addition, the conserved amino acid sequences of these stress
pro-teins provide invaluable information about the evolution of the immune system in the shrimp and other inverte-brates.
ACKNOWLEDGMENTS
We sincerely thank Dr C Wang, Institute of Molecular Biology, Academia Sinica, for kindly providing guidance and facilities. We also are very thankful to Prof Huai-Jen Tsai, Institute of Fisheries Science, National Taiwan Uni-versity for his helpful guidance. We are grateful to Prof C.W. Hu, Institute of Marine Biotechnology, National Tai-wan Ocean University, for valuable comments and for critically reviewing the manuscript. The Council of Ag-riculture, Republic of China, financially supported this project under grant 92AS-4.2.3-FD-Z3.
REFERENCES
Ali A, Salter-Cid L, Flajnik MF, Heikkila JJ. 1996. Isolation and char-acterization of a cDNA encoding a Xenopus 70kDa heat shock cognate protein, Hsc70. I. Comp Biochem Physiol B Biochem Mol
Biol 113: 681–687.
Ali KS, Dorgai L, Abraham M, Hermesz E. 2003. Tissue-and stress-or-specific differential expression of two hsc70 genes in carp.
Biochem Biophys Res Commun 307: 503–509.
Beckmann RP, Mizzen LA, Welch WJ. 1990. Interaction of Hsp 70 with newly synthesized proteins: implications for protein fold-ing and assembly. Science 248: 850–854.
Benaroudj N, Triniolles F, Ladjimi MM. 1996. Effect of nucleotides, peptides, and unfolded proteins on the self-association of the molecular chaperone HSC70. J Biol Chem 271: 18471–18476. Boorstein WR, Ziegelhoffer T, Craig EA. 1994. Molecular evolution
of the Hsp70 multigene family. J Mol Evol 38: 1–17.
Boutet I, Tanguy A, Rousseau S, Auffret M, Moraga D. 2003. Molec-ular identification and expression of heat shock cognate 70 (hsc70) and heat shock protein 70 (hsp70) genes in the Pacific oyster Crassostrea gigas. Cell Stress Chaperones 8: 76–85. Bukau B, Horwich AL. 1998. The Hsp70 and Hsp60 chaperone
ma-chines. Cell 92: 351–366.
Demand J, Luders J, Hohfeld J. 1998. The carboxy-terminal domain of Hsc70 provides binding sites for a distinct set of chaperone cofactors. Mol Cell Biol 18: 2023–2028.
Flaherty KM, DeLuca-Flaherty C, McKay DB. 1990. Three-dimen-sional structure of the ATPase fragment of a 70 K heat-shock cognate protein. Nature 346: 623–638.
Frydman J, Nimmesgern E, Ohtsuka K, Hartl FU. 1994. Folding of nascent polypeptide chains in a high molecular mass assembly with molecular chaperones. Nature 370: 111–117.
Gething MJ, Sambrook J. 1992. Protein folding in the cell. Nature 355: 33–45.
Gourdon I, Gricourt L, Kellner K, Roch P, Escoubas JM. 2000. Char-acterization of a cDNA encoding a 72 kDa heat shock cognate protein (Hsc72) from the Pacific oyster, Crassostrea gigas. DNA
Seq 11: 265–270.
Gross PS, Barlettt TC, Browdy RW, Chapman RW, Warr GW. 2001. Immune gene discovery by expressed sequence tag analysis of haemocytes and hepatopancreas in the Pacific white shrimp,
Litopenaeus vannamei, and the Atlantic white shrimp, L. setiferus. Dev Comp Immunol 25: 565–577.
Gupta RS, Singh B. 1994. Phylogenetic analysis of 70 kD heat shock
protein sequences suggests a chimeric origin for the eukaryotic cell nucleus. Curr Biol 4: 1104–1114.
Hansen LK, Houchins JP, O’Leary JJ. 1991. Differential regulation of HSC70, HSP70, HSP90 alpha, and HSP90 beta mRNA expres-sion by mitogen activation and heat shock in human lympho-cytes. Exp Cell Res 192: 587–596.
Hartl FU. 1996. Molecular chaperones in cellular protein folding.
Nature 381: 571–579.
Hartl FU, Hayar-Hartl M. 2002. Complex environment of nascent chain to folded protein. Science 295: 1852–1858.
Hasson KW, Hasson J, Aubert H, Redman RM, Lightner DV. 1997. A new RNA-friendly fixative for the preservation of penaeid shrimp samples for virological detection using cDNA genomic probes. J Virol Methods 66: 227–236.
Hu SM, Wang C. 1996. Involvement of the 10-kDa C-terminal frag-ment of hsc70 in complexing with unfolded protein. Arch
Bio-chem Biophys 332: 163–169.
Jacquier-Sarlin MR, Jornot L, Polla BS. 1995. Differential expression and regulation of hsp70 and hsp90 by phorbol esters and heat shock. J Biol Chem 270: 14094–14099.
Kampinga HH, Kanon B, Salomons FA, Kabakov AE, Patterson C. 2003. Overexpression of the cochaperone CHIP enhances Hsp70-dependent folding activity in mammalian cells. Mol Cell
Biol. 23: 4948–4958.
Karlin S, Brocchieri L. 1998. Heat shock protein 70 family: multiple sequence comparisons, function, and evolution. J Mol Evol 47: 565–577.
Kiang JG, Tsokos GC. 1998. Heat shock protein 70 kDa: molecular biology, biochemistry, and physiology. Pharmacol Ther 80: 183– 201.
Knowlton AA, Salfity M. 1996. Nuclear localization and the heat shock proteins. J Biosci 21: 123–132.
Kregel KC. 2002. Heat shock proteins: modifying factors in physio-logical stress responses and acquired thermotolerance. J Appl
Physiol 92: 2177–2186.
Kumar S, Tamura K, Jakobsen IB, Nei M. 2001. MEGA2: molecular evolutionary genetics analysis software. Bioinformatics 17: 1244– 1245.
Laemmli UK. 1970. Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227: 680–685. Lehnert SA, Wilson KJ, Byrne K, Moore SS. 1999. Tissue-specific
expressed sequence tags from the black tiger shrimp Penaeus
monodon. Mar Biotechnol (NY) 1: 465–476.
Leng CH, Brodsky JL, Wang C. 1998. Isolation and characterization of a DnaJ-like protein in rats: the C-terminal 10-kDa domain of hsc70 is not essential for stimulating the ATP-hydrolytic activity of hsc70 by a DnaJ-like protein. Protein Sci 7: 1186–1194. Lindquist S, Craig EA. 1988. The heat-shock proteins. Annu Rev
Gen-et 22: 631–677.
Mallouk Y, Vayssier-Taussat M, Bonventre JV, Polla BS. 1999. Heat shock protein 70 and ATP as partners in cell homeostasis (Re-view). Int J Mol Med 4: 463–474.
Nei M, Kumar S. 2000. Molecular Evolution and Phylogenetics. Oxford University Press, New York.
O’Malley K, Mauron A, Barchas JD, Kedes L. 1985. Constitutively expressed rat mRNA encoding a 70-kilodalton heat shock-like protein. Mol Cell Biol 5: 3476–3483.
Palleros DR, Welch WJ, Fink AL. 1991. Interaction of hsp70 with unfolded proteins: effects of temperature and nucleotides on the kinetics of binding. Proc Natl Acad Sci U S A 88: 5719–5723. Rensing SA, Maier UG. 1994. Phylogenetic analysis of the stress-70
protein family. J Mol Evol 39: 80–86.
Ritossa E. 1962. A new puffing pattern induced by heat shock and DNP in Drosophila. Experientia 18: 571–573.
Sadis S, Hightower LE. 1992. Unfolded proteins stimulate molecular chaperone Hsc70 ATPase by accelerating ADP/ATP exchange.
Biochemistry 31: 9406–9412.
Santacruz H, Vriz S, Angelier N. 1997. Molecular characterization of a heat shock cognate cDNA of zebrafish, hsc70, and devel-opmental expression of the corresponding transcripts. Dev
Gen-et 21: 223–233.
Takeda S, McKay DB. 1996. Kinetics of peptide binding to the bovine 70 kDa heat shock cognate protein, a molecular chaperone.
Bio-chemistry 35: 4636–4644.
Taviara M, Gabriele T, Kola I, Anderson RL. 1996. A hitchhiker’s guide to the human Hsp70 family. Cell Stress Chaperones 1: 23– 28.
Tsai MY, Wang C. 1994. Uncoupling of peptide-stimulated ATPase and clathrin-uncoating activity in deletion mutant of hsc70. J
Biol Chem 269: 5958–5962.
Vayssier M, Le Guerhier F, Fabien JF, et al. 1999. Cloning and anal-ysis of a Trichinella britovi gene encoding a cytoplasmic heat shock protein of 72 kDa. Parasitology 119: 81–93.
Wang C, Lee MR. 1993. High-level expression of soluble rat hsc70 in Escherichia coli: purification and characterization of the cloned enzyme. Biochem J 294: 69–77.
Xia X. 2000. Data Analysis in Molecular Biology and Evolution. Kluwer Academic Publishers, Boston, MA.
Xia X, Xie Z. 2001. DAMBE: data analysis in molecular biology and evolution. J Hered 92: 371–373.