Concurrent infections by two dengue virus serotypes among dengue patients in Taiwan
7
0
0
全文
(2) Dengue virus serotypes in Taiwan. different serotypes of dengue virus are expected to occur during epidemics involving multiple dengue virus serotypes [3,18]. The first case of dual infection with 2 dengue viral serotypes (DEN-1 and DEN-4) was reported in Puerto Rico in 1982 [13]. There were 6 cases of dual infections (DEN-1 and DEN-3) in New Caledonia in 1989 [14]. Since then, more cases of concurrent infections by multiple dengue virus serotypes have been reported in different countries [1518]. Although outbreaks involving more than 1 dengue virus serotype have been noted in Taiwan since 1987, cases concurrently infected by 2 or more serotypes of dengue virus have not been documented [9-12]. In this study, we used a modified multiplex reverse transcription-polymerase chain reaction (RT-PCR) assay [19,20] to examine acute plasma samples from suspected dengue patients during an outbreak in southern Taiwan in 2000. Of the 21 confirmed dengue patients, most were infected by either DEN-2 or DEN3 virus, whereas 2 were concurrent infections of DEN2 and DEN-3 viruses. We report here that concurrent infections do occur in Taiwan.. Materials and Methods Study participants The diagnoses of DF and DHF followed the WHO clinical definition [4]. Detection of dengue genomic sequences in plasma by a multiplex RT-PCR assay, which was modified from the previously described RTPCR assays, was the laboratory criteria of confirmation for all cases [4,19,20]. Between June 1 and August 31, 2000, during an outbreak in southern Taiwan, 21 DF patients from 5 hospitals (Kuo General Hospital, SinLau Christian Hospital, Yuan General Hospital, Pingtung Christian Hospital, and Tian-Sheng Memorial Hospital) were confirmed at our laboratory. The day of onset of fever is defined as day 1 of illness. One acute blood sample was collected for each patient in an ethylenediaminetetraacetic acid (EDTA)-containing tube within 8 days of illness. The plasma was prepared within 6 hours of collection and stored at –80oC until use [21]. Isolation of viral RNA Dengue viral RNA was isolated from the plasma or stock virus using the QIAamp viral RNA mini kit (Qiagen, Germany) as described previously [21]. Plasma samples obtained from 2 hepatitis C virus (HCV) carriers and 2 healthy donors were also subjected to RNA isolation. Stock viruses used in this study included the 4 serotypes of dengue virus, Hawaii (DEN90. 1), New Guinea (DEN-2), H-87 (DEN-3), and H-241 (DEN-4) strains, with titers of 2.1 x 106 plaque forming units per millimeter (pfu/mL), 1.3 x 106 pfu/mL, 2 x 106 pfu/mL and 1 x 106 pfu/mL, respectively. They were obtained from culture supernatants of infection of mosquito C6/36 cells and then titrated on BHK cells by standard plaque assay. Three Japanese encephalitis virus (JEV) strains (Nakayama vaccine strain, Beijing vaccine strain, and CH1949 Taiwan local strain) with titers ranging from 105 to 106 pfu/mL were also included in the analysis [21].. Multiplex RT-PCR The multiplex RT-PCR assay employed in this study was modified from the previously described RT-PCR assays, which can distinguish the 4 dengue serotypes by the size of the products [19,20]. It includes a step of RT-PCR using a highly conserved primer pair, D1 and D2, and a step of second-round PCR using the primer D1 and 4 serotype-specific primers, TS1, TS2, TS3, and TS4 (Fig. 1A). The sequences of the primers are as follows: D1, 5'-TCAATATGCTGAAACGCGCGAGAAACCG-3' (genome positions 134 to 161 of the DEN2 Jamaica strain [22]); D2, 5'-TTGCACCAACAGTCAATGTCTTCAGGTTC-3' (genome positions 644 to 611 of the DEN-2 Jamaica strain); TS1, 5'CGTCTCAGTGATCGGGGG-3' (genome positions 586 to 568 of the DEN-1 Hawaii strain [23]); TS2, 5'CGCCACAAGGGCCATGAAC AG-3' (genome positions 252 to 232 of the DEN-2 Jamaica strain); TS3, 5'-TAACATCATCATGAGACAGAGC-3' (genome positions 421 to 400 of the DEN-3 H-87 strain [24]); and TS4, 5'-CTCTGTTGTCTTAAACAAGAGA-3' (genome positions 527 to 506 of the DEN-4 H-241 strain [25]). RT-PCR and PCR were performed in a separate room from that used for RNA isolation, and precautions for PCR were followed to avoid contamination [26]. An aliquot (2 µL) of RNA eluate was subjected to RTPCR using the Superscript one-step RT-PCR kit (Gibco/ BRL, Life Technologies, Rockville, MD, US) under the conditions of 52oC for 40 min, and 95oC for 5 min, followed by 30 cycles of 94oC for 1 min, 60oC for 1 min, and 72oC for 1 min, and a final step of 72oC for 7 min. The expected size of the RT-PCR products is 511bp (D1 and D2). An aliquot (2 µL) of the diluted products (1:100) was subjected to the second round PCR using the Taq DNA polymerase (HT Biotechnology, Cambridge, England) under the conditions of 94oC for 5 min, and 35 cycles of 94oC for 30 sec, 60oC for 30 sec, and 72oC for 45 sec, and a final step of 72oC for 10 min. The expected sizes of the amplified products are.
(3) Wang et al. A. B. gel of ABI automated sequencers (Applied Biosystems ABI-377) as described previously [21].. Sequence analysis The nucleotide sequences of a fragment of the 119-bp products and those of the 290-bp products were aligned with sequences of the corresponding regions of the 4 dengue virus serotypes, including the prototype DEN1 (45AZ5), DEN-2 (New Guinea), DEN-3 (H-87), and DEN-4 (H-241) viruses, as well as of the JEV SA14wt strain, using the program Dnaman Version 4.15 (Lynnon Biosoft, Canada). The nucleotide sequence similarity of each comparison was calculated. A phylogenetic tree was generated by the program MEGA version 2.1 (Molecular Evolutionary Genetics Analysis, Pennsylvania State University, PA, US) using the neighbor-joining method.. Results. Fig. 1. (A) Schematic diagram of the dengue virus genome and the primers used in the modified multiplex RT-PCR assay. The predicted sizes of the products are shown. (B) The ethidium bromide stained gel of the products of the first round RT-PCR and second round PCR, using RNA derived from the 4 dengue virus serotypes, DEN-1 (Hawaii), DEN-2 (New Guinea), DEN-3 (H-87), and DEN-4 (H-241). Mock = no RNA template added; m = molecular size marker.. 482-bp (D1 and TS1 for DEN-1 virus), 119-bp (D1 and TS2 for DEN-2 virus), 290-bp (D1 and TS3 for DEN-3 virus), and 392-bp (D1 and TS4 for DEN-4 virus). The products were electrophoresed through 2% agarose gel, stained with ethidium bromide, and examined under ultraviolet light using a digital gel documentation system as described previously [21].. Sequencing of the amplification products The products of predicted size from the second round were cut from the gel and purified with the GFXTM PCR DNA and gel band purification kit (Amersham Pharmacia, Piscataway, NJ, US). The purified products were sequenced using the BigDye terminator cycle sequencing kit under conditions recommended by the manufacturer (PE Applied Biosystems, Foster City, CA, US). Samples were loaded onto 4.75% polyacrylamide. Typing of dengue viruses by a modified multiplex RT-PCR assay To determine the dengue virus serotypes of plasma samples, a multiplex RT-PCR assay was modified from the previously described RT-PCR assays [19,20]. This includes a step of RT-PCR using a highly conserved primer pair, D1 and D2, and a step of second-round PCR using the primer D1 and 4 serotype-specific primers, TS1, TS2, TS3, and TS4 (Fig. 1A). An aliquot of RNA eluates derived from stock viruses of the 4 dengue serotypes, Hawaii (DEN-1), New Guinea (DEN2), H-87 (DEN-3), and H-241 (DEN-4) strains, was subjected to RT-PCR first. As shown in Fig. 1B left, RT-PCR products of the predicted size of 511-bp were seen in reactions containing the RNA templates derived from the 4 dengue viruses but not in the reaction containing no template. An aliquot of the diluted products was then subjected to the second-round PCR. Amplified products of the expected sizes of 482-bp, 119-bp, 290-bp, and 392-bp were seen in the reactions of DEN-1, DEN-2, DEN-3, and DEN-4 viruses, respectively (Fig. 1B right). RNA templates derived from other flaviviruses prevalent in Taiwan, including 3 JEV and 2 HCV, as well as from the plasma of 2 healthy donors were also subjected to the multiplex RTPCR assay. None of these resulted in amplified products of expected size in the first-round RT-PCR or in the second-round PCR (data not shown). Determination of dengue viral serotypes of confirmed dengue patients Between June 1 and August 31, 2000, 49 acute blood 91.
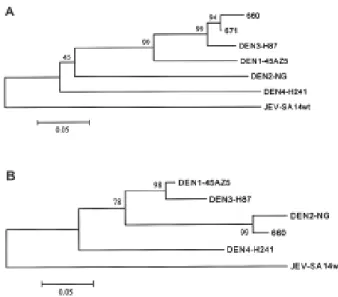
(4) Dengue virus serotypes in Taiwan. samples of 49 suspected dengue cases from 5 hospitals in southern Taiwan were sent to the laboratory. RNA isolated from plasma was subjected to the modified multiplex RT-PCR assay to detect and type dengue virus serotypes. Among the 49 samples, 21 were confirmed dengue cases based on the presence of amplified products of predicted sizes. The results of some samples are shown in Fig. 2. While most cases were found to have products of 119-bp, the predicted size of DEN-2 virus, some had products of 290-bp, the predicted size of DEN-3 virus, or 392-bp, that of DEN-4 virus. Interestingly, 2 different size RT-PCR products, 119bp and 290-bp, were seen in 2 cases (patients 660 and 671), suggesting that they were concurrently infected by 2 dengue viruses, DEN-2 and DEN-3 (Fig. 2). The serotypes of the 21 confirmed dengue patients were summarized in Table 1. There were 15 DEN-2 cases, 3 DEN-3 cases, 1 DEN-4 case, and 2 dual infections by DEN-2 and DEN-3 viruses.. Sequence analysis To confirm the identities of the amplified products of the 2 dual infected patients, the 119-bp bands and 290bp bands derived from both cases were purified from the gel and subjected to direct DNA sequencing. The sequences of a 124-nucleotide region of the 290-bp products from patients 660 and 671 were aligned with sequences of the corresponding region of the 4 prototype dengue viruses, DEN-1 (45AZ5), DEN-2 (New Guinea), DEN-3 (H-87), and DEN-4 (H-241), and the similarity of the nucleotide sequences was analyzed. As shown in Table 2, the nucleotide sequences of the 290-bp products from patients 660 and 671 had higher degrees of similarity (93.5% and 96.0%, respectively) to that of DEN-3 virus than to those of other serotypes,. Fig. 2. Determination of dengue virus serotypes of plasma samples by the modified multiplex RT-PCR assay. The patients’ ID is shown at the top. The predicted sizes of the amplified products of the four dengue viruses, DEN-1 (Hawaii), DEN-2 (New Guinea), DEN-3 (H-87), and DEN-4 (H-241), are shown at the right. m = molecular size marker.. 92. Table 1. Serotypes of dengue patients determined by the modified multiplex RT-PCR assay Serotype. No. of cases. DEN-2 DEN-3 DEN-4 DEN-2 and DEN-3. 15 3 1 2. Total. 21. suggesting that these were DEN-3 sequences. Phylogenetic analysis was also performed and revealed a clustering of the sequences from both patients with that of DEN-3 virus, indicating that these were indeed DEN-3 sequences (Fig. 3A). The sequence of a 47-nucleotide region of the 119bp product from patient 660 was also aligned with the corresponding sequences of the 4 dengue viruses and subjected to similar analysis. The 119-bp product from patient 671 was not included in this analysis, as the yield was too low for direct DNA sequencing. The nucleotide sequence of the 119-bp product from patient 660 was closer to that of DEN-2 virus (similarity, 93.6%) than to those of other serotypes, suggesting that this was DEN-2 sequence (Table 2). This was further supported by the clustering of this sequence with that of DEN-2 virus in the phylogenetic analysis (Fig. 3B). Taken together, the sequence analysis of the amplified. FIG. 3. Phylogenetic analysis of the sequences of the amplified products of 290-bp (A) and 119-bp (B) from two dual infected cases, patients 660 and 671, and the sequences of the four prototype dengue viruses, DEN-1 (45AZ5), DEN-2 (New Guinea), DEN-3 (H-87), and DEN-4 (H-241), using the program MEGA. The sequence of JEV strain Sa14wt was also included in the analysis. Numbers at nodes represent bootstrap values..
(5) Wang et al. Table 2. Nucleotide sequence (%) similarity between the amplified products from dual infected patients and the 4 dengue virus serotypes The 290-bp product a DEN-1c DEN-2c DEN-3c DEN-4c. The 119-bp productb. Patient 660. Patient 671. Patient 660. 83.1 67.7 93.5 63.7. 85.5 69.4 96.0 62.9. 80.9 93.6 74.5 63.8. a. A 124-nucleotide region of the PCR product was sequenced and analyzed. A 47-nucleotide region of the PCR product was sequenced and analyzed. c The 4 dengue viruses used in the analysis are 45AZ5 (DEN-1), New Guinea (DEN-2), H-87 (DEN-3), and H241 (DEN-4). b. products from these 2 patients confirmed that they were concurrently infected by 2 dengue viruses, DEN-2 and DEN-3.. Discussion It is generally believed that concurrent infections by different serotypes of dengue virus will occur during epidemics in which multiple dengue virus serotypes are being transmitted. Although co-circulation of multiple serotypes of dengue virus in the same region has been recognized in several countries for decades [3], it was not until 1982 that the first case of concurrent infections with 2 dengue virus serotypes was reported [13]. The detection of concurrent infections could be attributed to the common use of serotype-specific monoclonal antibodies in the immunofluorescence assay to detect different dengue virus serotypes during routine virus isolation [13,27-29]. In addition, incorporation of serotype-specific primers in the RT-PCR assays further facilitates the detection of concurrent infections [1418]. In this study, we developed a modified multiplex RT-PCR assay to examine acute plasma samples from dengue patients during an outbreak in southern Taiwan in 2000. To our knowledge, this study reports for the first time that concurrent infections by 2 serotypes of dengue virus occur in Taiwan. Compared with the RT-PCR assay described previously by Lanciotti et al [19], which required 4 separate reactions to type dengue viruses, our modified multiplex RT-PCR assay was simpler and more convenient. The multiplex RT-PCR assay reported by Harris et al combined the steps of RT and PCR in 1 single tube, and therefore had only 1 round of PCR amplification [20]. Our multiplex RT-PCR assay employs 2 rounds of amplification (the first round RTPCR step and the second round PCR step) as the original RT-PCR assay by Lanciotti et al, of which the sensitivity was reported to be similar to that of the virus isolation or immunofluorescence assay system [18,19,29]. With regard to the specificity, our multiplex RT-PCR assay. uses the same primers as those of the RT-PCR assay by Lanciotti et al, which has been shown to be specific for dengue virus [20]. Consistent with this, the present assay can detect the 4 dengue virus serotypes, but not other flaviviruses prevalent in Taiwan, including JEV and HCV (data not shown). In this study, we identified 2 cases of concurrent infection by the RT-PCR assay, which is one of the laboratory criteria for confirmation of dengue cases according to the WHO clinical definition [4]. Since only acute plasma samples were available, serological tests could not be used for confirmation. Sequence analysis was therefore conducted to confirm the identities of the amplified products of the 2 dually infected patients. Due to technical limitation in sequencing DNA directly from PCR products, only a 47-nucleotide fragment of the 119bp band was sequenced. Comparison of the nucleotide sequence of this region of patient 660 with those of the 4 dengue serotypes revealed that it had a similarity of 93.6% to DEN-2 virus, which is within the range of nucleotide similarity between dengue viruses of the same serotype in this region (Table 2 and data not shown). The nucleotide similarities of this region to those of DEN-1, DEN-3, and DEN-4 viruses were from 63.8 % to 80.9%, which were in the range of the nucleotide similarity between dengue viruses of different serotypes (Table 2 and data not shown) [30]. These results indicate that this was indeed the DEN-2 sequence, which was also confirmed by phylogenetic analysis (Fig. 3B). Similarly, comparison of the nucleotide sequences of a 124-nucleotide region of the 290-bp products from patients 660 and 671 with those of the 4 dengue virus serotypes, together with the phylogenetic analysis, demonstrated that the 290-bp products from both patients were DEN-3 sequences (Table 2 and Fig. 3A). It should be noted that the possibility of cross-contamination between samples was remote, since the nucleotide distance of the 124nucleotide capsid region between patients 660, 671, and the DEN-3 virus (H87 strain) used in this study was 93.
(6) Dengue virus serotypes in Taiwan. from 2.4% to 6.5%, a range higher than that of sequence diversity of the capsid gene of DEN-3 virus within the same patient (mean pairwise p-distance 0.12% to 1.02%) [31]. With the dramatic changes in the global epidemiology of dengue virus infection during the past 50 years, there are not only increases in the number of countries reported to have dengue cases but also in the number of severe and fatal cases [1,3,5]. The change in the epidemiological patterns from hypoendemicity (only 1 serotype) to hyperendemicity (multiple serotypes) has been seen in many countries [1,3]. Using a modified multiplex RT-PCR assay, this study demonstrated that concurrent infections also occur in Taiwan. The frequency of concurrent infection in this study, 9.5% (2/21) was close to the frequencies reported by LoronoPino et al (11% in Indonesia and 5.5% in Mexico, Puerto Rico, and Indonesia together) [18]. Common occurrence of concurrent infections by multiple dengue virus serotypes has been recognized recently and is expected to be seen in the future [18]. Whether concurrent infections are associated with more severe diseases remain to be investigated. Although the epidemiological pattern of dengue in Taiwan, like those in other parts of the world, has changed in the past 20 years, active surveillance, epidemiological investigation, effective mosquito control, and public health infrastructure all together are believed to contribute to the reduction in the number of cases during each outbreak [12]. The epidemiology of dengue in Taiwan is very different from that in other southeast Asian countries in that it contains imported cases every year and has not been considered yet as a hyperendemic region [12]. The present findings, however, indicate that one of the features commonly seen in hyperendemic areas is also found in southern Taiwan. Obviously, the future virologic surveillance of dengue outbreaks in Taiwan should be conducted from a new perspective.. Acknowledgments We thank Shu-Mei Chang at the Yuan’s General Hospital, Hsiu-O Kuo at the Pingtung Christian Hospital, Chiang-Chyuan Su at the Tian-Sheng Memorial Hospital, Shih-Ting Ho at the Sin Lau Christian Hospital, and Shih-Chung Lin at the Kuo General Hospital for kindly providing clinical samples, and Tzu-Ling Sung, Yu-Chen Tsai, and Tsai-Yu Lin for technical assistance. We thank Dr. DJ Gubler for the DEN-2 New Guinea strain, and the DEN-3 H87 strain. We also thank Dr. Chuan-Liang Kao for providing the four stock viruses. This work was supported by the National Health Research Institute (NHRI-CN94. CL8903P), and in part by the National Science Council (NSC90-2320-B-002-150), Taiwan, Republic of China. References 1. Monath TP. Dengue: the risk to developed and developing countries. Proc Natl Acad Sci USA 1994;91:2395-400. 2. Innis BL. Dengue and dengue hemorrhagic fever. In: Porterfield JS, ed. Exotic Viral Infections. London: Chapman & Hall; 1995: 103-46. 3. Gubler DJ. Dengue and dengue hemorrhagic fever. Clin Microbiol Rev 1998;11:480-96. 4. World Health Organization. Dengue hemorrhagic fever: diagnosis, treatment and control. 2nd ed. Geneva: World Health Organization, 1997. 5. World Health Organization. Fact sheet No 117, April 2002. Geneva: World Health Organization, 2002. 6. Hsieh WC, Chen MF, Lin KT, Hsu ST, Ma CI, Wu SS. Study of outbreak of dengue fever in 1981 in Liouchyou Shiang, Pingtung County. J Fornosan Med Assoc 1982;81:1388-95. 7. Wu YC. Epidemic dengue 2 on Liouchyou Shaing, Pingtung County in 1981. Chinese J Microbiol Immunol 1986;19:20311. 8. Bureau of Communicable disease Control. Preliminary investigation report of an outbreak of dengue fever in Kaohsiung, southern Taiwan. Epidemiol Bull 1987;December: 93-5. 9. Ko YC. Epidemiology of dengue fever in Taiwan. Kaohsiung J Med Sci 1989;5:1-11. 10. Wu YC, Lien JC, Chen HY. Recent outbreak of dengue in Taiwan. Trop Med 1993;35:201-7. 11. Huang KP. Dengue fever and dengue hemorrhagic fever. Formosan J Med 1997;1:50-6. 12. King CC, Wu YC, Chao DY, Lin TH, Chow L, Wang HT, Ku CC, Kao CL, Hwang KP, Lam SK, Gubler DJ. Major epidemics of dengue in Taiwan in 1991-2000: related to intensive virus activities in Asia. Dengue Bull. Geneva: World Health Organization 2000;24:1-10,. 13. Gubler DJ, Kuno G, Sather GE, Waterman SH. A case of natural concurrent human infection with two dengue viruses. Am J Trop Med Hyg 1985;34:170-3. 14. Laille M, Deubel V, Sainte-Marie FF. Demonstration of concurrent dengue 1 and dengue 3 infection in six patients by the polymerase chain reaction. J Med Virol 1991;34:51-4. 15. Maneekarn N, Morita K, Tanaka M, Igarashi A, Usawattanakul W, Sirisanthana V, Innis BL, Sittisombut N, Nisalak A, Nimmanitya S. Applications of polymerase chain reaction for identification of dengue viruses isolated from patient sera. Microbiol Immunol 1993;37:41-7. 16. Fang M, Chen H, Chen C, Tian X, Jiang L, Peng Y, Chen W, Guo H. Detection of flaviviruses by reverse transcriptionpolymerase chain reaction with the universal primer set. Microbiol Immunol 1997;41:209-13. 17. Kanesa-Thasan N, Chang GJ, Smoak BL, Magill A, Burrous MJ, Hoke Jr CH. Molecular and epidemiologic analysis of dengue virus isolates from Somalia. Emerg Infect Dis 1998;4: 299-303. 18. Lorono-Pino MA, Cropp CB, Farfan JA, Vorndam AV, Rodriguez-Angulo EM, Rosado-Paredes EP, Flores-Flores LF, Beaty BJ, Gubler DJ. Common occurrence of concurrent infections by multiple dengue virus serotypes. Am J Trop Med Hyg 1999;61:725-30..
(7) Wang et al. 19. Lanciotti RS, Calisher CH, Gubler DJ, Chang GJ, Vorndam AV. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J Clin Microbiol 1992;30:545-51. 20. Harris E, Roberts G, Smith L, Selle J, Kramer LD, Valle S, Sandoval E, Balmaseda A. Typing of dengue viruses in clinical specimens and mosquitoes by singe-tube multiplex reverse transcriptase PCR. J Clin Microbiol 1998;36:2634-9. 21. Wang WK, Lee CN, Kao CL, Lin YL, King CC. Quantitative competitive reverse transcription-PCR for quantification of dengue virus RNA. J Clin Microbiol 2000;38:3306-10. 22. Deubel V, Kinney RM, Trent DW. Nucleotide sequence and deduced amino acid sequence of the structural proteins of dengue type 2 virus, Jamaica genotype. Virology 1986;155: 365-77. 23. Mason PW, McAda PC, Mason TL, Fournier MJ. Sequence of the dengue-1 virus genome in the region encoding the three structural proteins and the major nonstructural protein NS1. Virology 1987;161:262-7. 24. Osatomi K, Sumiyoshi H. Complete nucleotide sequence of dengue type 3 virus genome RNA. Virology 1990;176:643-7. 25. Zhao B, Mackow E, Buckler-White A, Markoff L, Chanock. RM, Lai CJ, Makino Y. Cloning full-length dengue type 4 viral DNA sequences: analysis of genes coding for structural proteins. Virology 1986;155:77-88. 26. Kwok S, Higuchi R. Avoiding false positive with PCR. Nature 1989;339:237-8. 27. Henchal EA, McCown JM, Sequin MC, Gentry MK, Brandt WE. Rapid identification of dengue virus isolates by using monoclonal antibodies in an indirect immunofluorescence assay. Am J Trop Med Hyg 1983;32:164-9. 28. Gubler DJ, Kuno G, Sather GE, Velez M, Oliver A. Mosquito cell cultures and specific monoclonal antibodies in surveillance for dengue viruses. Am J Trop Med Hyg 1984;33:158-65. 29. Vorndam V, Kuno G. Laboratory diagnosis of dengue virus infections. In: Gubler DJ, Kuno G, eds. Dengue and Dengue Hemorrhagic Fever. London: CAB International, 1997:313-34. 30. Mandl CW, Heinz FX, Kunz C. Sequence of the structural proteins of tick-borne encephalitis virus (western subtype) and comparative analysis with other flaviviruses. Virology 1988; 166:197-205. 31. Wang WK, Sung TL, Lee CN, Lin TY, King CC. Sequence diversity of the capsid gene and the nonstructural gene NS2B of dengue-3 virus in vivo. Virology 2002;303:181-91.. 95.
(8)
數據


相關文件
You are given the wavelength and total energy of a light pulse and asked to find the number of photons it
好了既然 Z[x] 中的 ideal 不一定是 principle ideal 那麼我們就不能學 Proposition 7.2.11 的方法得到 Z[x] 中的 irreducible element 就是 prime element 了..
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
volume suppressed mass: (TeV) 2 /M P ∼ 10 −4 eV → mm range can be experimentally tested for any number of extra dimensions - Light U(1) gauge bosons: no derivative couplings. =>
We explicitly saw the dimensional reason for the occurrence of the magnetic catalysis on the basis of the scaling argument. However, the precise form of gap depends
For pedagogical purposes, let us start consideration from a simple one-dimensional (1D) system, where electrons are confined to a chain parallel to the x axis. As it is well known
The observed small neutrino masses strongly suggest the presence of super heavy Majorana neutrinos N. Out-of-thermal equilibrium processes may be easily realized around the
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
