國立高雄大學生命科學系
碩士論文
台灣養殖南美白對蝦感染
Enterocytozoon hepatopenaei
(EHP)檢驗方法之建立
Establishment of diagnostic methods to detect
Enterocytozoon hepatopenaei (EHP) infection in culture
Litopenaeus vannamei in Taiwan
研究生:劉于瑄 撰
指導教授:王俊順 博士
謝誌
首先感謝恩師 王俊順教授在就讀期間研究所三年來的用心與耐心 的指導與鼓勵,研究所三年來學生受益良多,並在待人處事、時間安排 及研究學習上給予許多的建議。感謝口試委員:溫秋明教授及屏東科技 大學水產養殖系 鄭文騰教授在口試及論文修改上提供寶貴的意見與鼓 勵,使得論文更加完整。 感謝黃永森、高佑靈、楊文仁、葛孟杰、黃重期、施能朗、溫秋明 等老師在課業上的指導;感謝系辦的明坤、美純給予學校課業及生活中 各方面的協助;感謝張瑞昕博士協助基因序列的比對及親緣關係的分 析,感謝鄭翔文學長分享及交流許多實驗上的經驗與協助;感謝蕭裕承 同學在實驗上的協助與平日的關心及鼓勵,讓我三年的研究所生活更加 充實、順利。 最後感謝我的家人,一路上在背後支持著我、鼓勵著我,讓我毫無 顧忌的往前邁進。 劉于瑄 僅誌 中華民國108 年 7 月I
目錄
目錄 ... I 圖目錄 ... III 表目錄 ... V 摘要: ... VI 第一章 前言 ... 1 1.1 南美白對蝦的習性介紹 ... 1 1.2 南美白對蝦之常見的疾病 ... 2 1.3 微孢子蟲的特性 ... 41.4 Enterocytozoon hepatopenaei (EHP)的特性與歸類 ... 5
1.5 Enterocytozoon hepatopenaei ( EHP )之地理分布及症狀... 6
1.6 Enterocytozoon hepatopenaei ( EHP )傳播方式及感染途徑 ... 7
1.7 Enterocytozoon hepatopenaei (EHP)的檢測方法 ... 8
1.7.1 外表病徵觀察 ... 8
1.7.2 電子顯微鏡技術 ... 9
1.7.3 組織病理切片 ... 9
1.7.4 DNA 探針 ... 10
1.7.5 聚合鏈連鎖反應(Polymerase chain reaction,PCR) ... 10
1.7.6 環介導的等溫擴增(loop-mediated isothermal amplification,LAMP ) . 10 1.8 實驗目的... 11 第二章 材料與方法 ... 12 2.1 南美白對蝦微孢子 EHP 病學之研究 ... 12 2.1.1 採樣 ... 12 2.1.2 病理組織切片觀察 ... 12 2.2 EHP 之部分基因片段序列選殖 ... 13 2.2.1 去氧核醣核酸的(Total DNA)萃取 ... 13 2.2.2 聚合酶連鎖反應(PCR)... 14 2.2.3 膠體純析(Gel elution) ... 15
2.2.4 DNA 接合(DNA Ligation) ... 15
2.2.5 質體轉型(Transformation)... 15
2.2.6 質體抽取(Plasmid extraction) ... 16
2.2.7 限制酶素切割(Restriction enzyme digestion) ... 16
2.2.8 核酸序列之分析與比對 ... 16
2.3 EHP 微孢子蟲之檢測方式與分析 ... 17
II
2.3.1.1 探針之製備與純化 ... 17
2.3.1.2 點墨雜交反應(Dot blot Hybridization) ... 18
2.3.2 原位雜交反應(In situ hybridization) ... 18
第三章 結果 ... 20
3.1 南美白對蝦 EHP 微孢子蟲流行病學之研究 ... 20
3.2 組織病理觀察 ... 21
3.3 EHP 微孢子蟲片段基因的選殖 ... 21
3.3.1 SSU rDNA 之片段基因 PCR 轉質和序列分析 ... 21
3.3.2 EHP 微孢子蟲 SSU RNA 片段基因序列比對 ... 22
3.4 EHP 微孢子蟲之檢測方式之建立與分析 ... 23
3.4.1 點墨雜交試驗(Dot blot hybridization) ... 23
3.4.2 原位雜交試驗(In situ hybridization) ... 23
第四章 討論 ... 40
第五章 結論 ... 45
第六章 參考文獻... 46
III
圖目錄
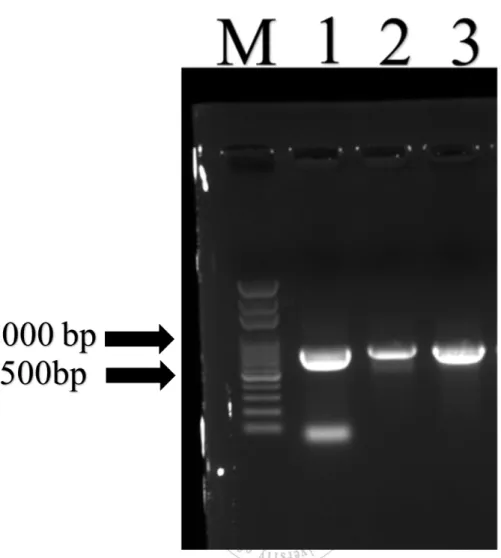
圖 1 利用 0.8%洋菜膠電泳圖,使用 EHP 微孢子蟲專一性引子 ENR779/ENF779 偵 測,檢測到白蝦感染 EHP 微孢子蟲。M 為 100bp ladder DNA。 ... 26 圖 2 0.8%洋菜膠電泳圖,使用 Nested PCR 偵測,更加確定白蝦感染... 27 圖 3 EHP 微孢子蟲感染白蝦肝胰臟病理切片圖 A 跟 C 圖為感染 EHP 微孢子蟲之
白蝦肝胰臟,肝胰臟組織中分布EHP 微孢子蟲(框起來的位子)。B 圖是 A 圖
肝胰臟感染範圍放大中的EHP 微孢子蟲的分布情形。D 圖為 C 圖肝胰臟感染
範圍放大中的EHP 微孢子蟲分布情形。 ... 28 圖 4 pGEM-T Easy 載體回收純化 DNA 片段,以 TA cloning 的方式,植入到
pGEM-T Easy 載體中,經轉型作用(Transformation)到大腸桿菌 JM109 中,白
色是成功轉型到大腸桿菌 JM109 的菌株,藍色則是未成功轉型的菌株。 ... 29
圖 5 0.8%洋菜膠電泳分析圖,Lane 1-3 均為轉型後的重組基因,經限制酶切割 後,顯示重組基因含EHP 微孢子蟲片段基因(約 779 bp)。M 為 100bp ladder DNA。 ... 30 圖 6 EHP 微孢子蟲核酸定序與其他國家 EHP 微孢子蟲之比對圖:將 EHP 微孢子
蟲SSU rDNA 基因序列與泰國、印度、越南、中國、美國、拉丁美洲等國家
的EHP 微孢子蟲基因序列進行比對,分別得到 99%、99.87%、99.87%、
99.77%、99.22%的序列相似度 ... 34 圖 7 基因親緣關係比較圖 EHP 微孢子蟲 SSU rDNA 片段基因親緣關係之比較,台
灣 EHP 微孢子蟲與泰國、印度、美國、中國的親緣關係比較近,與西班牙 EHP 微孢子蟲的親緣關係比較遠。 ... 35 圖 8 0.8 %洋菜膠電泳分析圖,利用 PCR 的方法將探針進行 DIG 標定。Land 1-2 均
為經DIG 標識之 ATPase 之部分基因片段序列探針。M 為 100bp ladder DNA。 36
圖 9 點墨雜交法(Dot blot hybridization)之靈敏度試驗圖,將 DIG 標識的 EHP 微孢 子蟲探針,利用已建構完成之南美白對蝦兩組重組質體進行之靈敏度測試,從
圖片中可以看見含量高於13pg/μL 的 EHP 微孢子蟲核酸重組質體均可被測
得。 37
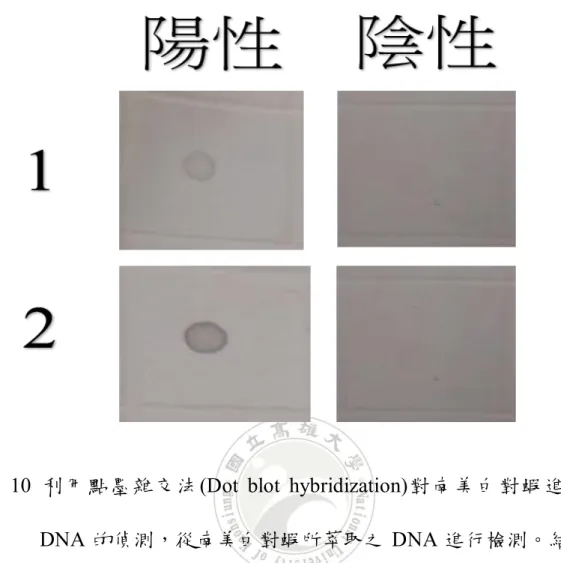
圖 10 點墨雜交法(Dot blot hybridization ),利用點墨雜交法(Dot blot hybridization )
對南美白對蝦進行組織DNA 的偵測,從南美白對蝦所萃取之 DNA 進行檢
測。結果可檢測到南美白對蝦感染EHP 微孢子蟲的病原 DNA。 ... 38 圖 11 原位雜交法(In situ hybridization),利用原位雜交法(In situ hybridization),以
DIG 標識的 EHP 微孢子蟲探針,偵測 EHP 微孢子蟲在南美白對蝦中分布的情 形,藍色沉澱表示陽性反應,A 圖病蝦的肝胰臟組織被 EHP 微孢子蟲感染, 呈現陽性反應,B 圖為 A 圖的放大圖,亦可發現組織中微量感染可被偵測出來
IV
V
表目錄
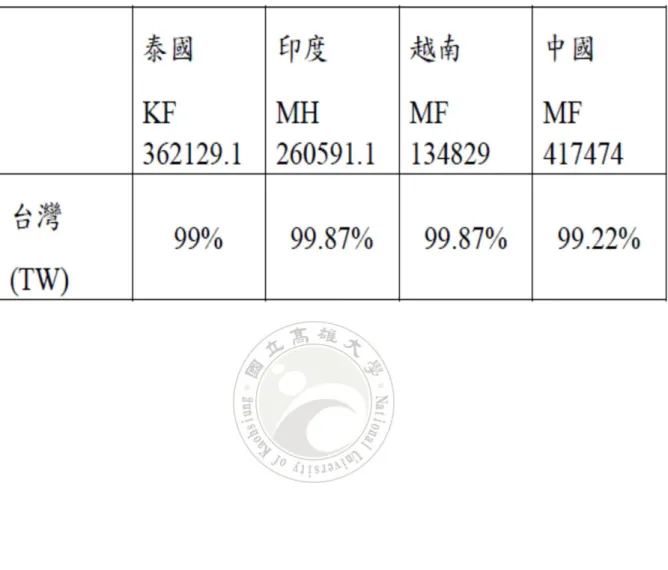
表 1 台灣南部地區養殖南美白對蝦採樣紀錄表 ... 24 表 2 台灣 EHP 與世界各國 EHP 的相似性 ... 25
VI
台灣養殖南美白對蝦感染
Enterocytozoon hepatopenaei
(EHP)檢驗方法之建立
指導教授:王俊順 博士 國立高雄大學生命科學系 學生:劉于瑄 國立高雄大學生命科學系 摘要:
本 研 究 針 對 台 灣 養 殖 南 美 白 對 蝦 (Litopenaeus vannamei) 感 染 Enterocytozoon
hepatopenaei (EHP)微孢子蟲之疫情進行調查,並利用分子生物學的技術建立微孢子
蟲的偵測方法。實驗中針對台灣南部地區屏東、高雄、台南等養殖場的南美白對蝦進 行採樣,以微孢子蟲EHP 專一性引子 ENF779/ENR779 及 ENF176/ENR176 分別進行 PCR 及 Nested PCR 的偵測,確認台灣養殖南美白對蝦確實受到 EHP 的感染,進而以
病理組織切片分析EHP 感染的組織趨性及病理變化,實驗中並將 PCR 反應增幅 EHP
的 SSU rDNA 部分基因片段(779 bp)進行基因轉殖、定序、序列的比對與親緣關係的
分析,同時利用PCR 的方式將此核酸片段進行 DIG 標識製成探針,建立點墨雜交法
(Dot blot hybridization)及原位雜交反應(In situ hybridization)檢驗 EHP 的偵測方法。實 驗結果顯示:在台南地區的南美白對蝦在 PCR 及 nested PCR 中,都有偵測到感染 EHP
VII
片的觀察,顯示南美白對蝦的肝胰臟組織可見上皮細胞有微孢子蟲的感染情況,同時
細胞質內呈現嗜酸性的顆粒,除了肝胰臟外的其他器官並無EHP 感染的情形。實驗
中EHP SSU rDNA 部分基因的基因序列定序、比對與親緣關係分析方面,顯示在台
灣所發現的EHP 與泰國、越南、印度、中國都有 99%以上的核酸相似性,因此應屬
同一種微孢子蟲,有關台灣的EHP 來源應進一步研究。在建立南美白對蝦 EHP 分子
生物的檢測方式方面,以點墨雜交法(Dot blot hybridization)檢測探針之靈敏度,顯示
只要13 pg/ul 的質體核酸即可呈現陽性反應,進而利用此方法檢測罹患 EHP 的南美
白對蝦所萃取出來的核酸,亦可呈現陽性反應而確認有受到EHP 的感染;原位雜交
反應(In situ hybridization)的檢驗,亦可成功的偵測到南美白對蝦的肝胰臟被 EHP 微 孢子蟲感染的現象,靈敏度優於傳統的組織切片。綜合本研究的結果:以 PCR、Nested PCR、組織病理切片、點墨雜交法(Dot blot hybridization)、原位雜交反應(In situ hybridization)等檢測方法,確認台灣養殖的南美白對蝦確實已受到 Enterocytozoon
hepatopenaei (EHP)微孢子蟲的感染,有關 EHP 的防治應進一步進行研究分析。
VIII
Establishment of diagnostic methods to detect
Enterocytozoon hepatopenaei (EHP) infection in culture
Litopenaeus vannamei in Taiwan
Advisor:Dr. Chun-Shum Wang Institute of Biotechnology National University of Kaohsiung
Student:Liu,Yu-Hsuan Institute of Biotechnology National University of Kaohsiung
ABSTRACT:
This study investigated the epizootic of Enterocytozoon hepatopenaei (EHP) infection from Litopenaeus vannamei cultured in Taiwan. In addition, the diagnostic methods for EHP infection were established using biotechnology. The diseased L. vannamei were collected in southern Taiwan, mainly in Pingtung, Kaohsiung and Tainan. The PCR and nested PCR were carried out with the specific primers (ENF779/ENR779 and ENF176/ENR176) in order to confirm the EHP infection. Furthermore, the tissues tropism and histopathological changes of EHP infection were evaluated by traditional histological method. The amplicon of EHP SSU rDNA partial gene fragments (779 bp) by PCR reaction were cloned, sequencing, and sequence alignment and phylogenetic analysis. The DIG-probe of EHP SSU rDNA partial gene fragments were prepared by PCR method. The dot blot hybridization and in situ
IX
hybridization detective methods for EHP were developed. The results showed EHP microsporidia were detected in L. vannamei in Tainan using PCR and Nested PCR. In Pingtung and Kaohsiung, EHP infection were confirmed by nested PCR. Histopathological study, the epithelial cells of hepatopancreas of L. vannamei were target tissue infected by EHP using H& E staining. Except for hepatopancreas, EHP infections were not observed in other organs in the diseased shrimp. The EHP SSU rDNA partial gene fragment of EHP isolated from Taiwan were found more than 99% nucleotide sequence similarities with isolates of Thailand, Vietnam, India, and China. The phylogenetic tree analysis, Taiwan isolate should be same as EHP isolated from L. vannamei cultured in Southeast Asia. The sensitivity of DIG-probe for EHP was 13 pg/ul using dot blot hybridization. By Dot blot hybridization, the EHP infection was detected in the diseased shrimp using the DNA extracted from EHP infected L. vannamei and confirmed it is suffered from EHP microsporidian. In situ hybridization test also successfully detected the EHP infection of the hepatopancreas of L. vannamei by DIG-probe. The sensitivity was excellent than that of traditional histopathology. Conclusion, the Enterocytozoon hepatopenaei (EHP) microsporidian infection was confirmed in the L. vannamei cultured in Taiwan using PCR, nested PCR, histopathology, dot blot hybridization, in situ hybridization methods. In future, the preventive and treatment methods for EHP should be further developed in Taiwan.
Keyword:Litopenaeus vannamei、Enterocytozoon hepatopenaei ( EHP )、PCR、Dot blot hybridization、In situ hybridization
1
第一章 前言
1.1 南美白對蝦的習性介紹
南美白對蝦(俗稱白蝦),學名:Litopenaeus vannamei,在分類學上屬 於節肢動物門(Phylum Arthropods)、甲殼綱(Superclass Crustacea)、十足目 (Decapoda)、對蝦科(Family Penaeidea) (李,2005)。南美白對蝦主要棲息 於中南美洲西部太平洋沿岸熱帶水域,自墨西哥南部至秘魯中部,以厄 瓜多爾沿岸分布最為集中。南美白對蝦為底棲性生物,分布於近岸水深 約0 ~ 200公尺的泥質大陸棚,成體蝦類多生活於沿岸水域,一到繁殖期 則會到河口進行繁殖,幼蝦則多生活於餌料豐富的河口地區生長,南美 白對蝦可接受的鹽度範圍很廣,在鹽度5 ~ 40 ppt皆可生長,甚至可以完 全淡水養殖,最適合鹽度為 25 ~ 30 ppt,南美白對蝦對溫度的適應性也 很強,在冬季15 ~ 18 ℃ 也可以生存,但成長率會降低,當溫度低於9℃ 以下則會死亡,最適合溫度為23 ~ 30℃。南美白對蝦是較偏屬於夜行性 生物,白天一般都會靜浮在海底,夕陽西下之後則活動力較為頻繁,南美 白對蝦脫殼時間大多都在晚上的時候進行,在進行脫殼的時候是南美白 對蝦最脆弱的時候,所以很容易發生互相殘食的情況(丁,2005)。南美白 對蝦於1985年引進台灣,剛開始交由高雄市養蝦業者及東港分所試養,
2
只可惜的是沒有養至收成,草蝦正值台灣養殖全盛期,所以就無人問津
了 , 由 於 草 蝦 在 1988 年 遭 遇 到 草 蝦 桿 狀 病 毒 感 染 (Penaeus
monodon,baculovirus : MBV),1992年又遭受白點病毒感染(White spot syndrome virus ; WSSV),使得台灣蝦類養殖遭到極大的打擊。南美白對 蝦於1995年再次進口無特定病原南美白對蝦種蝦,於台灣南部嚐試商業 化繁養殖,再於1998年起,由於其成長快速,收穫體型小,養成期間明顯 縮短,加上養殖技術進步,存活率高,使得養殖成效更佳,進而掀起台灣 地區南美白對蝦養殖的熱潮(陳弘成,2001)。南美白對蝦是商業養殖中被 眾多國家引進的水產養殖生物之一,以台灣地區來觀看,主要的養殖地 區分布以屏東、高雄、台南、嘉義等為主要的養殖地區。
1.2 南美白對蝦之常見的疾病
近年來由於台灣水產養殖之成敗受到了水質、寄生蟲、細菌及病毒 性疾病等因素影響,其中以病毒性的疾病防治最不易。在南美白對蝦的 病毒性疾病最主要以白點症(White Spot Syndrome,簡稱WSS)、傳染性及 皮下造血組織壞死病(Infectious Hypodermal and Hematopoietic Necrosis, 簡稱IHHN)、陶拉病(Taura syndrome virus,簡稱TS) (董明澄,2005)。白(Non-3 occluded Baculovirus),目前分類屬於Nimaviridae科之Whispovirus屬,為目 前嚴重危害台灣及其他亞洲地區甚至美洲地區之養殖蝦類,在台灣感染 包括白蝦、草蝦、班節蝦、砂蝦、紅尾蝦等,本病傳播迅速,死亡率最高 可達100%(台灣南區魚病中心等,1994)。陶拉病(TS)屬單股RNA之微小 RNA病毒,源自中南美,含美國佛州、德州、夏威夷之南美白對蝦,台灣 自1997 年中南美大量引進南美白對蝦後台灣已是TS之疫區(Tu et al., 1999; Yu and Song, 2000)。累計死亡率可達80 ~ 95%,但一般約有60%收 成 率 。 傳 染 性 皮 下 及 造 血 組 織 壞 死 病 (IHHN) 之 病 毒 屬 細 小 病 毒 (Parvovirus),單鏈DNA正20面體病毒(Bonami et al., 1990; Lightner et al., 1996),本病毒分布甚廣,美國、中南美、夏威夷、關島、大溪地、新加 坡、馬來西亞、泰國、印尼等均為疫區,台灣自1997年大量引進中南美白 蝦後亦成為疫區,白蝦感染引起慢性感染為主,感染蝦體型變形明顯,養 成池蝦大小參差不齊,產生許多超小體型蝦隻及畸形,其大小變異度達 30~50 %,造成經濟上損失,因蝦類仍有食慾而消耗飼料,但蝦隻不會相 對地正常長大(Bell and Lightner, 1984)。雖然病毒性疾病的對南美白對蝦 的傷害性很大,但細菌性、寄生蟲的疾病,例如弧菌及微孢子蟲感染都是 不容小覷(楊等,2016)。
4
1.3 微孢子蟲的特性
微孢子蟲(Microsporidea)為真菌界,羅茲菌門(Rozellomycota),羅 茲菌亞門(Rozellomycotina),微孢子蟲綱(Microsporidea)。微孢子蟲能寄生 於動物宿主,大部分的動物物種都可以被微孢子蟲寄生,包括人類。多數 微 孢 子 蟲 感 染 昆 蟲 , 也 是 甲 殼 類 及 魚 類 的 常 見 疾 病 (Patrick J., Keeling ., 2004)。在19世紀時,在家蠶中發現第一個微孢子蟲(Nosema
bombycis) (Nägeli KW., 1857)。至今微孢子蟲(Nosema bombycis)仍然是一
種致命的疾病,被稱為Pébrine,導致養蠶業的經濟損失(Bhat SA. 2009; Becnel JJ. 2014)。在魚類中,微孢子蟲的感染最常見的就是凹凸病以及黏 液微孢子蟲病,凹凸病是因微孢子蟲(Plistophora anguliiarum)寄生於魚類 肌肉所引起的疾病,主要感染鰻魚及香魚 ; 黏液微孢子蟲(Myxidium sp.) 寄生於眼睛、鰓部、外表皮膚以及鰭部,主要感染鰻魚以及淡水魚類(黃 世鈴和陳秀男,2006)。據報導,有幾屬主要以甲殼類為宿主的微孢子蟲, 如: Agmasoma,Ameson,Nosema,Pleistophora,Tuzetia,Thelohania,
Flabelliforma,Vavraia Ordospora
,
Nadelspora和Enterospora (Landgdon,1991; Larsson et al., 1996 ; Larsson et al., 1997 ; Larsson et al., 1998; Lightner, 1996; Canning et al., 2002 ; Refardt et al., 2002; Moodie et al., 2003; Amogan
et al., 2006; Stentiford and Bateman, 2007)。報導指出很少有微孢子蟲僅感
染甲殼類肝胰腺的小管上皮細胞(Anderson et. al., 1989; Hudson et. al., 2001;Wang and Chen., 2007;Stentiford and Bateman., 2007; Stentiford et
al., 2007)。Stentiford等人(2007),描述了兩種歐洲螃蟹物種的感染情況,
5 開啟了甲殼類動物感染腸球菌的可能性。在2003年泰國草蝦的養殖爆發 出生長綜合症(MSGS),但原因不明(Chayaburakul., 2004)。Tourtip等人 (2009),發現了新型的微孢子蟲,因新型微孢子的超微結構特徵,將其歸 類 在Enterocytozoon 屬 ,應 將 此 新 型 微 孢 子蟲 命 名 為
Enterocytozoon
hepatopenaei (EHP)。
1.4 Enterocytozoon hepatopenaei (EHP)的特性與歸類
微孢子蟲Enterocytozoon hepatopenaei(EHP)是一種細胞內形成孢子 的寄生蟲,它在肝胰腺中的小管上皮細胞的細胞質區域內複製(Tourtip et al., 2009)。微孢子蟲在細胞內會有不同的生長階段,多核正交瘧原蟲 (Plasmodia)與宿主細胞的細胞質直接接觸,並在表面含有大量的小泡。在 早期瘧原蟲發育期間發生了瘧原蟲核(Plasmodia)的二元裂變,並且在瘧 原蟲(Plasmodia)內形成了許多孢子細胞。緻密盤和極性小管的前體在早 期成孢子細胞從瘧原蟲(Plasmodia)表面出芽之前在瘧原蟲(Plasmodia)的 細胞質中發展。成熟的孢子是橢圓形的,尺寸為0.7×1.1 μm並且包含單核, 5-6個極性細絲線圈,後部液泡,附著到極性細絲的錨定盤和厚的緻密壁。 而緻密壁是由內生孢與外生孢所形成的(Tang et al.,2015)。EHP微孢子蟲 雖然歸類於Enterocytozoon屬,因其結構獨特,但於Enterocytozoon屬中的
6
來源可能是鳥類或哺乳動物 ,主要感 染水源為淡水而不是海水, 而
Enterocytozoon hepatopenaei (EHP)主要感染的是水生生物,主要為海水的
蝦類(Tourtip et al., 2009;Hayman et al., 2005)。
1.5
Enterocytozoon hepatopenaei (EHP)之地理分布及症狀
目前,已知EHP在亞洲廣泛存在,例如泰國,中國,印度,越南,等 國家(Ha et al.,2010)。而在泰國,EHP現在已引發白斑病毒引起的白斑病 (WSD)和弧菌引起的急性肝胰臟壞死病(AHPND)第三種嚴重的養殖蝦 類疾病,EHP對全球對蝦類產業已構成一定的威脅性。因此迫切需要更好 地了解其感染機制和寄生蟲發病機制,以促進預防和治療策略的發展。 近幾年來EHP是新興的微孢子蟲,而關於EHP如何與宿主相互作用的知識 仍然知之甚少(Tang et al., 2015; Rajendran et al., 2016)。已知感染EHP的蝦 類會有嚴重的生長遲緩進而影響蝦類養殖產量(Newman, 2015)。Flegel (2012)表明,在越南和泰國發現的南美白對蝦的白糞綜合症(White Feces Syndrome ,簡稱WFS)與嚴重感染微孢子蟲有關,而此微孢子蟲 的形態與EHP相似。Tangprasittipap等人(2013)報導指出在泰國微孢子蟲 廣泛感染南美白對蝦並出現WFS症狀,但蝦類卻沒有檢測出感染EHP微 孢子蟲,所以推測感染EHP不是南美白對蝦併發WFS的原因。Aranguren
7
等 人(2017) 報 導 , EHP 微 孢 子 蟲 感 染 會 惡 化 成 Acute hepatopancreatic
necrosis disease (AHPND)導致引發弧菌感染造成Septic hepatopancreatic necrosis (SHPN)。由於EHP微孢子蟲與AHPND都會導致上皮細胞從基底 膜脫落,在此種情況下,兩者共同感染明顯增加對肝胰腺的損害,而導致 全組織的發病;EHP微孢子蟲會增加蝦類對弧菌的易感性,而導致SHPN 的發病現象,這個發現可能是全球蝦類水產養殖中疾病爆發的模式。但 儘管EHP微孢子蟲尚未被科學證實有死亡原因,但野外觀察表明EHP微孢 子蟲的感染與嚴重生長遲緩有一定的相關性(Sritunyalucksana et al., 2014; Chaweepack et al., 2015)
1.6
Enterocytozoon hepatopenaei (EHP)傳播方式及感染途徑
Enterocytozoon hepatopenaei (EHP)是一種新型的微孢子蟲,近幾年
發現而好發於養殖的南美白對蝦,所以EHP的感染與傳播方式是大家所 特別關注的。Salchan等人,在2017年做一個有關EHP的水平感染實驗報 告。Salchan 等人,主要是把受感染的南美白對蝦(放養6隻)與未受感染的 南美白對蝦(放養6隻)一起飼養在同一個水族缸中,而受感染的南美白對 蝦放置在網狀籠子裡,再放置水族缸中飼養,未受感染的南美白對蝦則 是放養在水族缸中,以這樣的方式來測試EHP是否有水平感染的可能性。 放養14天後,使用PCR做為測試EHP是否有感染,在網狀籠子裡取感染南
8
美白對蝦3隻,再取放養在水族缸中原來未受感染的南美白對蝦3隻進行 PCR測試。實驗結果,發現原本未受感染的南美白對蝦經14天,確定已被 網狀籠子裡的南美白對蝦傳染。此傳染的南美白對蝦進行病理組織切片 及原位雜交(In situ hybridization),若可以清楚的檢測到EHP微孢子蟲的存 在。研究報告證實了EHP的感染與傳播途徑是可以藉由水平的方式感染 南美白對蝦。
1.7 Enterocytozoon hepatopenaei (EHP)的檢測方法
近幾年泰國、中國、越南、印度等地區爆發新型的微孢子蟲,亞洲 各國陸續有相似的病情報導,由於此微孢子蟲疾病無法治療,只能靠一 些較快速的方法來診斷預防以及控制疾病的發生,目前已有幾種方法可 應用於偵測蝦類微孢子蟲EHP,分述如下:
1.7.1 外表病徵觀察
感染EHP的蝦類在外觀上並不會有明顯的變化,如要預測是否有感 染EHP只有觀察生長速度是否遲緩(Suebsing et. al., 2013)。感染EHP的蝦 類與感染其他的微孢子蟲有很大的不同在於有的微孢子會寄生在肌肉而 使蝦子的腹部肌肉呈現白濁,例如微孢子蟲中的Tuzetia weidneri,外表病
9 徵觀察這方法雖簡便,但要利用此方法來判斷蝦類是否有感染EHP是有 一定困難度的,只能以活動力較低的養殖蝦類來猜測可能有感染微孢子 蟲EHP的可能性。
1.7.2 電子顯微鏡技術
使用電子顯微鏡可以觀察病毒的型態,是研究病毒感染位置最好的 方法。利用電子顯微鏡觀察到南美白對蝦感染EHP微孢子蟲,在細胞質中, 微孢子蟲的大小約為290-300 nm (Somjintana et al., 2009)。Paul等人以電 子顯微鏡的技術觀察到南美白對蝦感染EHP微孢子蟲在細胞質中含有大 量的微孢子蟲顆粒(Paul et al., 2017)。1.7.3 組織病理切片
此方法適用於魚病,而蝦類的病理也是可以運用此方法來觀察病理 變化,此方法較為傳統的診斷法,即是將蝦類以固定液固定後,進行包 埋、切片及染色,最後在光學顯微鏡下觀察病理變化,蝦類在罹患EHP時, 可見 EHP 的微孢子會存在於肝胰臟中(Rajendran et al., 2016)。10
1.7.4 DNA 探針
利用核酸一段基因,製作探針,再配合酵素呈色反應或是螢光染劑, 來檢驗南美白對蝦病毒。其靈敏度可達到1 pg以下的核酸,但缺點是處理 時間較長且技術高。DNA探針可以衍生的方法有Dot blot hybridization以 及In situ hybridization 。 將 EHP 部 分 核 酸 序 列 製 成 探 針 , 以 In situ
hybridization的方法對蝦病組織切片進行檢測觀察,結果顯示EHP微孢子 蟲主要分布於南美白對蝦的肝臟中(Taugprasittipap et al., 2013)。
1.7.5 聚合鏈連鎖反應( Polymerase chain reaction,PCR )
PCR技術目前已被廣泛的運用於人類及動物的病毒性及細菌性、寄 生蟲性的檢測上,對於特定的基因之存在與否及演化關係上,都有重大 的發展。目前很多使用此方法進行微孢子蟲檢測的文獻(Kathv et al.,2017; Rajendran et al.,2016)。
1.7.6 環介導的等溫擴增(loop-mediated isothermal
amplification,LAMP)
loop-mediated isothermal amplification (LAMP)是用於核酸增幅技術,
11
板(heat block)就可以使用(Notomi et al., 2000)。Sathish等人(2018)展示 了 一 種 非 常 簡 單 又 快 速 , 特 異 和 閉 合 的 管 視 覺 LAMP 測 試 , 使 用 SYBR™green I染料檢測EHP微孢子蟲。此方法具有許多優點,例如高特 異性和靈敏度,在等溫條件下可以進行測試,現場適應性,並且不需要復 雜的設備(Notomi et al. 2000)。
1.8 實驗目的
本實驗擬探討台灣養殖南美白對蝦微孢子蟲EHP 之特性,並利用已 知的EHP 專一性引子,將檢體以 PCR 放大 EHP 微孢子蟲核酸進行確認, 再進行基因選殖、核酸定序和序列分析比對,以了解微孢子蟲 EHP 在各 國的EHP 微孢子蟲的親緣關係。實驗中分別利用病理組織切片觀察、PCR檢 測 、 點 墨 雜 交 反 應(Dot blot hybridization) 、 原 位 雜 交 反 應 In situ
hybridization)檢測等分子生物學的技術建立南美白對蝦感染微孢子蟲 EHP 的偵測方法,藉此可提供未來選擇適合之檢測 EHP 微孢子的方式。
12
第二章 材料與方法
2.1 南美白對蝦微孢子 EHP 病學之研究
2.1.1 採樣
南美白對蝦主要採樣地點為屏東地區以及台南地區的蝦類養殖場, 採集體型較小的蝦子,用碎冰冰藏之,帶回實驗室進行檢測。
2.1.2 病理組織切片觀察
2.1.2.1 固定、脫水、包埋及切片
將採集蝦類的肝胰臟,以Davidson 溶液固定 24-48 小時,再依序以 酒精70% → 80% → 90% → 95% →100% 進行脫水,每次一小時,接著 以二甲苯反應一小時,再放入含少量二甲苯的石蠟中(溫度 62℃)一小時, 再移入只有石蠟(溫度 62℃)的浸潤槽一小時後,即進行包埋,而後將包 埋完成的組織塊,利用迴轉式切片機將組織切成厚度5-7 μm 的薄片,切 完放置水中展開,展開後置於含蛋清的載玻片上,等待日後染色。13
2.1.2.2 染色
2.1.2.2.1 蘇木精和伊紅(Hematoxylin and Eosin)染色法
利用二甲苯浸潤玻片2 次,每次 3 分鐘;再依序以酒精 100% → 90%
→ 80% → 70% → 50% 進行水合,每次 3 分鐘,再以水浸泡 1 分鐘。將 組織切片先以蘇木精(Hematoxylin)染劑,染 15 分鐘。在浸於 Scott solution
作用3 分鐘。以自來水沖洗 1 分鐘後,以酒精 50% → 70% 進行脫水,
每次30 秒後,置於 0.5% 伊紅(Eosin)溶液中,作用 5 分鐘,依序以酒精
80% → 90%→ 95% → 100% →100% 作用各 15 秒,在置於二甲苯溶劑中 2 次,每次 3 分鐘,利用 Entellan 封片,至於光學顯微鏡(Olympus IX50) 下觀察,並照相。
2.2 EHP 之部分基因片段序列選殖
2.2.1 去氧核醣核酸的(Total DNA)萃取
主要參考Sambrook 等人(1989)的方法,將蝦子的肝胰臟取出,以 Lysis
buffer(3% SDS; 10 mM EDTA; 10 mM Tris-HCL pH 8.0;0.1 mg/ml proteinase K)混合研磨後,置入 65℃中水浴 3 小時,而後加入等量的飽和 (Saturated phenol)混合均勻後,以 5000 xg (Hitachi CT 15RE)離心 5 分鐘, 抽取上層液,加入等量的飽和酚和氯仿混合液(Phenol and Chloroform),
14
在 4℃ 5000 xg 離心 5 鐘 , 抽取 上 層液 , 將上 層 液 與等 量 的氯 仿
(Chloroform)混合後,在 4℃ 5000 xg 離心 5 分鐘,抽取上層液。並加入 2
倍體積的Isopropanol 及 1/10 體積的 3 M 醋酸鈉(Sodium acetate),存放於
-20℃,冰藏 16-18 個小時。以 4℃ 12,000 xg 離心 5 分鐘後,倒掉上清液, 置於 70℃中 20 分鐘後,將沉澱物懸浮於 50μL 的二次蒸餾水中,置於-20℃中。
2.2.2 聚合酶連鎖反應(PCR)
以1 μL 的蝦子核酸做為模板,加入 PCR 專用的微量離心管中,此離 心管包括物質:10 倍的 PCR 緩衝溶液(附錄)、10 mM dNTPs、1μL 的 EHP 專一性引子( ENF779:5`-CAGCAGGCGCGAAAATTGTCCA-3`;ENR779: 5`-AAGAGATATTGTATTGCGCTTGCTG-3`及 ENF176: CAACGCGGGAAAACTTACCA ; ENR176 : ACCTGTTATTGCCTTCTCCCTC)
(Tangprasittipap et al., 2013)及 0.5 單位的 Taq DNA 聚合酶,共 60 μL。將 反應液置於 PCR 機器中(Bio-rad Thermal Cycler)進行反應,反應程式為 (94℃ 3 分鐘,而後進行 94℃ 20 秒、54℃ 30 秒、72℃ 45 秒;共 40 個循
環,最後以72℃ 5 分鐘、4℃ 5 分鐘,結束 PCR 反應)。在聚合酶連鎖反
15
2.2.3 膠體純析(Gel elution)
將約779 bp 的 EHP 病毒部分基因片段序列自 0.8%的洋菜膠體中切
下,利用Wizard SV Gel and PCR Clean-Up System(Promega)來進行純化的
工作。最後將EHP 微孢子蟲之部分基因片段序列沉澱物懸浮於 50 μL 的
二次蒸餾水中,直接進行以下實驗。
2.2.4 DNA 接合(DNA Ligation)
取純化後1μL 約 779 bp 的 EHP 病毒部分基因片段序列,加入到 1.5
ml 微量離心管中混合包括:0.5 μL 的 2 倍 Ligase 緩衝溶液、0.5 μL 的 pGEM-Easy 載體 DNA、2 μL 的 T4 DNA ligase,加入 2 μL 蒸餾水使反應
總體積為5 μL,於 14℃下反應 4 小時以上,反應完後直接進行以下實驗。
2.2.5 質體轉型(Transformation)
將勝任細胞(JM109)與 Ligation 好的 DNA 產物混合,先置於 4℃,20 分鐘後,以42℃加熱 45 秒,立即置於 4℃ 2 分鐘,加入 LB 培養液,於 37℃下旋轉 1.5 小時後,加入 200 mg/ml IPTG 4 μL 及 20 mg/ml X-gal 40 μL 混和均勻後,塗抹於含 Ampicillin(50 μg/ml)的 LB 培養基上,於 37℃ 恆溫箱中培養16-24 小時。16
2.2.6 質體抽取(Plasmid extraction)
篩選LB 培養基上的白色菌落,接種於 3 ml 的 LB-ampicillin (50 μg/ml)
培養液中,於37℃下旋轉培養 14-16 小時後,以 10000 xg 離心 5 分鐘,
將細菌沉澱下來,加入 Pure YieldTM Plasmid Miniprep System (Promega)
來進行抽取質體DNA 的工作,最後將重組質體之沉澱溶解於 100 μL 的
二次蒸餾水中,即得純化的重組質體。
2.2.7 限制酶素切割(Restriction enzyme digestion)
將上述所得的重組質體混合限制酶EcoR I,在 37℃下反應 3 小時後,
將產物以0.8%洋菜膠電泳分析,確認所插入的病毒部分基因片段序列長
度後,再將質體DNA 送至民間生技公司(基龍米克斯)進行核酸定序。
2.2.8 核酸序列之分析與比對
將選質到 EHP 病毒之 SSU rDNA 片段基因序列,利用生物資訊
MEGA7 軟體及 NCBI 網站(Nucleotide-nucleotide BLAST)進行排序以及序 列相似性比對,進行親緣關係分析(Saitou N. and Nei M., 1987 ., Tamura K., Nei M. and Kumar S., 2004)。
17
2.3 EHP 微孢子蟲之檢測方式與分析
2.3.1 點墨雜交(Dot blot Hybridization)
2.3.1.1 探針之製備與純化
以PCR 的方法將探針進行 DIG 標定。將 1 ng 含 EHP 微孢子蟲的質
體為模板,加入PCR 專用的微量離心管中,此離心管包括物質為:10 倍
的PCR 緩衝溶液 (附錄)、5 μL 的 DIG-label dUTP、1 μL 的 EHP 微孢子
蟲專一性引子(ENF779:CAGCAGGCGCGAAAATTGTCCA、ENR779: AAGAGA TATTGTATTGCGCTTGCTG)及 0.5 單位的 Taq DNA 聚合酶,
最後加水至50 μL。將反應液置於 PCR 機器中(Thermal Cycler)進行反應, 設定執行程式(94℃,5 分鐘,而後進行 94℃ 1 分鐘、54℃1 分鐘、72℃ 1 分鐘;共 40 個循環,最後以 72℃ 5 分鐘、4℃ 5 分鐘,結束 PCR 反 應)。在聚合酶連鎖反應後,進行洋菜膠體電泳分析,將約 779 bp 的 EHP 微孢子蟲部分基因片段序列自0.8%的洋菜膠體中切下,利用上述 2.2.3 步 驟來純化探針。
18
2.3.1.2 點墨雜交反應(Dot blot Hybridization)
將不同來源的 DNA(蝦類組織 DNA 及含已選殖之 EHP 微孢子蟲部
分基因片段序列的重組質體)95%變性 10 分鐘,置於 4℃ 1 分鐘,取 1 μL
滴 於 Nylon membrane 格 子 上 , 再 利 用 UV crosslinker (Spectronic
corporation)做 crosslink 2 次。待 Membrane 風乾後,加入 Pre-hybridization Solution (5X SSC ( 附 錄 ) 、 1% Block reagent buffer ( 附 錄 ) 、 0.1% N-lauroylsarco sin、0.02% SDS、500ng DNA probe),42℃下反應 16-18 小時。
用2X SSC 洗 2 次(每次 5 分鐘,42℃)後,再以 0.1X SSC 替換洗 2 次(每
次15 分鐘,42℃),以 Buffer 1(附錄)洗 1 分鐘。加入 Buffer 2(附錄)在溫
室下作用30 分鐘。加入以 1/5000 Alkaline phosphatease 連結之抗 DIG 抗
體,於室溫下作用30 分鐘。以 Buffer 1 洗 2 次(每次 15 分鐘)。Buffer 3(附
錄)洗 1 次(2 分鐘)。而後以含 2%呈色液之 NBT 的 Buffer3(附錄),於黑
暗中呈色12 小時。最後以 Buffer 1 終止反應,進行觀察。
2.3.2 原位雜交反應(In situ hybridization)
將南美白對蝦組織玻片浸於二甲苯(Xylene)中脫蠟 3 分鐘,依序以
酒精進行水合,各3 分鐘。滴入 Proteinase K(100 μg/ml)20-30 μL 於玻片
上,37℃反應 10-15 分鐘。加入 0.4% Formaldehyde,4℃固定 5 分鐘後,
19
formaldehyde、1 μg/ml DIG 標識的探針、5 mg/ml sonicated salmon sperm DNA、1% Dextran sulfate、4X SSC),於 95℃下作用 10 分鐘。置於 4℃ 1
分鐘,將玻片置於 Humid chamber 上,42℃作用 3 小時。以 2X SSC 5 分
鐘,洗2 次;0.1X SSC 10 分鐘,42℃ 1 次。再以 Buffer 1 洗 2 分鐘,加
入500 倍稀釋含 Alkaline phosphatease 連結的抗 DIG 抗體。室溫反應 1
小時後,用 Buffer 1 洗 10 分鐘,2 次後再以 Buffer 3(附錄)洗,而後以含
2%呈色液 NBT/BCIP 的 Buffer 3,於黑暗反應 12 小時,呈色後的切片以 酒精脫水,Eosin Y 作比對染色,再以 Entellan 封片,將完成之組織切片 以光學顯微鏡(Olympus IX50)進行觀察、拍照。
20
第三章 結果
3.1 南美白對蝦 EHP 微孢子蟲流行病學之研究
實驗中所採集的南美白對蝦於高雄、屏東以及台南地區的養殖場疑 似罹患EHP 微孢子蟲的病蝦,進行採樣。隨機抽取三隻南美白對蝦的肝 胰 臟 萃 取 核 酸 , 並 利 用 EHP 專 一 性 引 子 (ENF779/ENR779 ; ENF176/ENR176)進行聚合酶連鎖反應(PCR)檢驗確認是否受到 EHP 微孢 子蟲的感染,而檢測病例結果如表 1 所示,可發現在台灣屏東、高雄以 及台南地區所養殖的南美白對蝦(編號 1.2.3.4)等 12 個檢體,引子 779 bp 在高雄以及屏東所檢測結果,呈現陰性反應,相反之在台南地區的南美 白對蝦,呈現陽性反應(圖 1),再者使用 Nested PCR 在高雄、屏東以及台 南均呈現陽性反應(圖 2),進而更加確認高雄、屏東以及台南地區的養殖 南美白對蝦都有感染的現象,此外在台南地區有檢驗草蝦是否有感染 EHP 微孢子的現象,使用了 EHP 專一性引子做檢測,引子 779 bp 呈現陰 性反應,再者使用Nested PCR 有呈現陽性反應,可推測草蝦也有可能受 到 EHP 微孢子蟲感染情形。而在台南所採集的南美白對蝦為約放養 20 天,即可感染EHP 微孢子蟲。需更進一步的以基因排序檢驗確認 EHP 微 孢子蟲之特性。在養殖池,觀察一般罹患 EHP 微孢子蟲的南美白對蝦, 除了體型較小之外,在外表其他部位並無任何受損之症狀出現。21
3.2 組織病理觀察
將採集所得EHP 微孢子的南美白對蝦,頭部肝胰臟的部份取出出來
進行石蠟包埋切片,以蘇木精及伊紅
(Hematoxylin and Eosin)
染色,觀察南美白對蝦在罹患EHP 微孢子蟲後的病理變化,結果發現南美白對蝦 的肝胰臟組織中有EHP 微孢子蟲在肝胰臟呈現嗜酸性的胞液體的分布情 形,在其他組織沒有發現(圖 3(A.B.C.D))。
3.3 EHP 微孢子蟲片段基因的選殖
3.3.1 SSU rDNA 之片段基因 PCR 轉質和序列分析
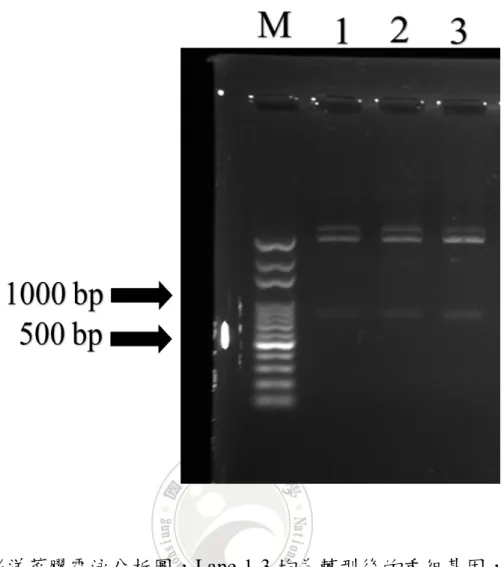
將以確認感染EHP 微孢子蟲的檢體選取台南所採樣的 3 組南美白對 蝦,分別從組織中萃取 DNA,利用 EHP 專一性引子以 PCR 反應進行擴 增,經0.8%的洋菜膠體電泳分析,結果擴增出與預計大小相符的 776 bp單一核酸片段。將此條帶切下,再依 Wizard SV Gel and PCR Clean-UP
System (Promega)來回收純化此 DNA 片段,然後以 TA cloning 的方式,
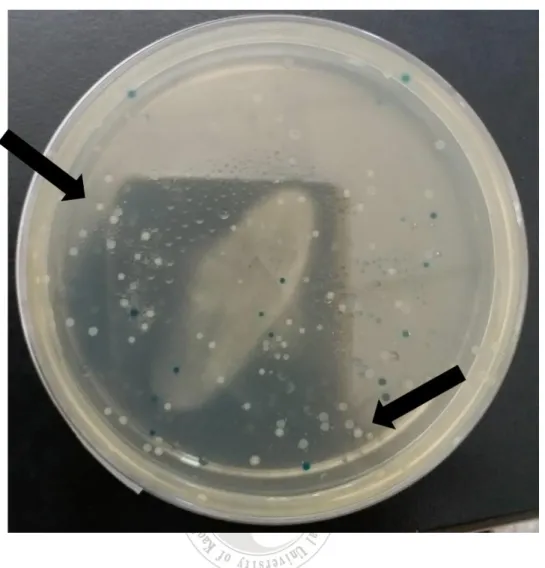
植入到pGEM-T Easy 載體中,而後經轉型作用(Transformation)到大腸桿
菌 JM109 中(圖 4)。抽取重組質體後,經 EcoR I 切割,得到相對應大小 的片段(圖 5),證明重組載體含有 EHP 微孢子蟲片段基因,進而進行定序
22
及比對的工作。
3.3.2 EHP 微孢子蟲 SSU RNA 片段基因序列比對
將重組質體基因經定序後,所得 EHP 微孢子蟲核酸序列長度為 779 bp。序列比與 NCBI 基因資料庫(Database)內已知基因序列進行比較,如 圖6、圖 7 所示,顯示 EHP 微孢子蟲的核酸序列與其他 EHP 微孢子蟲之 SSU rDNA 部份基因序列有很高的相似性至少達百分之 99%以上,其中 與編號 KF362129.1(來自泰國之 EHP 微孢子蟲)相似度均達 99%;編號 MH260591.1( 來 自 印 度 之 EHP 微 孢 子 蟲 ) 相 似 性 為 99.87% ; 編 號 MF134829( 來 自 越 南 之 EHP 微 孢 子 蟲 ) 相 似 性 為 99.87% ; 編 號 KY593127(來自美國之 EHP 微孢子蟲)相似性為 98.75%;與來自拉丁美 洲之南美白對蝦相似性為 99.77%;編號 MF417474(來自中國之 EHP 微 孢子蟲)相似性為 99.22%;編號 KF135642(來自西班牙之 EHP 微孢子蟲) 相似性為92.75%。以上的基因序列分析結果可發現來自不同國家所發現 的 EHP 微孢子蟲與本實驗中的 EHP 微孢子有很高的相似性,進而可以 推論泰國、印度、越南、中國,甚至是美國與拉丁美洲所發現的EHP 微 孢子蟲與台灣所發現的 EHP 微孢子蟲之間的基因序列有很高的相似性。
23
3.4 EHP 微孢子蟲之檢測方式之建立與分析
3.4.1 點墨雜交試驗(Dot blot hybridization)
利用ENF779/ENR779 引子對,以 PCR 反應的方式,將 EHP 微孢子
蟲 SSU rDNA 之片段基因以 DIC 標識製成病毒的檢驗探針,結果如圖 8
所示。在探針評估此探針之靈敏度,以Dot blot hybridization 的方式,分
別利用已建構完成之不同重組質體,直接去進行DNA 與 DNA 間的相互
雜交試驗,結果顯示此探針能專一與重組基因質體做結合靈敏度約 13
pg(圖 9)。實驗進一步以南美白對蝦所萃取之核酸直接進行檢測,檢測是
否將核酸萃取出來直接進行Dot blot hybridization ,以更快速的方法檢測
到南美白對蝦是否有感染到 EHP 微孢子蟲,結果如圖 10 所示,確定感
染之2 個檢體均呈現陽性反應。
3.4.2 原位雜交試驗(In situ hybridization)
利用原位雜交的方法,檢驗EHP 微孢子蟲在南美白對蝦的肝胰臟中
分布的情形,結果顯示(圖 11(A、B))以 DIG 標識的探針,能偵測到肝胰
臟 EHP 微孢子蟲在細胞內的感染,可發現其他組織無法偵測到 EHP 微
孢子蟲,EHP 微孢子蟲只感染肝胰臟,所以此呈色所分布於組織間的程 度來判斷感染的嚴重性。
24 表 1 台灣南部地區養殖南美白對蝦採樣紀錄表 採樣器官皆為肝胰臟 序號 來源 蝦種 採樣 時間 蝦體重量 EHP 微孢子蟲偵測 PCR Nested PCR In situ hybridization 1 屏東 南美白對蝦 2018.9 5.0g 0/3 3/3
-
2 高雄 南美白對蝦 2018.11 10g 0/3 3/3-
3 台南 南美白對蝦 2018.12 3g 3/3 3/3+
4 台南 南美白對蝦 2019.5 0.5g 0/3 2/3 - 5 台南 草蝦 2019.5 0.5g 0/3 3/3 -25
26
圖 1 利 用 0.8% 洋 菜 膠 電 泳 圖 , 使 用 EHP 微 孢 子 蟲 專 一 性 引 子
ENR779/ENF779 偵測,檢測到南美白對蝦感染 EHP 微孢子蟲。M 為 100 bp ladder DNA。
27
圖 2 0.8%洋菜膠電泳圖,使用 Nested PCR 偵測,更加確定南美白對蝦
28 圖 3 EHP 微孢子蟲感染南美白對蝦肝胰臟病理切片圖 A 跟 C 圖為感染 EHP 微孢子蟲之白蝦肝胰臟,肝胰臟組織中分布 EHP 微孢子蟲(框 起來的位子)。B 圖是 A 圖肝胰臟感染範圍放大中的 EHP 微孢子蟲 的分布情形。D 圖為 C 圖肝胰臟感染範圍放大中的 EHP 微孢子蟲 分布情形。
29
圖 4 pGEM-T Easy 載體回收純化 DNA 片段,以 TA cloning 的方式,植
入到pGEM-T Easy 載體中,經轉型作用(Transformation)到大腸桿菌
JM109 中,白色是成功轉型到大腸桿菌 JM109 的菌株,藍色則是未 成功轉型的菌株。
30
圖 5 0.8%洋菜膠電泳分析圖,Lane 1-3 均為轉型後的重組基因,經限制
酶切割後,顯示重組基因含EHP 微孢子蟲片段基因(約 779 bp)。M
34
圖 6 EHP 微孢子蟲核酸定序與其他國家 EHP 微孢子蟲核酸序列之比對
圖:將 EHP 微孢子蟲 SSU rDNA 基因序列與泰國、印度、越南、
中國、美國、拉丁美洲等國家的 EHP 微孢子蟲基因序列進行比對,
35
圖 7 基因親緣關係比較圖 EHP 微孢子蟲 SSU rDNA 片段基因親緣關係之
比較,台灣 EHP 微孢子蟲與泰國、印度、美國、中國的親緣關係比
36
圖 8 0.8 %洋菜膠電泳分析圖,利用 PCR 的方法將探針進行 DIG 標定。 Land 1-2 均為經 DIG 標識之 ATPase 之部分基因片段序列探針。M
37
圖 9 點墨雜交法(Dot blot hybridization)之靈敏度試驗圖,將 DIG 標識的 EHP 微孢子蟲探針,利用已建構完成之南美白對蝦兩組重組質體進
行之靈敏度測試,從圖片中可以看見含量高於 13 pg/μL 的 EHP 微
38
圖 10 利用點墨雜交法(Dot blot hybridization)對南美白對蝦進行組織 DNA 的偵測,從南美白對蝦所萃取之 DNA 進行檢測。結果可檢
39
圖 11 利用原位雜交法(In situ hybridization),以 DIG 標識的 EHP 微孢子
蟲探針,偵測EHP 微孢子蟲在南美白對蝦中分布的情形,藍色沉
澱表示陽性反應,A 圖病蝦的肝胰臟組織被 EHP 微孢子蟲感染, 呈現陽性反應,B 圖為 A 圖的放大圖,亦可發現組織中微量感染 可被偵測出來(箭頭)。
40
第四章 討論
自2003 年,泰國斑節對蝦(Penaeus monodon)的養殖業受到了生長綜
合症(MSGS)的影響(Chayaburakul et al.,2004;Flegel et al.,2004),在土池 飼養的斑節蝦生長速度大幅降低,導致明顯的經濟損失,但引起生長綜
合 症(MSGS) 的 病 因 卻 不 清 楚 也 尚 未 闡 明 (Chayaburakul et al., 2004;
Anantasomboon et al., 2006)。但值得關注的是,在斑節對蝦的淋巴器官中 觀 察 到 多 個 球 狀 體 並 且 顯 示 含 有 已 知 和 未 知 類 型 的 微 孢 子 蟲 (Anantasomboon et al., 2006;Srtunyalucksana et al., 2006)。於 2009 年,從
泰國斑節蝦中鑑定出Enterocytozoon 屬,但此微孢子蟲與 Enterocytozoon
屬中的微孢子蟲E.beiniusi 的 SSU rRNA 基因序列相對性低,進而新命名
Enterocytozoon hepatopenaei(EHP) (Tourtip et al., 2009)。
在2003 年至 2009 年期間,因不清楚微孢子蟲 EHP 對蝦類養殖的影
響以及危害,所以在很大的程度上被忽略了,也因為EHP 並不構成蝦類
養殖上的主要威脅(Tang et al.,2016)。然而,在近幾年蝦類養殖中,已經
嚴重被微孢子蟲 EHP 普遍感染了,而最主要影響到中國、越南、泰國、
印度、馬來西亞等東南亞國家(Ha et al.,2010;Tang et al.,2015;Rajendran
et al.,2016)。在台灣地區,雖然還沒有爆發感染微孢子蟲 EHP 的消息,
41 高雄、台南是否有嚴重感染微孢子蟲 EHP 的跡象。 PCR 技術是目前使用最廣泛、檢測快速及靈敏的檢測方式,檢測時 通常需要高專一性的引子來進行分析,雖然微孢子蟲的疾病很容易因為 突變而發生基因序列的改變,但是微孢子蟲之間通常含有相當高的相似 性 和 保 守 性 的 基 因 序 列 。 本 實 驗 所 利 用 ENF779/ENR779 以 及 ENF176/ENR176 等兩組 EHP 微孢子蟲專一性的引子對進行 PCR 及 Nested PCR,經 PCR 反應進行檢測結果 ENF779/ENR779 引子對只能檢 測是否有感染現象,如果南美白對蝦感染情形沒有很嚴重的情況下, ENF779/ENR779 引子對在 PCR 中是檢測不到的,需要再利用 Nested PCR 做更進一步的檢測,才會偵測到EHP 微孢子蟲是否有真正感染南美白對 蝦的情形,在本實驗中利用ENF779/ENR779 引子對在屏東以及高雄所採 樣的白蝦中沒有檢測出來,需再利用ENF176/ENR176 引子對做更進一步 的檢測,才可偵測到EHP 微孢子蟲的感染,這說明屏東以及高雄地區感 染 EHP 微孢子蟲的嚴重性較低,而 ENF779/ENR779 引子對在台南所採 樣的白蝦中有檢測出 EHP 微孢子蟲的感染,再利用 ENF176/ENR176 引 子 對 做 更 進 一 步 的 確 認 , 結 果 ENF779/ENR779 引 子 對 以 及 用 ENF176/ENR176 引子都有檢測到南美白對蝦感染 EHP 微孢子蟲,這說 明台南、屏東、高雄地區均有感染EHP 微孢子蟲的現象。
42
據報導,將 Enterocytozoon hepatopenaei (EHP)與不同種類的微孢子
蟲做部分rDNA 的基因序列,來比較出 Enterocytozoon hepatopenaei (EHP)
與不同種類的微孢子蟲是否有親緣上的相關性。Tourtip 等人(2009),將來
自泰國對蝦感染的微孢子蟲 Agmasoma penaei 的 SSU rRNA 基因序列與
Enterocytozoon hepatopenaei (EHP)的 SSU rRNA 基因序列僅具有 71%的
同一性,所以這兩者親緣性差距較遠。Tang 等人(2015),將 Enterocytozoon
hepatopenaei (EHP) 18S rRNA 的基因序列與其他微孢子蟲 Nucleospora salmonis、Nucleospora cyclopteri 以及 Enterocytozoon bieneusi 的 18S rRNA
的基因序列具有82-83%的同一性。本實驗利用 NCBI 資料庫,將 EHP 微
孢子蟲SSU rDNA 核酸基因序列與其他國家的 EHP 微孢子蟲核酸基因序
列做比對,比對結果在泰國、印度、越南、中國、美國與拉丁美洲所發現
的 EHP 微孢子蟲的比對中分別為 99%、99.87、99.87、99.22、98.87%、
99.77%,所以台灣所發現的 EHP 微孢子蟲與其他國家所發現的 EHP 微
孢子具有親緣關係,但在比對的結果中,西班牙所發現的EHP 微孢子蟲
與台灣發現的EHP 微孢子蟲親緣關係較遠。
本實驗利用 DIG 標示的探針,以點墨雜交(Dot blot hybridization)檢
測探針之靈敏度,結果在含量高於13 pg/μL 的病毒核酸重組質體均可被
偵測到;若直接以病蝦所萃取之核酸進行檢測,結果可專一性的檢測到
43
中利用DIG 標示的探針,進行原位雜交(In situ hybridization)檢測,在實
驗結果中發現受感染的南美白對蝦肝胰臟中有明顯的陽性反應,在顯微
鏡下觀察時,可以看到南美白對蝦肝胰臟中有EHP 微孢子在細胞內,但
由於感染情況沒有很嚴重,所以EHP 微孢子蟲的分布沒有很多。在印度
Rajendran 等人(2016)以原位雜交(In situ hybridization)的方式觀察受 EHP 微孢子蟲感染的南美白對蝦(Penaeus vannamei),受感染的南美白對蝦 (Penaeus vannamei)在肝胰臟中,呈現正反應,在顯微鏡下觀察可以清楚
看見 EHP 微孢子蟲在細胞內。Tourtip 等人(2009)首次發現泰國斑節蝦
(Penaeus monodon) 感 染 EHP 微 孢 子 蟲 , 使 用 了 原 位 雜 交 (In situ hybridization)的方法檢測到 EHP 微孢子蟲,利用顯微鏡觀察到 EHP 微孢 子蟲在細胞內的狀態。 在上述的各項實驗結果,對於EHP 微孢子的發生在蝦類養殖中一年 四季都很有可能爆發,先前報告指示EHP 微孢子蟲雖然不會造成南美白 對蝦的死亡,但會使南美白對蝦生長遲緩,而且可以由水平傳播的方式 相互感染,所以 EHP 微孢子蟲具有很高的傳染力。在感染 EHP 微孢子蟲 的南美白對蝦,外觀上是不容易發現的,所以在本實驗中利用病理組織 切片觀察,PCR 檢測(以 EHP 專一性引子進行靈敏度檢測)、點墨雜交反 應(Dot blot hybridization)、原位雜交(In situ hybridization)等分子生物學的
44
EHP,但實驗中所使用 PCR 及 Nested PCR 的方式檢測 EHP 之靈敏度的
效果是較好的,因在操作上可以降低汙染風險,再者可以利用Nested PCR 來更進一步的檢測出南美白對蝦感染 EHP 之靈敏度。 EHP 微孢子蟲是近幾年發生的寄生蟲疾病,對於 EHP 微孢子蟲主要 的傳染源以及傳染因子尚未了解,所以根據報導所示,南美白對蝦在SPF 無特定病源的後期幼蟲中未檢測到EHP 微孢子蟲,但放養至養殖水池後 感染 EHP 微孢子蟲,是表明了 EHP 微孢子蟲主要是由養殖水池傳播而
來的 (Tangprasittipap et. al.,2013)。除了養殖水池的會感染 EHP 微孢子蟲
的可能性,另外在飼料上也是有可能存在 EHP 微孢子蟲,Tang 等人(2015) 將額外添加到蝦類飼料中的多毛類、魷魚以及豐年蝦,檢測是否是 EHP 微孢子蟲的攜帶者,使用PCR 做檢測,檢測結果在多毛類以及魷魚中使 用的 EHP 專一性引子沒有產生交叉反應,而在豐年蝦中檢測到 EHP 微 孢子蟲,並且使用來自越南EHP 微孢子 1.1 kb 基因片段做基因序列的檢 測,結果在豐年蝦中檢測到的 EHP 微孢子蟲與來自越南的 EHP 微孢子 蟲基因序列具有99.9 % 的同一性,這表示在蝦類飼料的額外添加中,需 要建立有效的控制與監測,來降低攜帶EHP 微孢子蟲的生物進入蝦類養 殖場,進而造成蝦類養殖感染的風險。
45
第五章 結論
本實驗中在上述所建立之檢測方法外,在未來的研究方向,將需要 朝著其他更靈敏及快速的檢測技術的研究,檢測技術固然重要,但更重
要的是需要對EHP 微孢子蟲的傳播、傳染 EHP 微孢子的特徵以及 EHP
微孢子感染其他甲殼類作為感染宿主的潛在作用進行研究。但預防重於
治療,所以需建立 EHP 微孢子蟲的疫苗研發以降低藥物的濫用,需建
立如何改善蝦類養殖環境來降低外來攜帶者生物感染養殖蝦類,以提供 更有價值的檢測方法來提升台灣蝦類養殖產業的技術。
46
第六章 參考文獻
丁雲源 (2005) 白蝦養殖發展史。水產試驗所特刊第 6 號:3-4。 台灣南區魚病中心, 中區於病中心, 台南市家畜疾病防治所, 屏東縣家畜 疾病防治所, 高雄縣家畜疾病防治所, 宜蘭縣家畜疾病防治所及台東 縣家畜疾病防治所(1994)台灣草蝦大量死亡病例之病因學,疫學,病理 組織學及電子顯微鏡觀察之研究. 83 年動物衛生報導, 行政院農委會, 台灣省政府農林廳編,雲林縣家畜疾病防治所出版, 南投, 37-49。 李定安 (2005) 對蝦類的分類學研究。水產試驗所特刊第 6 號:25-32。 陳弘成 (2001) 白蝦養殖要點。養殖漁業經濟管理手冊 技術篇。行政院 農委會漁業署編制。 黃世鈴、陳秀男 (2006) 鰻魚寄生蟲病。水產試驗所特刊第 8 號:13-50。 董明澄 (2005) 白蝦疾病診斷與防治。水產試驗所特刊第 6 號:101-105。 楊明樺、鄭金華、陳紫媖 (2016) 海水蝦的微孢子蟲感染症及其防治。水 試專訊:42-44。47
Anantasomboon, G., Sriurairatana, S., Flegel, T.W. and Withyachumnarnkul, B. (2006) Unique lesions and viral-like particles found in growth retarded black tiger shrimp Penaeus monodon from East Africa. Aquaculture 253, 197-203.
Amogan, H.P., Martinez, J.P., Ciuffette, L.M., Field, K.G and Reno, P.W. (2006) Karyotype and genome size of Nadelspora canceri determined by pulsed field gel electrophoresis. Acta Protozool. 45, 249-254.
Anderson, I.G., Shariff, M and Nash, G. (1989) A hepatopancreatic microsporidian parasite in pond-reared tiger shrimp, Penaeus monodon, from Malaysia. J. Invertebr. Pathol. 53, 278-280.
Bell, T. A. and D. V. Ligtner. (1984) IHHNV virus: Infectivity and pathogenicity studies in Penaeus stylirostris and Penaeus vannamei.
Aquaculture. 38:185-194.
Bonami, J. R., M. Brehelin., J. Mari., B. Trumper and D. V. Lightner. (1990) Purification and characterization of IHHN virus of penaeid shrimps. J.
Gen. Virol. 71, 2657-2664.
Bhat, S.A., Bashir, I and Kamili, A.S. (2009) Microsporidiosis of silkworm, Bombyx mori L. (Lepidoptera: Bombycidae): a review. African J. Agric.
Res. 4, 1519-1523.
Becnel, J.J. and Andreadis, T.G. (2014) Microsporidia in insects. In: Weiss LM, Becnel JL,editors. Microsporidia: Pathogens of Opportunity. First Ed. West Sussex:Wiley Blackwell. 521-570.
Canning, E.U., Curry, A and Overstreet, R.M., (2002) Ultrastructure of Tuzetia
weidneri sp. n. (Microsporidian parasitea: Tuzetiidae) in skeletal muscle
of Litopenaeus setiferus and Farfantepenaeus aztecus (Crustacea: Decapoda) and new data on Perezia nelsoni (Microsporidian parasite: Pereziidae) in L. setiferus. Acta Protozool. 41, 63-77.
Chayaburakul, K., Nash, G., Pratanpipat, P., Sriurairatana, S and Withyachumnarnkul, B., (2004) Multiple pathogens found in growth-retarded black tiger shrimp Penaeus monodon cultivated in Thailand. Dis.
48
Aquat. Org. 60, 89-96.
Chaweepack, T., Muenthaisong, B., Chaweepack, S and Kamei, K., (2015) The potential of galangal (Alpinia galangal Linn.) extract against the pathogens that cause white feces syndrome and acute hepatopancreatic necrosis disease (AHPND) in Pacific white shrimp (Litopenaeus
vannamei). Int. J. Biol. 7, 8-17.
Flegel, T.W., (2012) Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 110, 166-173.
Flegel, T.W., Nelson, L., Thamavit, V., Kongtim, S and Pasharawipas, T., (2004) Presence of multiple viruses in non-diseased, cultivated shrimp at harvest.
Aquaculture 240, 55-68.
Ha, N.T.H., Ha, D.T., Thuy, N.T and Lien, V.T.K., (2010) Enterocytozoon
hepatopenaei has been detected parasitizing tiger shrimp (Penaeus monodon) cultured in Vietnam and showing white feces syndrome. (In
Vietnamese with English abstract). Agric. Rural Dev. Sci. Technol. (translation from Vietnamese) 12, 45-50.
Hayman, J.R., Southern, T.R and Nash, T.E. (2005) Role of sulfated glycans in adherence of the microsporidian Encephalitozoon intestinalis to host cells in vitro. Infect Immun.73:841-848.
Hudson, D.A., Hudson, N.B and Pyecroft, S.B., (2001) Mortalities of Penaeus japonicas prawns associated with microsporidean infection. Aust. Vet. J. 79, 504-505.
Vávra, J and Lukeš. (2013) Microsporidia and 'the art of living together'. Adv.
Parasitol. 82, 253-319.
Kurita, J., Nakajima, K., Hirono, I and Aoki, T. (1998) Polymerase chain reaction (PCR) amplification of DNA of red sea bream iridovirus (RSIV).
Fish Pathol. 33, 17-23.
Landgdon, J.S. (1991) Microsporidiosis due to a pleistophorid in arron, Cherax tenuimanus (Smith), (Decapoda: Parastacidae). J. Fish Dis. 14, 33-44.
49
Lightner, D. V. (1996) The penaeid shrimp viruses IHHNV and TSV: epizootiology, production impacts and role of international trade in their distribution in the Americas. Rev. Sci. Tech. Off. Int. Epiz. 15: 579-601. Larsson, J.I.R., Ebert, D., Vävra, J and Voronin, V.N. (1996) Redescription of
Pleistophora intestinalis Chatton, 1907, a microspridian parasite of Daphnia magna and Daphnia pulex, with establishment of a new genus Glugoides (Microspora, Glugeidae). Eur. J. Protistol. 32, 251-261.
Larsson, J.I.R., Ebert, D., Vävra, J., (1997) Ultrastructural study and description of Ordospora colligata gen. et sp. nov. (Microspora, Ordosporidae fam. nov.), a new microsporidian parasite of Daphnia magna (Crustacea, Cladocera). Eur. J. Protistol. 33, 432-443.
Larsson, J.I.R., Ebert, D., Mangin, K.L., Vávra, J., (1998) Ultrastructural study and description of Flabelliforma magnivora sp. n. (Microspora: Duboscqiidae), a microsporidian parasite of Daphnia magna (Crustacean: Cladocera: Daphnia magna (Crustacea: Cladocera: Daphniidae). Acta
Protozool. 37, 41-52.
Leaño, E and Mohan, C., (2012) Disease Advisory: “Early Mortality Syndrome (EMS)/Acute Hepatopancreatic Necrosis Syndrome (AHPNS): An Emerging Threat in the Asian Shrimp Industry”. NACA, Bangkok, Thailand.
Leobert, D., Cabillon, N.A.R., Catedral, D.D., Amar, E.C., Usero, R.C., Monotilla, W.D., Calpe, A.T., Fernandez, D.D and Saloma, C.P., (2015) Acute hepatopancreatic necrosis disease (AHPND) outbreaks in Penaeus
vannamei and P. monodon cultured in the Philippines. Dis. Aquat. Org.
116, 251-254.
Lom, J., Nilson, F and Dykova, I., (2001) Dimorphic life cycle and taxonomic affinities, as indicated by ultrastructural and molecular study. Parasitol.
Res. 87, 860-872.
Lobo, M.L., Xiao, L., Cama, V., Magalhaes, N., Antunes, F., Matos, O. (2006) Identification of potentially human-pathogenic Enterocytozoon bieneusi genotypes in various birds. Appl. Environ. Microbiol. 72, 7380-7382.
50
Luis Fernando Aranguren, Jee Eun Han and Kathy F.J. Tang. (2017)
Enterocytozoon hepatopenaei (EHP) is a risk factor for acute
hepatopancreatic necrosis disease (AHPND) and septic hepatopancreatic necrosis (SHPN) in the Pacific white shrimp Penaeus vannamei.
Aquaculture 471, 37-42.
Kumar., Navaneeth Krishnan., Joseph Sahaya Rajan., Makesh, M., Jithendran, K., Alavandi, S. V and Vijayan, K. (2018) Visual loop-mediated isothermal amplification (LAMP) for the rapid diagnosis of
Enterocytozoon hepatopenaei (EHP) infection. Parasitol. Res. 117,
1485-1493
Kumar, S., Stecher, G., and Tamura, K. (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870-1874.
Moodie, E.G., Le Jambre, L.F and Katz, M.E., (2003) Thelohania
montirivulorum sp. nov. (Microspora: Thelohaniidae), a parasite of the
Australian freshwater crayfish, Cherax destructor (Decapoda: Parastacidae): fine ultrastructure, molecular characteristics and phylogenetic relationships. Parasitol. Res. 91, 215-228.
Mingxiao N., Panpan W., Hui S., Xihe W., Mingjian J., Xiangqian L., Hao S., Yi Q., Ge J., Wei G., Wen W., Li W. and Qingguo M. (2019) Proteomic and metabolomic responses in hepatopancreas of whiteleg shrimp
Litopenaeus vannamei infected by microsporidian Enterocytozoon hepatopenaei. Fish Shellfish Immun. 87, 534-545.
Mastan, S.A., (2015) Incidences of white feces syndrome (WFS) in farm-reared shrimp Litopenaeus vannamei, Andhra Pradesh Indo Am. J. Pharm.
51
Nägeli KW. (1857) Über die neue krankheit der seidenraupe und verwandte organismen. Bot Z. 15:760-761.
Notomi T, Okayama H and Masubuchi H et al. (2000) Loop-mediated isothermal amplification of DNA. Nucleic Acids Res.28, E63.
Newman, S.G., (2015) Microsporidian impacts shrimp production—industry efforts address control, not eradication. Glob. Aquac. Advocate 16-17. Oshima, S., Hata, J.I., Segawa, C., Hirasawa, N and Yamashita, S. (1996)
Amethod for direct DNA amplification of uncharacterized DNA virus and for development of a viral polymerase chain reaction assay application to the red sea bream iridovirus. Anal. Biochem. 242, 15-19.
Pasharawipas, T., Flegel, T.W., Chaiyaroj, S., Mongkolsuk, S and Sirisinha, S., (1994) Comparison of amplified RNA gene sequences from microsporidian parasites (Agmasoma or Thelohania) in Penaeus
merguiensis and P. monodon. Asian Fisheries Sci. 7, 169-178.
Pattana Jaroenlak, Dominic Wiredu Boakye, Rapeepun Vanichviriyakit, Bryony A. P. Williams, Kallaya Sritunyalucksana and Ornchuma Itsathitphaisarn. (2018) Identification, characterization and heparin binding capacity of a spore-wall, virulence protein from the shrimp microsporidian, Enterocytozoon hepatopenaei (EHP). Parasites &
Vectors. 11:177.
Patrick j. Keeling. (2004). Diversity and evolutionary history of plastids and their hosts. Am. J. Bot. 91, 1481-1493.
Refardt, D., Canning, E.U., Mathis, A., Cheney, S.A., Lafranchi-Tristem, N.J., Ebert, D., (2002) Small subunit ribosomal DNA phylogeny of microsporidia that infect Daphnia (Crustacea: Cladocera). Parasitol. 124, 381-389.
Rajendran, K., Shivam, S., Praveena, P.E., Rajan, J.J.S., Kumar, T.S., Avunje, S., Jagadeesan, V., Babu, S.P., Pande, A and Krishnan, A.N., (2016). Emergence of Enterocytozoon hepatopenaei (EHP) in farmed Penaeus
52
Rodriguez-Tovar., L.E., Speare, D.J and Markham R.J.F. (2011). Fish microsporidia: immune response, immunomodulation and vaccination.
Fish Shellfish Immunol. 30, 999-1006.
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989) Molecular cloning, a
laboratory manual. 2nd. Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, New York.
Salachan, P.V., Jaroenlak, P., Thitamadee, S., Itsathitphaisarn, O., and Sritunyalucksana, K. (2017) Laboratory cohabitation challenge model for shrimp hepatopancreatic microsporidiosis (HPM) caused by
Enterocytozoon hepatopenaei (EHP). BMC Vet. Res. 13, doi:
10.1186/s12917-016-0923-1.
Saitou, N. and Nei, M. (1987) The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol Evol. 4:406-425.
Sritunyalucksana, K., Apisawetakan, S., Boon-nat, A., Withyachumnarnkul, B., and Flegel, T.W., (2006). A new RNA virus found in black tiger shrimp Penaeus monodon from Thailand. Virus Res. 118, 31-38.
Stentiford, G.D and Bateman, K.S., (2007). Enterospora canceri sp., an intranuclear microsporidian parasite infection of hermit crab Eupagurus.
Dis. Aquat. Org.75, 73-78.
Stentiford, G.D., Bateman, K.S., Longshaw, M and Feist, S.W., (2007)
Enterospora canceri n. gen., n. sp., intranuclear within the
hepatopancreates of the European edible crab Cancer pagurus. Dis. Aquat.
Org. 75, 61-72.
Sprague, V., Becnel, J.J. and Hazard, E.I. (1992) Taxonomy of phylum Microspora. Crit. Rev. Microbiol. 18, 285-395.
Sritunyalucksana, K., Sanguanrut, P., Salachan, P.V., Thitamadee, S and Flegel, T.W., (2014). Urgent Appeal to Control Spread of the Shrimp Microsporidian Parasite Enterocytozoon hepatopenaei (EHP).
53
Sokolova, Y, Pelin, A, Hawke, J and Corradi, N. (2015) Morphology and phylogeny of Agmasoma penaei (microsporidia) from the type host,
Litopenaeussetiferus, and the type locality, Louisiana. USA Int. J. Parasitol. 45:1-16.
Tang, K.F.J., Pantoja, C.R., Redman, R.M., Han, J.E., Tran, L.H. and Lightner, D.V. (2015) Development of in situ hybridization and PCR assays for the detection of Enterocytozoon hepatopenaei (EHP), a microsporidian parasite infecting penaeid shrimp. J Invertebr Pathol. 130, 37-41.
Tangprasittipap, A., Srisala, J., Chouwdee, S., Somboon, M., Chuchird, N., Limsuwan, C., Srisuvan, T., Flegel, T.W and Sritunyalucksana, K. (2013) The microsporidian Enterocytozoon hepatopenaei is not the cause of white feces syndrome in whiteleg shrimp Penaeus (Litopenaeus)
vannamei. BMC Vet. Res. 9139-9,148.
Tang, K.F., Han, J.E., Aranguren, L.F., White-Noble, B., Schmidt, M.M., Piamsomboon, P., Risdiana, E and Hanggono, B. (2016) Dense populations of the microsporidian Enterocytozoon hepatopenaei (EHP) in feces of Penaeus vannamei exhibiting white feces syndrome and pathways of their transmission to healthy shrimp. J. Invertebr. Pathol. 140, 1-7.
Tamura, K., Nei, M., and Kumar, S. (2004) Prospects for inferring very large phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Sciences (USA) 101:11030-11035.
Tourtip, S., Wongtripop, S., Stentiford, G.D., Bateman, K.S., Sriurairatana, S., Chavadej, J., Sritunyalucksana, K and Withyachumnarnkul, B., (2009)
Enterocytozoon hepatopenaei sp. nov. (Microsporida:
Enterocytozoonidae), a parasite of the black tiger shrimp Penaeus monodon (Decapoda: Penaeidae): Fine structure and phylogenetic
relationships. J. Invertebr. Pathol. 102, 21-29.
Tomita, N., Mori, Y., Kanda, H and Notomi, T. (2008) Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products. Nat Protoc 3, 877-882.