行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
Genome wide analysis of expression of the fimbrial operons in
Klebsiella pneumoniae
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 94-3112-B-009-004-
執行期間: 九十四 年 五 月 一 日 至 九十七 年 四 月 三十 日
計畫主持人:彭慧玲
共同主持人:
計畫參與人員:吳健誠,黃盈蓉,廖朝陽,黃登魁,鐘純珊
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
■出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,■一年□二年後可公開查詢
執行單位: 交通大學生物科技學系
中 華 民 國 九十七 年 三 月 二十八 日
TABLE OF CONTENTS
I. Abstracts
Ia. Abstract in Chinese
page II-IIIIb. Abstract in English
page IV-VII. Background and Significance
page 1III. Material and Methods
page 3IV. Progress Report and Discussion
page 6V. Literature Cited
page 22I. Abstract
Ia. Abstract in Chinese
關鍵詞:克雷白氏肺炎桿菌,kpa,kpb,kpc,kpd,kpe,kpf 和 kpg 纖毛,FimA、 MrkA、MrkD、MrkF 多株抗體,第三型纖毛表現載體,MrkD、KpbD、KpfD 黏附蛋白,FimB、PhgS、PhgM、FimK、KpdR 調控蛋白。 在克雷白氏肺炎桿菌NTUH K-2044 基因體中,除了已知的第一型及 第三型纖毛外,我們另外發現了七套未被發表過的纖毛基因組,分別命名 為 kpa,kpb,kpc,kpd,kpe,kpf 和 kpg。為了了解這九套纖毛基因組在 黏附特異性及表現調控上的異同,這三年完成的工作如下: 1. 以 ClustalW 工具分別進行纖毛蛋白質的序列分析,可發現這九套纖毛之 間並沒有明顯的親緣關係。纖毛基因存在分佈率的研究結果顯示,不同來 源的105株克雷白氏肺炎桿菌分離株,大多都含有kpa、kpe、kpg、fim,以 及mrk纖毛基因組 (87% to 100%)。此外,我們發現kpb和kpc纖毛基因組的 分佈則與K1血清型有密切的相關性(P < 0.0001);在35株K1莢膜血清型分 離株中,有32株具有相同的纖毛基因組合。 然而,這九套纖毛基因組的 存在率與疾病種類並沒有明顯的關聯性。 2. 我們分別將這九套纖毛基因組起使子轉殖到LacZ通報系統,然而,除了第 一、三型纖毛外,其他七套基因組起使子因活性太低而無法進一步分析; 我們也將七套纖毛基因組自克雷白氏肺炎桿菌 NTUH K-2044中選殖出 來,並且以不會生成纖毛的大腸桿菌為宿主,分別表現於大腸桿菌表面, 以SDS-PAGE 分析可以偵測到 kpb 和 kpc纖毛的表現,目前在準備製造 KpbA和KpcA抗體,未來將以免疫螢光顯微鏡觀察來確認二者的表現;另 外,我們還把八套纖毛的黏附蛋白基因選殖置換第三型纖毛表現載體上的 mrkD黏附蛋白基因,以血球凝集測試發現KpbD和KpfD會凝集未經單寧酸 處理的兔子和天竺鼠紅血球,以KpcD 或FimH取代MrkD黏附蛋白時會減 少第三型纖毛的單元聚合;最後,我們還製備了MrkA、MrkD、MrkF 及 FimA 多株抗體,可分別第一、三型纖毛的表現。 3. 經由穿透式電子顯微鏡、西方墨點法以及線毛活性的測定,證實黏附蛋白 MrkD基因序列的變化影響了第三型線毛的組裝和活性;藉雷射鑷夾量,
我們發現MrkDv3對膠原蛋白的黏附力最大;最後,我們也證明MrkF為第 三型纖毛的結構蛋白質,具有MrkF 組成會減少第三型纖毛的單元聚合而 使第三型纖毛較短而糾結,可以提高細菌生物膜的形成。 4. 我們證明第一型及第三型纖毛的表現有互相拮抗的效應,在CG43或 NTUHK-2044菌株中可以偵測到第三型纖毛的表現,而相對的只有在MrkA 基因被剔除時,才能偵測到第一型纖毛的表現;而在大量表現FimB的菌株 會活化第一型纖毛的表現,卻無法偵測到第三型纖毛的表現。 5. 啟動子活性測試顯示-phgS 和phgM的基因缺損會降低Pkpg的活性;而剔除 活化夾膜多醣合成基因表現的調控蛋白RcsB基因,可以提高PfimA 活性而 不影響 PmrkA的活性。另外,在大量表現FimB的菌株中發現PmrkA活性降低。 6. 大量表現KpdR可降低生物膜的生成,同時增加TEM-116 β-lactamase 酵素 的生成而提高細菌對β-lactam抗生素的抗性;而大量表現FimK能降低夾膜 多醣的合成,其作用機制仍待證明;大量表現KpbR和KpfR卻抑制細菌的 生長,我們將改可以甘露醣誘導表現載體進一步分析這些轉錄調控蛋白的 功能。
I. Abstract
Ib. Abstract in English
Keywords:Klebsiella pneumoniae, kpa, kpb, kpc, kpd, kpe, kpf, and kpg fimbriae,
FimA, MrkA, MrkD, MrkF polycolona antibodies, type 3 fimbriae expression plasmid, MrkD, KpbD and KpfD ahesins, FimB, PhgS, PhgM, FimK and KpdR regulatory proteins.
In the genome of Klebsiella pneumoniae NTUH K-2044, there are nine putative fimbrial operons encoding respectively type 1 and type 3 fimbriae, and seven novel fimbriae kpa, kpb, kpc, kpd, kpe, kpf, and kpg. The accomplished experiments in analysis of the expression of these fimbriae are as following:
(1) Multiple sequence alignment of the fimbrial genes by ClustalW revealed that the four major components of the nine fimbrial operons are distinct to each other. The prevalence analysis of the nine fimbrial gene clusters among 105 K.
pneumoniae clinical isolates from various types of disease indicated that kpa, kpe, kpg, fim, and mrk genes were contained in most of the isolates (87% to 100%). In
addition,kpb and kpc genes were present in a higher frequency (P < 0.0001) in K1
isolates. In addition, 32 of 35 K1 isolates appeared to possess identical repertoire of fimbrial operons. However, no obvious correlation between the fimbrial types and diseases could be identified.
(2) Each of the putative promoters has been cloned respectively into a LacZ reporter vector. However, except the promoters of type 1 and 3 fimbrial genes, the promoter activities of the other seven were too low to be detected. We have also tried to isolate each of the fimbrial operons by PCR-based cloning from K.
pneumoniae NTUH K-2044 and the fimbriae displayed on the surface of an
afimbriated E. coli. Heterologous expression of the recombinant type 3 fimbriae, and kpb and kpc fimbriae could be demonstrated by SDS-PAGE analysis. Polyclonal antibodies against KpbA and KpbC are currently prepared for immunofluorescence microscopy analysis. Furthermore, the filament pattern analysis revealed that the replacement of mrkD on type 3 fimbriae displaying plasmid with either kpcD or
could agglutinate both erythrocytes of guinea pig and rabbit. Finally, polyclonal antibodies against MrkA, MrkD, MrkF and FimA have been obtained and used for the analysis of the expression of type 1 and type 3 fimbriae.
(3) As demonstrated using TEM and western blotting analysis, and the assessments for fimbrial activity, allelic variations of mrkD adhesin were shown to affect fimbriation and activity of type 3 fimbriae. We have also employed optic tweezers to measure the adhesive force to collagen and the result revealed that the fimbriae expressing MrkDv3 had the highest binding force to collagen. Finally, MrkF was shown to be a component of type 3 fimbriae and the incorporation of MrkF conferred the recombinant type 3 fimbriae shorter and meshwork like morphology which led to a higher level of biofilm formation activity.
(4) We have demonstrated a counter-expression between type 1 and 3 fimbriae. The expression of type 3 fimbriae could be readily detected in either K. pneumoniae CG43 or NTUH-K2044. Expression of type 1 could only be detected in mrkA deletion mutant. Overexpression of FimB was shown to switch on the expression of type 1 fimbriae but block the type 3 fimbrial expression.
(5) Promoter activity measurement using LacZ as reporter indicated that the deletion of either phgS or phgM reduced the activity of Pkpg. The deletion of rcsB, which encoding the positive regulator for CPS synthesis, increased the activity of PfimA but had no apparent activity of PmrkA. Whereas overexpression of FimB was shown to reduce PmrkA activity.
(6) Over-expression of KpdR reduced the biofilm formation and increased the expression of β-lactamase TEM-116 in both K. pneumoniae NTUH-K 2044 and CG43S3. The FimK overexpression was shown to reduce the capsular polysaccharide biosynthesis. However, overexpression of either KpbR or KpfR was shown to inhibit the bacterial growth. A pBAD-based mannose inducible expression vector will be used to further investigate functional roles of the regulatory proteins.
II. Background and Significance
The biosynthesis and binding properties of type 1 fimbriae have been well studied in
Escherichia coli (15, 16, 17, 18). Although with less information, type 1 fimbriae
have also been isolated and characterized in K. pneumoniae (4, 8, 9). Type 3 fimbrial adhesin, referred to as mannose-resistant Klebsiella-like (MR/K) hemagglutinin, has been shown to be produced by a few of K. pneumoniae strains (1, 9, 10). In addition to the two fimbrial adhesins, three more including the fimbrial antigen KPF-28, the afimbrial adhesin CF29K, and an afimbrial adhesin composed of capsule-like extracellular material have recently been reported in Klebsiella isolates (2, 3, 6).
Adhesin specificity plays a major role in determining the host range and tissue tropism in bacterial infection (21). Depending on the expression of type 1 or P fimbriae, UPEC (uropathogenic E. coli) vary significantly in their abilities to colonize and persist within the bladder or kidneys (27). PapB, a regulator for phase variation of P fimbriae, and its functional homologue, SfaB of S fimbriae, both were shown to be able to increase the frequency of ON-to-OFF phase for type 1 fimbriae (12, 26). This indicated the presence of a regulatory network in controlling differential expression of type 1-, P-, and S-fimbriae in UPEC. In E. coli, type 1 fimbriae has been demonstrated to be essential for the bacteria in establishment and persistence of urinary tract infections (15, 27). Most likely, the fimbriae are also required for K. pneumoniae to colonize urinary tract. In addition to uroepithelial cells, the adherence to respiratory epithelia, intestinal and endothelial cells of the K.
pneumoniae strains expressing type 3 fimbriae have been demonstrated (20, 22).
However, the specificity and regulation of fimbrial adherence in K. pneumoniae are largely unknown.
Prevalence analysis of Salmonella enterica sp. Typhi isolates revealed a unique repertoire of fimbrial gene clusters possibly due to specific selective pressures (23, 24). Expression analysis using flow cytometry of eleven S. Typhimurium fimbrial operons indicated further that in vivo growth conditions drastically alter the
expression of repertoire of fimbrial antigens (14). Thus, differential expression of the nine fimbriae for K. pneumoniae to adhere to different cells or abiotic surfaces, or for the bacterial self-aggregation or biofilm formation could be foreseen. Using bioinformatic analysis, nine sets of fimbriae encoding operons of the chaperone-usher assembly pathway which includes type 1-, type 3- and seven novel-type fimbriae, namely kpa, kpb, kpc, kpd, kpe, kpf, and kpg in the genome of
K. pneumoniae NTUH K-2044 (http://genome.nhri.org.tw/kp/index.php), a K1
isolate of pyogenic liver abscess (5), were identified. The specific aims proposed to be accomplished included:
(1) Analysis of the fimbrial gene clusters in K. pneumoniae using the tools of bioinformatics
(2) Investigation of the prevalence of the fimbrial gene clusters in K.
pneumoniae clinical isolates
(3) Determination of the regulatory control of expression of the fimbrial operons
(4) Characterization of the adherence specificities displayed by each of the fimbrial adhesins
III. Material and Methods
Bacterial strains and growth condition. K. pneumoniae NTUH-K2044, of K1
serotype, was provided by Dr. Jin-Town Wang from National Taiwan University Hospital (5). Clinical isolates of K. pneumoniae used in this study were recovered from different tissue specimens of patients with a variety of infections at the Veteran General Hospital, Taipei, from 1991 to 1998. The strains were identified and serotype determined. The bacteria were grown in Luria-Bertani (LB) broth or agar at 37°C and stored at -80°C before use.
PCR amplification for prevalence analysis. The PCR mixture contained 20 mM
Tris-HCl (pH 8.4), 50 mM KCl, 1.5 mM MgCl2, 200 μM (each) deoxynucleoside
triphosphates (dATP, dCTP, dGTP, and dTTP), and 1 U of recombinant Taq DNA polymerase (Violet Bioscience Inc.) along with each of the K. pneumoniae genomic DNA and the specific primers. The primer pairs designed for the respective adhesin and pilin encoding genes of each fimbrial operons are shown in Table 1. The amplification cycle consisted of an initial 1 min hold at 95°C followed by 35 cycles of 95°C for 1 min, 50°C for 1 min, and 72°C for 1 min, and finally an elongation step for 10 min at 72°C. The amplified PCR product was then analyzed by electrophoresis on a 1% agarose gel.
Hemagglutination activity. The agglutination assay was carried out as described1
with some modifications. In brief,overnight grown bacteria were suspended in PBS to approximately 109 CFU/ml and the collected erythrocytes of guinea pig and rabbit washed twice with PBS. A series of two-fold dilution of the bacterial suspension were mixed with an equal volume of 3% (vol/vol) erythrocytes in PBS. The mixture was incubated at room temperature for 30 min to allow erythrocytes settle to the 96-well
V-bottomed microtiter plates.
Antiserum preparation. The coding regions corresponding to FimA and MrkA, the
major pilin subunit of type 1 fimbriae and type 3 fimbriae, were amplified by PCR and cloned into expression vector pET30, respectively. The expression plasmid was then transformed into E. coli Nova-Blue (DE3) and overexpression of the recombinant pilin was induced by addition of 0.5 mM IPTG (isopropyl-β-D-thiogalactopyranoside). The MrkA and FimA proteins fusion with His-tag were then purified using a nickel charged resin (Novagen, Madison, WI). In order to raise antibody, five-week-old female BALB/c mice, purchased from the animal center of National Taiwan University, were immunized intraperitoneally with 5 μg of the purified MrkA or FimA. Ten days later, the mice were immunized again with 5 μg of the MrkA or FimA protein. Finally, the antisera were obtained by intracardic puncture.
Western blot analysis of the expression of type 1 and type 3 fimbriae. The
clinical isolates of K. pneumoniae were grown overnight in either LB, for optimal expression of type 1 fimbriae, or GCAA (minimal medium supplemented with 1% glycerol and 0.3% casamino acids) broth for optimal expression of type 3 fimbriae. Total cellular proteins of the bacteria were resolved by SDS-PAGE and electrophoretically transferred from the gels onto PVDF membranes (ImmobilonTM-P, Millipore). The membranes were then blocked with 5% skimmilk at room temperature for 1 h, and washed 3 times with 1 X phosphate-buffered saline (PBS). Subsequently, the membranes were incubated with diluted anti-FimA or anti-MrkA serum at room temperature for 1 h. After 3 washes with 1 X PBS, a 3000-fold diluted alkaline phosphatase-conjugated anti-mouse immunoglobulin G
was added and the incubation continued for one more hour. The blot was again washed and the bound antibodies were detected by using the chromogenic reagents BCIP (5-bromo-4-chloro-3-indolyl phosphate) and NBT (Nitro blue tetrazolium).
Type 3 filament analysis Total proteins were isolated from the recombinant bacteria
E. coli, which has been cultured for 20 h in GCAA medium, and resolved by 7.5%
SDS-PAGE. The filament polymer was then assessed using Coomassie blue staining or western blotting analysis against anti-MrkA polyclonal antibody.
Transmission electron microscopy (TEM). Twenty microliters of bacterial
suspension (108 cfu/ml) were added to collodion-coated copper grids (300 mesh) and negatively stained by 2% phosphotungstic acid, pH 7.2. The grids were then examined under a JEOL JEM 2000EXII transmission electron microscope at an operating voltage of 100 kV.
Promoter activity measurement. β-galactosidase activity assay was carried out
essentially as described by Miller. The data presented were derived from a single experiment which was representativeof at least three independent experiments. Every sample was assayed in triplicate, and the average activity and standard deviation were presented.
IV. Progress Report and Discussion
Specific Aim 1- Analysis of the fimbrial gene clusters in K. pneumoniae using the tools of bioinformatics. Using HMMER to search for the Pfam fimbrial family
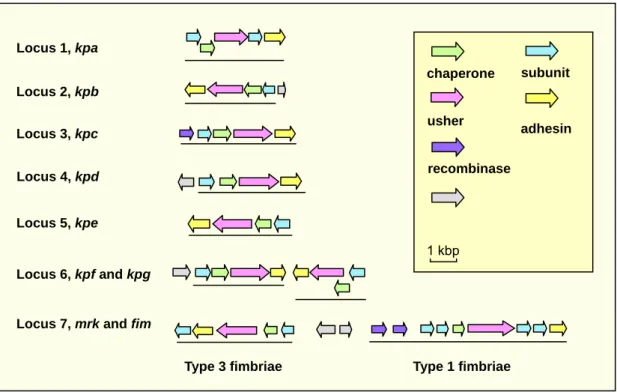
and BLAST analysis of the K. pneumoniae NTUH K-2044 genome, nine distinct putative fimbrial operons including type 1 and type 3, and seven novel types of fimbriae, namely kpa, kpb, kpc, kpd, kpe, kpf, and kpg were identified (Fig. 1).
FIG. 1. Fimbrial gene clusters in K. pneumoniae NTUH-K2044
(http://genome.nhri.org.tw/kp/index.php). Each of the putative fimbrial operons is underlined.
The BLAST analysis also revealed that, except the kpbABCD and kpeABCD operons, homologs could be identified in other bacteria for kpa, kpc, kpd, kpg, and
kpf gene clusters. The kpaABCDE operon, is likely an orthologue of Salmonella
Typhimurium LT2 sthABCDE operon, which has been reported to be required for intestinal persistence in mice. In kpa operon,both kpaA and kpaD genes appeared to
Locus 1, kpa Locus 2, kpb Locus 3, kpc Locus 4, kpd Locus 5, kpe Locus 6, kpf and kpg Locus 7, mrk and fim
chaperone usher subunit adhesin 1 kbp recombinase
be capable of encoding a fimbrial subunit suggesting that they are respectively the major and minor subunits of the fimbrial rod. The kpcABCD are homologous to the putative fimbrial genes, plu2159 to plu2156, of Photorhabdus luminescens subsp. laumondii TTO1. The kpdABCDencodes a homologous operon of Reut02000958 to Reut02000961 in Ralstonia metallidurans CH34 (unfinished sequence, GenBank: AAAI00000000). The physically linked kpfABCD operon and kpgABCD genes appeared to be respectively an orthologue of Edwardsiella tarda KG8401 etfABCD operon, and an orthologue of the fimbrial operon, plu2159 to plu2156, in P.
luminescens subsp. laumondii TTO1.
Upstream of the kpbABCD operon, a divergently transcribed gene encoding a transcriptional regulator with a DNA-binding domain was identified and designated
kpbR (Fig. 1). A DNA recombinase encoding gene, namely kpcI, located upstream of kpcABCD, was found to be an orthologue of several fimbrial invertases including
MrpI, MrxI (AAM91928) and MrfI. As shown in Fig. 2, an 11-bp inverted repeat was identified in the putative promoter region of kpc operon, impling a phase-variation control by DNA inversion for the operonexpression. Moreover, TCS (two component system) response regulator encoding genes, namely kpdR and kpfR, were found to be located upstream of kpdABCD and kpfABCD operons (Fig. 1).
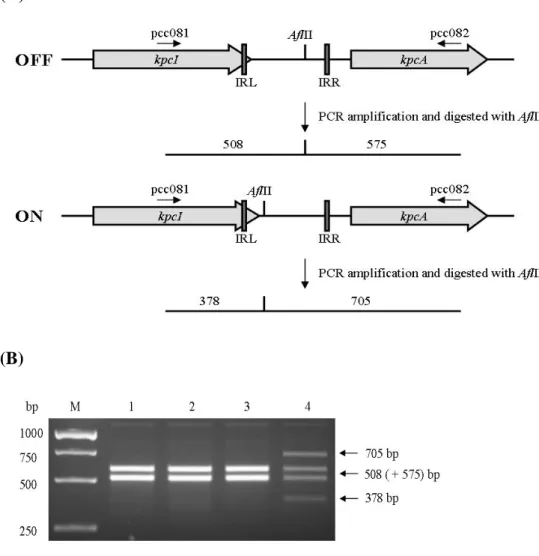
While introducing into the bacteria NTUH-K2044 with a kpcI containing plasmid pKC-19, inversion of the noncoding DNA upstream of KpcA start codon was observed (Fig. 3) which implying the presence of a phase variation control for the expression of kpc fimbriae.
(A)
(B)
FIG. 3. (A) Schematic presentation of the predicted invertible DNA in-between
kpcI and kpcA. (B) Effect of KpcI expression on the DNA inversion. The isolated
PCR products were digested with AflII and resolved on 1.2% agarose gel. Lanes 1, NTUH-K2044[pAW03] in 0.4% glucose; 2, NTUH-K2044[pAW03] in 0.4% mannose; 3, NTUH-K2044[pKC-19] in 0.4% glucose; NTUH-K2044[pKC-19] in 0.4% mannose.
(B) No. of isolates (n = 123) presenting specific fimbriae-related DNA sequences (%)
(A) Distribution of fimbrial genes in combination with each other among different K serotype bacteria
No. with genes (%) Presence of gene 122 (99) + + – + + + + + + + + + + + + + + + + kpa 6 (7) 32 (91) + + + + + + + + 123 (100) 106 (86) 120 (98) 88 (72) 116 (94) 103 (84) 40 (33) 64 (52) 2 (2) 0 + – + – + + – – 1 (1) 0 + + + – + + + + 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 2 (2) 4 (5) 6 (7) 7 (8) 10 (12) 9 (11) 25 (29) Other serotypes 0 + – + – + + – – 5 0 + + + – + + – + 0 + – + – – – – – 1 (3) + + + – + + – – 1 (3) + + + + + + – + 0 + + + + + – – – 1 1 (3) + + + + + + – – 0 + – – – + + – – 0 + – – + + + – – 0 + – + – – – – + 0 + – + – + + + – 0 + – – – + + – + 0 + – + + + – – + 0 + – + – + + – + 0 + + + + + – – + 1 0 + + + – + – – – K2 serotype K1 serotype mrk fim kpg kpf kpe kpd kpc kpb
(B) No. of isolates (n = 123) presenting specific fimbriae-related DNA sequences (%)
(A) Distribution of fimbrial genes in combination with each other among different K serotype bacteria
No. with genes (%) Presence of gene 122 (99) + + – + + + + + + + + + + + + + + + + kpa 6 (7) 32 (91) + + + + + + + + 123 (100) 106 (86) 120 (98) 88 (72) 116 (94) 103 (84) 40 (33) 64 (52) 2 (2) 0 + – + – + + – – 1 (1) 0 + + + – + + + + 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 1 (1) 2 (2) 4 (5) 6 (7) 7 (8) 10 (12) 9 (11) 25 (29) Other serotypes 0 + – + – + + – – 5 0 + + + – + + – + 0 + – + – – – – – 1 (3) + + + – + + – – 1 (3) + + + + + + – + 0 + + + + + – – – 1 1 (3) + + + + + + – – 0 + – – – + + – – 0 + – – + + + – – 0 + – + – – – – + 0 + – + – + + + – 0 + – – – + + – + 0 + – + + + – – + 0 + – + – + + – + 0 + + + + + – – + 1 0 + + + – + – – – K2 serotype K1 serotype mrk fim kpg kpf kpe kpd kpc kpb
Specific aim 2- Investigation of the prevalence of the fimbrial gene clusters in K. pneumoniae clinical isolates. Prevalence analysis via PCR detection of the
respective pilin and adhesin encoding genes in 105 K. pneumoniae clinical isolates from various types of diseases (collected by Dr. CP Fung from the Veteran General Hospital Taipei) revealed that kpa, kpe, kpg, fim, and mrk genes were contained in most of the isolates (87% to 100%). As shown in Table 1, the distribution of kpb, kpc,
kpd, and kpf genes were 50%, 32%, 83%, and 70%, respectively. In addition, 32 of
35 K1 isolates appeared to possess identical repertoire of fimbrial operons. No obvious correlation between the fimbrial types and diseases could be identified.
TABLE 1. Frequency and repertoire of fimbrial genes among K. pneumoniae
62 (50) 43 (3)
10 (2) 14 (2)
Type 3 fimbriae
% of isolates expressing fimbriae under the specific conditions (No.)
1 (8) 14 (1) 0 (0) 71 (10) Type 1 fimbriae Other serotypes (81) K2 isolates (7) K1 isolates, nonKLA (21) K1 isolates, KLA (14)
K. pneumoniaeisolates (No.)
Expression of fimbriae 62 (50) 43 (3) 10 (2) 14 (2) Type 3 fimbriae
% of isolates expressing fimbriae under the specific conditions (No.)
1 (8) 14 (1) 0 (0) 71 (10) Type 1 fimbriae Other serotypes (81) K2 isolates (7) K1 isolates, nonKLA (21) K1 isolates, KLA (14)
K. pneumoniaeisolates (No.)
Expression of fimbriae
Nevertheless, kpb and kpc gene clusters were found to be more prevalent (P < 0.0001) in the isolates of K1 serotype (Table 2). The results indicated that the specific combination and regulation of these fimbrial operons may play important roles in K. pneumoniae K1 pathogenesis.
TABLE 2. Prevalence and distribution of kpb (A) and kpc (B) genes.
*, P < 0.0001 31/88 (35)* 33/35 (94)* Total Other serotypes K1 serotype 4/9 Ascites 4/18 7/8 Blood 6/18 6/6 Wound 5/15 5/5 sputum 4/14 1/2 Urine 4/9 Bile 4/5 14/14 Liver abscess
No. of strains/total no. of strains isolated (%) K. pneumoniae isolates *, P < 0.0001 31/88 (35)* 33/35 (94)* Total Other serotypes K1 serotype 4/9 Ascites 4/18 7/8 Blood 6/18 6/6 Wound 5/15 5/5 sputum 4/14 1/2 Urine 4/9 Bile 4/5 14/14 Liver abscess
No. of strains/total no. of strains isolated (%) K. pneumoniae isolates *, P < 0.0001 8/88 (11)* 32/35 (91)* Total Other serotypes K1 serotype 1/9 Ascites 2/18 7/8 Blood 1/18 6/6 Wound 1/15 4/5 sputum 0/14 1/2 Urine 1/9 Bile 2/5 14/14 Liver abscess
No. of strains/total no. of strains isolated (%) K. pneumoniae isolates *, P < 0.0001 8/88 (11)* 32/35 (91)* Total Other serotypes K1 serotype 1/9 Ascites 2/18 7/8 Blood 1/18 6/6 Wound 1/15 4/5 sputum 0/14 1/2 Urine 1/9 Bile 2/5 14/14 Liver abscess
No. of strains/total no. of strains isolated (%)
K. pneumoniae
isolates
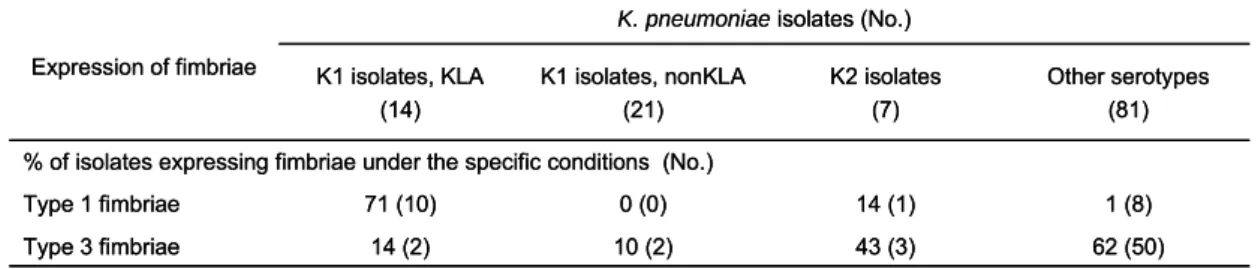
Interestingly, western blotting hybridization using anti-FimA and anti-MrkA antibodies, respectively, revealed that only 15.7% and 46.3% of the isolates expressed in vitro the major subunit of type 1 and type 3 fimbriae. Nevertheless, type 1 fimbriae were found to be prevalently expressed (71%) in liver abscess isolates of K1 serotype suggesting a not yet identified role of type 1 fimbrial expression for the infection of K1 isolates causing liver abscess (Table 3).
TABLE 3. In vitro expression of type 1 and type 3 fimbriae.
Specific aim 3- Determination of the regulatory control of expression of the fimbrial operons. In order to measure the expression of each of the fimbriae, the
putative promoter regions of each of the nine fimbrial gene clusters have been isolated from K. pneumoniae NTUH-K2044, and cloned into the reporter plasmid placZ15 containing a promoterless lacZ gene, which allow determine the signals that affect the fimbriae expression and the promoter activity of each of the fimbriae (Fig. 3). The promoter activities were measured in the presence of different cultural conditions such as changes of nutrient sources, growth temperature, osmolarity, and iron concentration. kpa kpb kpc kpd kpe kpf and kpg 1 kb kpa kpb kpc kpd kpe kpf and kpg 1 kb
FIG. 4. Schematic diagram of the putative promoters of kpa, kpb, kpc, kpd, kpe, kpf, and kpg operons
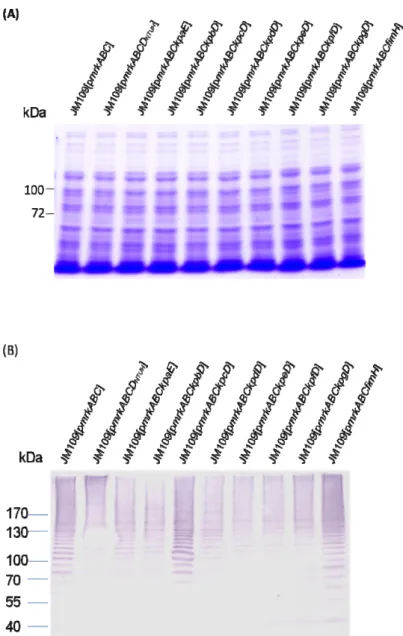
The heterologous expression constructs in Escherichia coli for each of the pilins have been generated. As shown in Fig. 4, expression of the major pilin of kpb and
FIG. 5. Heterologous expression of kpb- and kpc-encoded fimbriae. The plasmids
pKPB and pKPC respectively contain kpb and kpc genes and the expression was induced by 0.5 mM IPTG for 3 h . Protein marker (M); E. coli Novablue (DE3) containing pET30 (lanes 1 and 2), pKPB (lanes 3 and 4), or pKPC (lanes 5 and 6). The arrows respectively mark the expression of major subunit of kpb and kpc fimbriae.
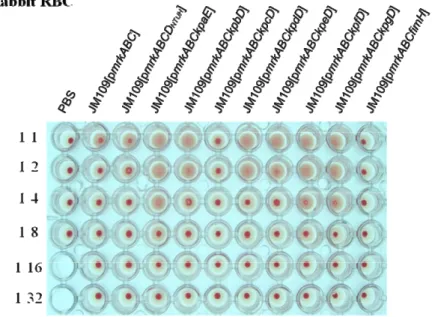
The filament pattern analysis revealed that the replacement of mrkD on type 3 fimbriae displaying plasmid with either kpcD or fimH reduced the polymer formation of the MrkA filament (Fig. 6A and B). In addition, the hemagglutination assay indicated that the displayed KpfD adhesin could bind to erythrocytes of guinea pig and rabbit (Fig. 7A and B). In analogy with the recent structural analysis of type 1 fimbriae, further investigation of the interacting domain of each of the adhesins with MrkA pilin could be predicted.
IPTG (mM) 0.5 N 0.5 pET30 14 18.4 28.5 37.6 66.2 116 87.4 pKPB pKPC N N 0.5 M 1 2 3 4 5 6
FIG. 6. Analysis of polymerization pattern of the recombinant type 3 fimbriae. The total proteins were isolated from the recombinant bacteria E. coli ,
which has been cultured for 20 h in GCAA medium, and resolved by 7.5% SDS-PAGE. The filament polymer was assessed using Coomassie blue stain (A) or western blotting analysis against anti-MrkA polyclonal antibody (B). Molecular mass markers are also indicated.
FIG. 7. Hemagglutination assay. Aliquots (109 CFU/ml) of the recombinant E.
coli, which has been cultured for 20 h in GCAA medium, were suspended in PBS
and then diluted by 2-fold gradient in PBS (from 1:1 to a 1:32). Fifty microliters of the bacterial suspension was then added to each well with an equal volume of 3% rabbit RBC(A) or guinea pig RBC(B), and the plate stood at room temperature till the appearance of agglutination.
Specific aim 4- Determination of the regulatory control of expression of the fimbrial operons.
Four novel mrkD alleles were identified and the growth of type 3 fimbriae is
MrkD adhesin dependent . Four mrkD variants were identified from 17 meningitis
isolates using PCR-RFLP analysis, and each of the PCR products was cloned, sequences determined, and named mrkDV1, mrkDV2, mrkDV3, and mrkDV4 (under the
GenBank accession number AY225462, AY225463, AY225464, and AY225465). The TEM analysis (Fig. 8) revealed different forms of fimbriae indicating the filament growth is adhesin dependent and effects of the allelic variation on the fimbrial activity had been demonstrated (13).
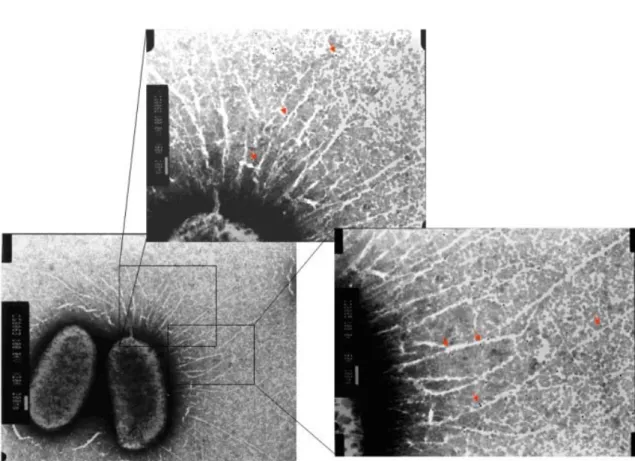
MrkF is a component of type 3 fimbriae. Fimbriation analysis of the E. coli
displaying with various recombinants of type 3 fimbriae including mrkABC,
mrkABCD, mrkABCF, mrkABCDF using transmission electronic microscopy (Fig. 9),
western blotting analysis, and co-immunoprecipitation, demonstrated that MrkF is a component of type 3 fimbriae.
FIG. 8. Transmission electron micrographs of the type 3 fimbriae displaying on E.
coli. One drop of bacterial suspension was placed on a carbon-coated copper grid,
and negatively stained with 2 % phosphotungstic acid. (A) E. coli JM109 [pGEM-T], (B) E. coli JM109 [pmrkABC], (C) E. coli JM109 [pmrkABCF], (D) E. coli JM109 [pmrkABCDV1F], (E) E. coli JM109[pmrkABCDV2F], (F) E. coli JM109
[pmrkABCDV3F],(G) E. coli JM109 [pmrkABCDV4F].
FIG. 9. Type 3 fimbriae labeled using anti-MrkF immuno-gold. The magnified
A cross-talk regulation is present for the expression of type 1 and 3 fimbriae. As
shown in Fig. 10A, the gene clusters respectively encoding type 1 and type 3 fimbriae are physically linked. Deletion of mrkA, which encoding the major pilin of type 3 fimbriae, increased the expression of type 1 fimbriae (Fig. 10B). As shown in Fig. 10C, The overexpression of fimB recombinase enhanced the expression of type 1 fimbriae but reduced the expression of type 3 fimbriae (Fig.10C).
(A)
(B)
FIG. 10. (A) Organization of the fim-mrk gene clusters of type 1 fimbriae and type 3
fimbriae. (B) and (C) Western blot analysis of the expression of FimA and MrkA proteins.
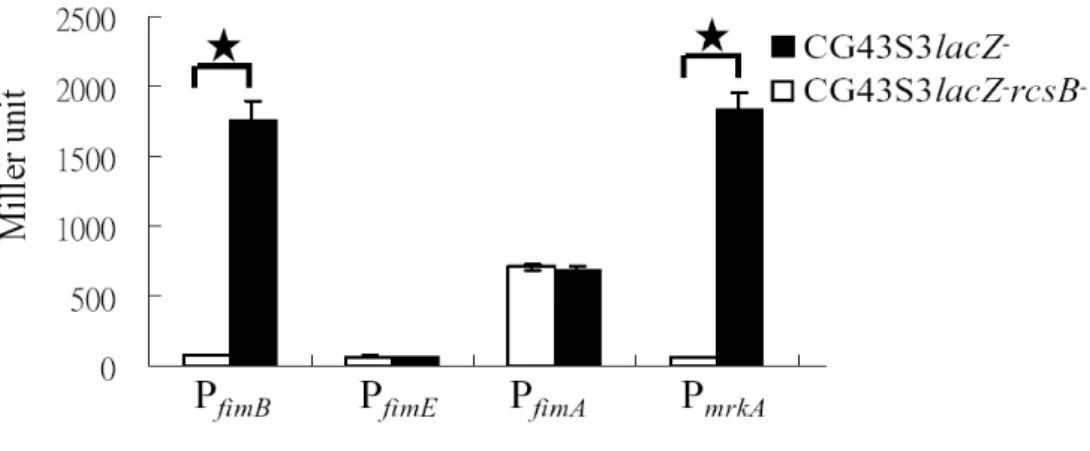
Promoter activity measurement as shown in Fig. 11, the deletion of rcsB appeared to reduce the activity of PfimB, which is consistent with the recent finding in E. coli5 that RcsB acts as a positive regulator for the expression of type 1 fimbriae. Nevertheless, rcsB deletion also reduced PmrkA activity (Fig. 11) indicating that the regulator responsible for the counter-expression of type 1 and 3 fimbriae has yet to be identified.
FIG. 11. Deletion effects of rcsB on the activity of PfimA, PfimB, PfimE, and
PmrkA. A significant difference (P < 0.001) was marked by a star.
FimK probably acts to regulate the expression of type 1 fimbriae and cps. The
fimK gene, located at the end of the type 1 fimbriae gene cluster, encodes a protein
with a DNA binding domain at the N-terminus and an EAL domain at the C-terminus (Fig. 12A), which suggesting a c-di-GMP phosphodiesterase activity of FimK protein. As shown in Fig. 12B, deletion of fimK increased the activity of type 1 fimbriae but decreased the CPS amount.
(A)
(B)
( )
FIG. 12. (A) Gene organization of type 1 fimbriae in K. pneumoniae NTUH-K2044.
FimK contains HTH domain (77.6% aligned) and EAL domain (100% aligned). (B) Assessment of type 1 fimbrial activity by yeast agglutination test and sedimentation test to determine the CPS synthesis (lower panel).
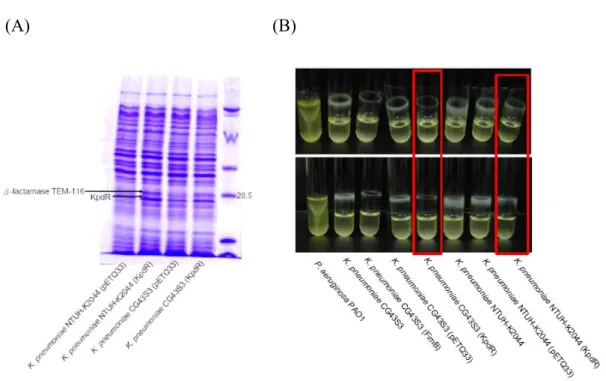
Overexpression of KpdR enhanced the synthesis of β-lactamse TEM-116 but reduced the biofilm formation. The overexpression of the 2CS response regulator
encoding gene kpdR, located upstream of the kpd fimbriae gene cluster, increased the synthesis of an unknown protein (Fig. 13A). Subsequently, the MS/MS analysis of the protein revealed a perfect match to β-lactamase TEM-116. Consistent with the report by Gallant CV et al., 2005(7), the increase synthesis of TEM-116 not only
increased the bacterial resistance to β-lactam antibiotic but also decreased the biofilm formation (Fig. 13B).
(A) (B)
FIG. 13. (A) Overexpression of KpdR in both K. pneumoniae NTUH-K2044 and K.
pneumoniae CG43S3. The bacteria were grown overnight in LB broth with addition
of 25 μg/ml Kanamycin and 1 mM IPTG at 37℃. (B) Biofilm formation analysis. Bacteria were grown at 37℃ in LB broth with addition of 25 μg/ml Kanamycin and 1 mM IPTG.
V. Literature Cited
1. Allen, B.L., G. –F. Gerlach, and S. Clegg. 1991. Nucleotide sequence and functions of mrk determinants necessary for expression of type 3 fimbriae in
Klebsiella pneumoniae. J. Bacteriol. 173:916-920.
2. Di Martino, P., Y. Bertin, J.P. Girardeau, V. Livrelli, B. Joly, and A.
Darfeuille-Michaud. 1995. Molecular characterization and adhesive properties
of CF29K, an adhesin of Klebsiella pneumoniae strains involve in nosocomial infections. Infect. Immun. 63:4336-4344.
3. Di Martino, P., V. Livrelli, D. Sirot, B. Joly, and A. Darfeuille-Michaud. 1996. A new fimbrial antigen harbored by CAZ-5/SHV-4-producing Klebsiella
pneumoniae strains involved in nosocomial infections. Infect. Immun.
64:2266-2273.
4. Fader, R. C., K. Gondesen, B. Tolley, D. G. Ritchie, and P. Moller. 1988. Evidence that in vitro adherence of Klebsiella pneumoniae to ciliated hamster tracheal cells is mediated by type 1 fimbriae. Infect. Immun. 56:3011-3013. 5. Fang, C.T., Y.P Chuang, C.T. Shun, S.C. Chang, and J.T. Wang. 2004. A
novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J. Exp. Med. 199:697-705.
6. Favre-Bonte, S., A. Darfeuille-Michaud, and C. Forestier. 1995. Aggregative adherence of Klebsiella pneumoniae to human intestine-407 cells. Infect. Immun. 63:1318-1328.
7. Gallant, C.V., C. Daniels, J.M. Leung, A.S. Ghosh, K.D. Young, L.P. Kotra,
and L.L. Burrows. 2005. Common β-lactamase inhibit bacterial biofilm
formation. Mol. Microbiol. 58:1012-1024.
8. Gerlach, G. –F., and S. Clegg. 1988. Characterization of two genes encoding antigenically distinct type-1 fimbriae of Klebsiella pneumoniae. Gene 64:231-240.
9. Gerlach, G. –F., S. Clegg, and B.L. Allen. 1989. Identification and characterization of the genes encoding the type 3 and type 1 fimbrial adhesins of Klebsiella pneumoniae. J. Bacteriol. 171:1261-1270.
10. Gerlach, G. B., L. Allen, and S. Clegg. 1989. Type 3 fimbriae among
Enterobacteriaceae and the ability of spermidine to inhibit MR/K
11. Gossert A.D., Bettendorff P., Puorger C., Vetsch M., Herrmann T.,
Glockshuber R., Wüthrich K. 2008. NMR structure of the Escherichia coli
type 1 pilus subunit FimF and its interactions with other pilus subunits. J Mol
Biol. 375(3):752-63.
12. Holden, N.J., M. Totsika, E. Mahler, A.J. Roe, K. Catherwood, K. Lindner,
U. Dobrindt, and D.L. Gally. 2006. Demonstration of regulatory cross-talk
between P fimbriae and type 1 fimbriae in uropathogenic Escherichia coli. Microbiol. 152: 1143-1153.
13. Holden, N.J., B.E. Uhlin, and D.L. Gally. 2001. PapB paralogues and their effect on the phase variation of type 1 fimbriae in Escherichia coli. Mol Microbiol 42: 319–330.
14. Huang, Y.J., C.C. Wu, M.J. Chen, C.P. Fung, and H.L. Peng. 2006. Characterization of the type 3 fimbriae with different MrkD adhesins: possible role of the MrkD containing an RGD motif. Biochem. Biophys. Res. Commun. 350: 537-542.
15. Humphries, A.D., M. Raffatellu, S. Winter, E.H. Weening, R.A. Kingsley, R.
Droleskey, S. Zhang, J. Figueiredo, S. Khare, J. Nunes, L.G. Adams, R.M. Tsolis, andA.J. Baumler. 2003. The use of flow cytometry to detect expression
of subunits encoded by 11 Salmonella enterica serotype Typhimurium fimbrial operons. Mol Microbiol. Jun; 48 (5):1357-76.
16. Johnson, D.E., C.V. Lockatell, ,R.G. Russell, J.R. Hebel, M.D. Island, and
A. Stapleton. 1998. Comparison of Escherichia coli strains recovered from
human cystitis and pyelonephritis infections in transurethrally challenged mice. Infect Immun 66: 3059–3065.
17. Klemm, P., B.J. Jorgensen, I. van Die, H. de Ree, and H. Bergmans. 1985. The fim genes responsible for synthesis of type 1 fimbriae in Escherichia coli, cloning and genetic organization. Mol. Gen. Genet. 199:410-414.
18. Klemm, P. 1986. Two regulatory fim genes, fimB and fimE, control the phase variation of type 1 fimbriae in Escherichia coli. EMBO J. 5:1389-1393.
19. Klemm, P. and G. Christiansen. 1987. Three fim genes required for the regulation of length and mediation of adhesion of Escherichia coli type 1 fimbriae. Mol. Gen. Genet. 2.8:439-445.
20. Laemmli U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227(5259):680-5.
21. Low A.S., Holden N., Rosser T., Roe A.J., Constantinidou C., Hobman J.L.,
Smith D.G., Low J.C., Gally D.L. 2006. Analysis of fimbrial gene clusters and
their expression in enterohaemorrhagic Escherichia coli O157:H7. Environ
22. Romling U., and D. Amikam. 2006. Cyclic di-GMP as a second messenger. Curr Opin Microbiol. 9:218-228.
23. Schwan W.R., Shibata S., Aizawa S., Wolfe A.J. 2007. The two-component response regulator RcsB regulates type 1 piliation in Escherichia coli. J
Bacteriol. 189(19):7159-63.
24. Sebghate, T.A., T.K. Korhonen, D.B. Kornick, and S. Clegg. 1998. Characterization of the type 3 fimbrial adhesins of Klebsiella strains. Infect. Immun. 66:2887-2894.
25. Soto, G.E., and S.J. Hultgren. 1999. Bacterial adhesins: common themes and variations in architecture and assembly. J Bacteriol. Feb; 181(4):1059-71. 26. Tarkkanen, A.M., R. Virkola, S. Clegg, and T.K. Korhonen. 1997. Binding
of the type 3 fimbriae of Klebsiella pneumoniae to human endothelial and urinary bladder cells. Infect. Immun. 65:1546-1549.
27. Townsend, S.M., N.E. Kramer, R. Edwards, S. Baker, N. Hamlin, M.
Simmonds, K. Stevens, S. Maloy, J. Parkhill, G. Dougan, and A.J. Baumler.
2001. Salmonella enterica serovar Typhi possesses a unique repertoire of fimbrial gene sequences. Infect Immun. May; 69(5):2894-901.
28. Van der Velden, A.W., A.J. Baumler, R.M Tsolis., and F. Heffron. 1998. Multiple fimbrial adhesins are required for full virulence of Salmonella
typhimurium in mice. Infect Immun. Jun; 66(6):2803-8.
29. Westerlund-Wikstrom, B., and T.K. Korhonen. 2005. Molecular structure of adhesin domains in Escherichia coli fimbriae. Int. J. Med. Microbiol. 295:479-486.
30. Xia, Y., D. Gally, K. Forsman-Semb, and B.E. Uhlin. 2000. Regulatory cross-talk between adhesin operons in Escherichia coli: inhibition of type 1 fimbriae expression by the Pap B protein. EMBO. 19:1450-1457.
31. Zhang, L., B. Foxman, S.D. Manning, P. Tallman, and C.F. Marrs. 2000 Molecular epidemiologic approaches to urinary tract infection gene discovery in uropathogenic Escherichia coli. Infect Immun 68: 2009–2015.
Coutiño-Rodríguez R, Hernández-Cruz P., Giles-Ríos H. 2001. Lectins in fruits having gastrointestinal activity: their participation in the hemagglutinating property of Escherichia coli O157:H7. Arch Med Res. 32(4):251-7.
VI. Self-Assessment
We have carried out the experiments as planned, published two papers (1, 2), and several manuscripts (3~5) are in the final stage to be submitted for publication. In addition, many interesting findings allow the anticipation of important contributions to unravel the regulation of fimbrial expression at genomic level, the cross-talk regulatory circuit in-between the nine fimbrial gene clusters, and also their roles in infections, which hold promise to provide intervening targets for antimicrobial agents.
1. Huang YJ, Wu CC, Chen MJ, Fung CP and Peng HL. (2006) Characterization of the type 3 fimbriae with different MrkD adhesins: possible role of the MrkD containing an RGD motif. BBRC 350:537-542.
2. Chang B, Huang YJ, et al., Measurement of adhesive force between a single type 3 fimbria and collagen IV using optic tweezers.(2006) BBRC 350: 33-38. 3. Huang YJ, Liao HW, Wu CC, and HL Peng (2007) Characterization of MrkF, a
component of type 3 fimbriae.
4. Huang YJ, Wu CC, and HL Peng (2008) A coordinated regulation of the expression of type 1 and 3 fimbriae in Klebsiella pneumoniae CG43.
5. Wu CC, Huang YJ, Chen YS, Chen HY, Chang HY, CP Fung, and HL Peng (2008) Distribution analysis of the putative fimbrial gene clusters in Klebsiella
pneumoniae clinical isolates.
6. Wu CC and HL Peng (2008) Role of FimK in regulation of the expression of type 1 fimbriae and cps genes in Klebsiella pneumoniae.