Mechanisms Involved in the Antiplatelet Activity of Rutin, a
Glycoside of the Flavonol Quercetin, in Human Platelets
J
OEN-R
ONGS
HEU,*
,†,§G
EORGEH
SIAO,
§P
O-H
SIUC
HOU,
†M
ING-Y
IS
HEN,
†ANDD
UEN-S
UEYC
HOU§Graduate Institutes of Medical Sciences and Pharmacology, Taipei Medical University, Taipei, Taiwan
The aim of this study was to systematically examine the inhibitory mechanisms of rutin, a well-known flavonoid in platelet aggregation. In this study, rutin concentration-dependently (250 and 290µM) inhibited platelet aggregation in human platelets stimulated by agonists (i.e., collagen). Rutin (250 and 290µM) did not significantly interfere with the binding of FITC-triflavin to the glycoprotein IIb/ IIIa complex in human platelets. Rutin (250 and 290 µM) markedly inhibited intracellular Ca2+ mobilization and thromboxane A2 formation in human platelets stimulated by collagen. Rapid
phosphorylation of a platelet protein ofMr47000 (P47), a marker of protein kinase C activation, was
triggered by collagen (1µg/mL). This phosphorylation was markedly inhibited by rutin (250 and 290
µM). On the other hand, rutin (250 and 290µM) did not significantly increase the formations of cyclic AMP and nitric oxide/cyclic GMP in platelets. In conclusion, these results indicate that the antiplatelet activity of rutin may involve the following pathways: rutin inhibited the activation of phospholipase C, followed by inhibition of protein kinase C activity and thromboxane A2formation, thereby leading to
inhibition of the phosphorylation of P47 and intracellular Ca2+mobilization, finally resulting in inhibition
of platelet aggregation.
KEYWORDS: Rutin; platelet aggregation; phospholipase C; protein kinase C; thromboxane A2 INTRODUCTION
It is known that polyphenols are a large family of natural
compounds derived from various plant sources. Recent studies
in vitro and in vivo show that some polyphenols exhibit
antioxidant and free radical-scavenging properties (1). Therefore,
dietary polyphenolics, which possess antioxidant activity, may
play a role in human health, particularly in diseases believed to
involve, in part, oxidation, such as coronary heart disease,
inflammation, and mutagenesis leading to carcinogenesis (2, 3).
Rutin, a glycoside of the flavonol quercetin, is one of the major
polyphenols. This compound has been extensively studied and
is known to exhibit antitumor (4), anti-inflammatory (5),
antiplatelet (6), and vasodilation (7) activities.
Intravascular thrombosis is one generator of a wide variety
of cardiovascular diseases. The initiation of an intraluminal
thrombosis is believed to involve platelet adherence and
aggregation. In normal circulation, platelets cannot aggregate
by themselves. However, when a blood vessel is damaged,
platelets adhere to the disrupted surface, and the adherent
platelets release some biologically active constituents and
aggregate (8). Thus, platelet aggregation may play a crucial role
in the atherothrombotic process. Indeed, antiplatelet agents (e.g.,
ticlopidine and aspirin) have been shown to reduce the incidence
of stroke in high-risk patients (9).
Rutin has been shown to inhibit platelet aggregation in both
in vitro and ex vivo experiments (6, 7, 10). However, no data
are available concerning the detailed effects of rutin in platelet
aggregation. Therefore, the inhibitory mechanisms of rutin
underlying the signaling pathways in platelets remain obscure.
We therefore systematically examined the influence of rutin in
washed human platelets and utilized the findings to characterize
the mechanisms involved in this influence.
MATERIALS AND METHODS
Materials. Collagen (type I, bovine achilles tendon), rutin, sodium
citrate, luciferin-luciferase, indomethacin, myoinositol, prostaglandin E1(PGE1), arachidonic acid, apyrase, and heparin were purchased from Sigma Chemical Co. (St. Louis, MO). Fura 2-AM and fluorescein isothiocyanate (FITC) were purchased from Molecular Probes (Eugene, OR). Trimeresurus flaVoViridis venom was purchased from Latoxan (Rosans, France). Myo-2-[3H]inositol and thromboxane B
2, cyclic AMP, andGMP EIA kits were purchased from Amersham (Buckinghamshire, HP, U.K.). VCl3was obtained from Aldrich (Milwaukee, WI). Rutin was dissolved in dimethyl sulfoxide (DMSO) and stored at -4°C until use.
Preparation of Human Platelet Suspensions. Human platelet
suspensions were prepared as previously described (11). In this study, human volunteers gave informed consent. In brief, blood was collected from healthy human volunteers who had taken no medicine during the preceding 2 weeks and was mixed with acid/citrate/glucose (9:1, v/v). * Address correspondence to this author at the Graduate Institute of
Medical Sciences, Taipei Medical University, No. 250 Wu-Hsing St., Taipei 110, Taiwan (telephone/fax +886-2-27390450; e-mail [email protected]).
†Graduate Institute of Medical Sciences. §Graduate Institute of Pharmacology.
4414
J. Agric. Food Chem. 2004, 52, 4414
−
4418
10.1021/jf040059f CCC: $27.50 © 2004 American Chemical Society Published on Web 06/19/2004
After centrifugation at 120g for 10 min, the supernatant (platelet-rich plasma; PRP) was supplemented with PGE1(0.5µM) and heparin (6.4 IU/mL), then incubated for 10 min at 30°C, and centrifuged at 500g for 10 min. The washed platelets were finally suspended in Tyrode’s solution containing bovine serum albumin (BSA) (3.5 mg/mL) and adjusted to a concentration of 4.5 × 108 platelets/mL. The final concentration of Ca2+in Tyrode’s solution was 1 mM.
Platelet Aggregation. The turbidimetric method was applied to
measure platelet aggregation (11), using a Lumi-Aggregometer (Payton, Canada). Platelet suspensions (4.5× 108platelets/mL, 0.4 mL) were prewarmed to 37°C for 2 min (stirring at 1200 rpm) in a silicone-treated glass cuvette. Rutin (250 and 290µM) was added 3 min before
the addition of platelet aggregation inducers (i.e., collagen). The reaction was allowed to proceed for at least 6 min, and the extent of aggregation was expressed in light transmission units. While ATP release was measured, 20µL of a luciferin-luciferase mixture was added 1 min
before the addition of agonists, and ATP release was compared with that of the control.
Analysis of the Platelet Surface Glycoprotein IIb/IIIa Complex by Flow Cytometry. Triflavin, a specific fibrinogen receptor
(glyco-protein IIb/IIIa complex) antagonist, was prepared as previously described (12). Fluorescence-conjugated triflavin was also prepared as previously described (13). The final concentration of FITC-conjugated triflavin was adjusted to 1 mg/mL. Human platelet suspensions were prepared as described above. Aliquots of platelet suspensions (4.5× 108/mL) were preincubated with rutin (250 and 290µM) for 3 min, followed by the addition of 2µL of FITC-triflavin. The suspensions
were then incubated for another 5 min, and the volume was adjusted to 1 mL/tube with Tyrode’s solution. The suspensions were then assayed for fluorescein-labeled platelets with a flow cytometer (Becton Dick-inson, FACScan Systems, San Jose, CA). Data were collected from 50000 platelets per experimental group. All experiments were repeated at least four times to ensure reproducibility.
Measurement of Platelet [Ca+2]i Mobilization by Fura 2-AM Fluorescence. Citrated whole blood was centrifuged at 120g for 10
min. The supernatant was protected from light and incubated with Fura 2-AM (5µM) at 37°C for 1 h. Human platelets were then prepared as described above. Finally, the external Ca2+concentration of the platelet suspensions was adjusted to 1 mM. The rise in [Ca2+]i was measured using a fluorescence spectrophotometer (CAF 110, Jasco) at excitation wavelengths of 340 and 380 nm and at an emission wavelength of 500 nm. [Ca2+]i was calculated from the fluorescence, using 224 nM as the Ca2+-Fura 2 dissociation constant (14).
Measurement of Thromboxane B2Formation. Washed human
platelet suspensions (4.5× 108/mL) were preincubated for 3 min in the presence or absence of rutin (250 and 290µM) before the addition
of collagen (1µg/mL). Six minutes after the addition of agonists, 2
mM EDTA and 50 µM indomethacin were added to the reaction
suspensions. The vials were then centrifuged for 3 min at 15000g. The thromboxane B2(TxB2) levels of the supernatants were measured using an EIA kit according to the instructions of the manufacturer.
Estimation of Platelet Cyclic AMP and Cyclic GMP Formations.
The method of Karniguian et al. (15) was followed. In brief, platelet suspensions were warmed to 37°C for 1 min, and then either PGE1 (10µM), nitroglycerin (10 µM), or rutin (250 and 290 µM) was added
and incubated for 6 min. The incubation was stopped, and the solution was immediately boiled for 5 min. After cooling to 4°C, the precipitated protein was collected as sediment after centrifugation. Fifty microliters of supernatant was used to determine the cyclic AMP and cyclic GMP contents by EIA kits following acetylation of the samples as described by the manufacturer.
Estimation of Nitrate in Human Platelet Suspensions. NO was
assayed in platelet suspensions as previously described (16). In brief, platelet suspensions (1× 109/mL) were preincubated with collagen (1
µg/mL) or rutin (250 and 290 µM) for 3 min, respectively, followed
by centrifugation. The amount of nitrate in the platelet suspensions (10µL) was measured by adding a reducing agent (0.8% VCl3in 1 M HCl) to the purge vessel to convert nitrate to NO, which was stripped from the platelet suspensions by a helium purge gas. The NO was then drawn into a Sievers nitric oxide analyzer (Sievers 280 NOA, Sievers
Inc., Boulder, CO). Nitrate concentrations were calculated by com-parison with standard solutions of sodium nitrate.
Measurement of Protein Kinase C Activity. Washed human
platelets (2 × 109/mL) were incubated for 60 min at 37 °C with phosphorus-32 (0.5 mCi/mL). Platelet suspensions were next washed twice with Tris-saline buffer. The32P-labeled platelets were preincu-bated with rutin (250 and 290µM) in an aggregometer at 37°C for 3 min, and then collagen (1µg/mL) was added for 1 min to trigger protein
kinase C activation. Activation was terminated by the addition of Laemmli sample buffer, followed by electrophoresis (12.5%, w/v) as described previously (17). The gels were dried, and the relative intensities of the radioactive bands were analyzed using a Bio-imaging analyzer system (FAL2000, Fuji, Tokyo, Japan) and expressed as photostimulated luminescence (PSL) per millimeter.
Statistical Analysis. Experimental results are expressed as the means
( SEM and are accompanied by the number of observations. Data were assessed by the method of analysis of variance (ANOVA). If this analysis indicated significant differences among the group means, then each group was compared using the Newman-Keuls method. A p value of <0.05 was considered to be statistically significant.
RESULTS
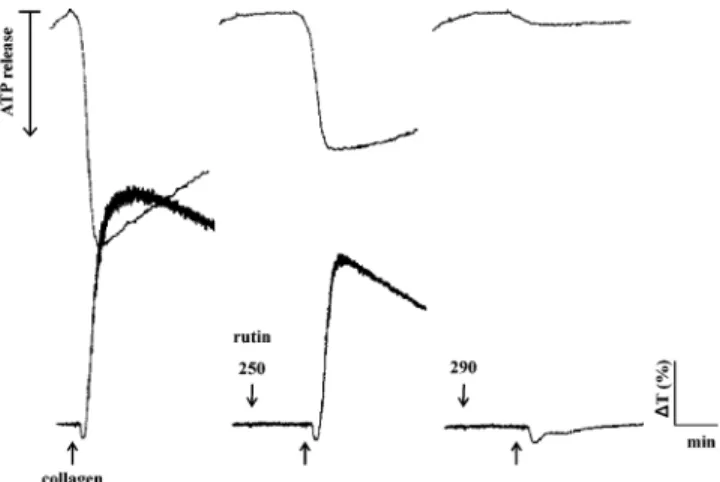
Effect of Rutin on Platelet Aggregation in Human Platelet
Suspensions. Rutin (250 and 290
µM)
concentration-depen-dently inhibited platelet aggregation stimulated by collagen (1
µg/mL) (Figure 1), thrombin (0.1 unit/mL), and arachidonic
acid (60
µM) in human platelets (data not shown). Furthermore,
rutin inhibited the ATP-release reaction when stimulated by
agonists (i.e., collagen) (Figure 1). On the other hand, its solvent
control (0.5% DMSO) did not significantly affect platelet
aggregation stimulated by agonists (data not shown). In the
following experiments, we used collagen as an agonist to explore
the inhibitory mechanisms of rutin in platelet aggregation.
Effect of Rutin on Collagen-Induced Conformational
Change of the Glycoprotein IIb/IIIa Complex in Human
Platelets. Triflavin is an Arg-Gly-Asp-containing antiplatelet
peptide purified from T. flaVoViridis snake venom (12). Triflavin
inhibits platelet aggregation through direct interference with
fibrinogen binding to the glycoprotein IIb/IIIa complex (RIIb
β
3integrin) (12). There is now a multitude of evidence suggesting
that the binding of fibrinogen to the glycoprotein IIb/IIIa
complex is the final common pathway for agonist-induced
platelet aggregation. Therefore, we decided to further evaluate
Figure 1. Tracing curves of rutin on collagen (1 µg/mL)-induced
aggregation and ATP release in washed human platelets. Platelets were preincubated with rutin (250 and 290 µΜ) for 3 min, and then collagen was added to trigger aggregation (lower tracings) and ATP release (upper tracings). A luciferin−luciferase mixture (20 µL) was added 1 min before collagen addition to measure the ATP-release reactions. The profiles are representative examples of five similar experiments.
whether rutin binds directly to the platelet glycoprotein IIb/IIIa
complex, leading to inhibition of platelet aggregation induced
by agonists.
In this study, the relative intensity of fluorescence of
FITC-triflavin (2
µg/mL) bound directly to collagen (1
µg/mL)-activated platelets was 581.5 ( 54.2 (Figure 2A), and it was
markedly reduced in the presence of 5 mM EDTA (negative
control, 75.0 ( 1.9, p < 0.001) (Figure 2B). Rutin (250 and
290
µM) did not significantly inhibit FITC-triflavin binding
to the glycoprotein IIb/IIIa complex in platelet suspensions (250
µM, 520.8 ( 45.6; 290 µM, 552.3 ( 64.4) (Figure 2C,D),
indicating that the mechanism of rutin’s inhibitory effect on
platelet aggregation does not involve directly binding to the
glycoprotein IIb/IIIa complex.
Effect of Rutin on [Ca
2+]i Mobilization. As shown in
Figure 3, collagen (1
µg/mL) evoked an increase of [Ca
2+]i
from 28.1 ( 4.5 to 287.5 ( 27.6 nM. This collagen-evoked
increase in [Ca
2+]i was markedly inhibited in the presence of
rutin (250
µΜ, 62.7 ( 8.1%; 290 µΜ, 81.1 ( 6.3%). This
suggests that rutin exerts an inhibitory effect on [Ca
2+]i
mobilization in human platelets stimulated by collagen.
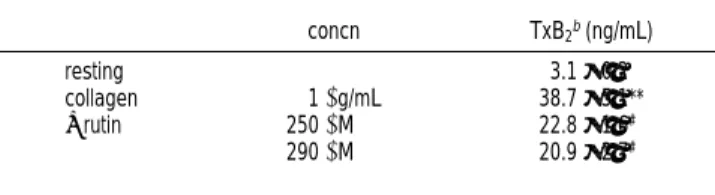
Effect of Rutin on Thromboxane B
2Formation. As shown
in Table 1, resting platelets produced relatively little TxB2
compared with collagen-activated platelets. PGE1
(10
µM)
inhibited TxB2
formation in collagen-activated platelets by 81%
(data not shown). Furthermore, results obtained using various
concentrations of rutin indicated that rutin (250 and 290
µM)
concentration-dependently inhibited TxB2
formation in platelets
stimulated by collagen (1
µg/mL) about 41 and 46%,
respec-tively (Table 1). This result suggests that rutin exerts an
inhibitory effect on TxA2
formation.
Effect of Rutin on Cyclic AMP, Cyclic GMP, and Nitrate
Formations in Washed Human Platelets. The level of cyclic
AMP in unstimulated platelets was low (2.5 ( 0.8 pmol/mL).
Addition of PGE1
(10
µM) increased the cyclic AMP level to
25.5 ( 1.0 pmol/mL (Table 2). When platelet suspensions were
preincubated with various concentrations of rutin (250 and 290
µM) for 3 min at 37
°C, we found that rutin did not significantly
increase the cyclic AMP level (Table 2). We also performed
similar studies measuring cyclic GMP response. The level of
cyclic GMP in unstimulated platelets was very low, but when
nitroglycerin (10
µΜ) was added to the platelet suspensions,
the cyclic GMP level increased from the resting level to 1.2 (
0.3 pmol/mL (Table 2). However, addition of rutin (250 and
290
µM) resulted in no significant increase in platelet cyclic
GMP levels (0.5 ( 0.1 and 0.5 ( 0.0 pmol/mL) (Table 2).
On the other hand, NO was quantified using a sensitive and
specific ozone redox-chemiluminescence detector. As shown
in Table 2, collagen (1
µg/mL) caused a
∼1.8-fold rise in nitrate
Figure 2. Flow cytometric analysis of FITC−triflavin binding to human platelets in the absence or presence of various concentrations of rutin (250 and 290 µΜ): (A) solid line represents the fluorescence profiles of FITC−triflavin (2 µg/mL) in the absence of rutin as a positive control; (B) in the presence of EDTA (5 mM) as the negative control; or in the presence of rutin (C) (250 µΜ) and (D) (290 µΜ). The profiles are representative examples of four similar experiments.
Figure 3. Effect of rutin on collagen-induced intracellular Ca2+mobilization in Fura 2-AM-loaded human platelets. Platelet suspensions were prein-cubated with Fura 2-AM (5 µΜ), followed by the addition of collagen (1
µg/mL) in the absence or presence of rutin (250 and 290 µΜ), which
was added 3 min prior to the addition of collagen. The profiles are representative examples of four similar experiments.
Table 1. Effect of Rutin on Thromboxane B2Formation Stimulated by Collagen in Washed Human Plateletsa
concn TxB2b(ng/mL)
resting 3.1±0.2
collagen 1 µg/mL 38.7±5.1**
+rutin 250 µM 22.8±1.6#
290 µM 20.9±2.7#
aPlatelet suspensions were preincubated with rutin (250 and 290 µM) for 3
min at 37°C, and then collagen (1 µg/mL) was added to trigger thromboxane B2
formation. Data are presented as the means±SEM (n ) 5).b**, p < 0.001, as
compared with the resting group;#, p < 0.05 as compared with the collagen group. Table 2. Effect of Rutin on Cyclic AMP, Nitrate, and Cyclic GMP Formations in Washed Human Plateletsa
concn cyclic AMPb (pmol/mL) nitrateb (µM) cyclic GMP (pmol/mL) resting 2.5±0.8 4.4±0.3 0.5±0.1 prostaglandin E1 10 µM 25.5±1.0** collagen 1 µg/mL 7.8±0.3** nitroglycerin 10 µM 1.2±0.2* rutin 250 µM 2.1±1.0 3.9±0.3 0.5±0.1 290 µM 1.4±0.4 4.3±0.4 0.5±0.0
aPlatelet suspensions were preincubated with rutin (250 and 290 µΜ) for 3
min at 37°C. Addition of prostaglandin E1, collagen, and nitroglycerin into the
platelet suspensions served as positive control of cyclic AMP, nitrate, and cyclic GMP formations, respectively. Data are presented as the means±SEM (n ) 4).
formation, compared to that in resting platelets. In the presence
of rutin (250 and 290
µM), nitrate production did not
signifi-cantly increase after incubation with platelets for 3 min (Table
2). These results imply that the antiplatelet activity of rutin may
not act through stimulation of the NO/cyclic GMP pathway in
human platelets.
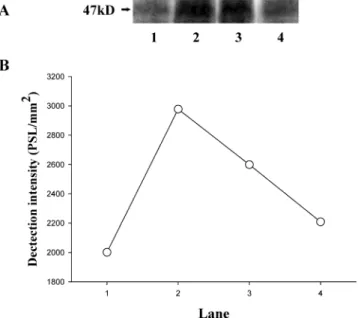
Effect of Rutin on Collagen-stimulated Phosphorylation
of the 47-kDa Protein. Stimulation of platelets with a number
of different agonists (i.e., collagen) induces activation of protein
kinase C, which then phosphorylates proteins of Mr
40000-47000 in addition to other proteins (18). In this study,
phos-phorylation experiments were performed to examine the role
of rutin in the activation of protein kinase C in human platelets.
When collagen (1
µg/mL) was added to human platelets
prelabeled with
32PO4
for 2 min, a protein with an apparent Mr
of 47000 (P47) was predominately phosphorylated as compared
with resting platelets (Figure 4). On the other hand, rutin (250
and 290
µM) concentration-dependently inhibited the
phos-phorylation of P47 in human platelets stimulated by collagen
(Figure 4). In this study, the extent of radioactivity in P47 was
expressed as a relative detection density (PSL/mm
2) of the
radioactive bands (Figure 4B).
DISCUSSION
The results obtained from this study demonstrate that rutin
provides antiplatelet activity. The principal objective of this
study was to describe the detailed mechanisms involved in the
inhibition of agonist-induced human platelet aggregation by
rutin. This inhibitory effect of rutin was demonstrable with the
use of various agonists (i.e., collagen). In this study, platelet
aggregation induced by these agonists (i.e., collagen) appeared
to be affected in the presence of rutin. Therefore, this partly
implies that rutin may affect Ca
2+release from intracellular Ca
2+storage sites (i.e., dense tubular systems or dense bodies) (Figure
3), and this is in accord with the concept that intracellular Ca
2+release is responsible for platelet aggregation (19).
Although the action mechanisms of various platelet
aggrega-tion agonists, such as collagen, thrombin, and arachidonic acid,
differ, rutin significantly inhibited platelet aggregation stimulated
by all of them. This implies that rutin may block a common
step shared by these inducers. These results also indicate that
the site of action of rutin is not at the receptor level of individual
agonists. Triflavin acts by binding to the glycoprotein IIb/IIIa
complex on the platelet surface membrane, resulting in
interfer-ence with the interaction of fibrinogen with its specific receptor
(12). In this study, we found that rutin did not significantly affect
FITC-triflavin binding to the glycoprotein IIb/IIIa complex,
indicating that the antiplatelet activity of rutin is possibly not
directly due to interference with the binding of fibrinogen to
its specific receptor on the platelet membrane.
Stimulation of platelets by agonists (i.e., collagen) results in
phospholipase C-catalyzed hydrolysis of the minor plasma
membrane phospholipid, phosphatidylinositol 4,5-bisphosphate,
with concomitant formation of inositol 1,4,5-trisphosphate and
diacylglycerol (20). There is strong evidence that inositol
1,4,5-trisphosphate induces the release of Ca
2+from intracellular
stores (21). Diacylglycerol activates protein kinase C, inducing
protein phosphorylation and a release reaction. Rutin
signifi-cantly inhibited collagen-induced activation of protein kinase
C (Figure 4). Furthermore, our study revealed that
phospho-inositide breakdown of collagen (1
µg/mL)-activated platelets
was significantly inhibited by rutin (250
µM) (data not shown),
suggesting that lowering of intracellular Ca
2+mobilization by
rutin is related to inhibition of phospholipase C. Moreover, TxA2
is an important mediator of the release reaction and aggregation
of platelets (22). Collagen-induced TxB2
formation, a stable
metabolite of TxA2, was markedly inhibited by rutin (250 and
290
µM) (Table 1). It has been demonstrated that
phospho-inositide breakdown can induce TxA2
formation via free
arachidonic acid release by diglyceride lipase or by endogenous
phospholipase A2
from membrane phospholipids (23). Thus, it
seems likely that TxB2
formation plays a role in mediating the
inhibitory effect of rutin on human platelets.
The activation of human platelets is inhibited by two
intracellular pathways regulated by either cyclic AMP or NO/
cyclic GMP (24). The importance of cyclic AMP in modulating
platelet reactivity is well established (15). In addition to
inhibiting most platelet responses, elevated levels of cyclic AMP
decrease intracellular Ca
2+concentration by the uptake of Ca
2+into the dense tubular system (24) and negatively affect the
action of protein kinase C (24). Signaling by NO/cyclic GMP
somehow interferes with the agonist-stimulated phosphoinositide
turnover that creates Ca
2+-mobilizing second messengers and
protein kinase C (25). However, neither cyclic AMP nor NO/
cyclic GMP mediated the inhibitory effect of rutin on
agonist-induced platelet aggregation.
In conclusion, the most important observations of this study
suggest that rutin markedly inhibits agonist-induced human
platelet aggregation. This inhibitory effect may possibly involve
the following mechanisms: rutin inhibits the activation of
phospholipase C, followed by inhibition of protein kinase C
activation and thromboxane A2
formation, thereby leading to
inhibition of the phosphorylation of P47 and intracellular Ca
2+mobilization, finally resulting in inhibition of platelet
aggrega-tion. These findings suggest that rutin may be an effective tool
in treating thromboembolic-related disorders.
Figure 4. Effect of rutin on phosphorylation of a protein of Mr47000 (P47) in human platelets challenged with collagen. Platelets were preincubated with rutin (250 and 290 µΜ) before challenge with collagen (1 µg/mL): (lane 1) platelets with Tyrode’s solution only (resting group); (lane 2) platelets with collagen (1 µg/mL); (lane 3) platelets with rutin (250 µΜ); (lane 4) platelets with rutin (290 µΜ) for 3 min followed by the addition of collagen (1 µg/mL). (A) The profiles are representative examples of four similar experiments. The arrow indicates a protein of Mr 47000 (P47). (B) The relative detection densities of the radioactive bands are expressed as PSL/mm2(PSL, photostimulated luminescence).
LITERATURE CITED
(1) Visioli, F.; Galli, C. Olive oil polyphenols and their potential effects on human health. J. Agric. Food Chem. 1998, 46, 4292-4296.
(2) Kushi, L. H.; Lenart, E. B.; Willet, W. C.; Sacks, S.; Trichopou-lou, A.; Drescher, G.; FerroLuzzi, A.; Helsin, E.; Trichopoulos, D. Mediterranean diet pyramida cultural model for healthy eating. Am. J. Clin. Nutr. 1995, 61, 1402S-1406S.
(3) Hertog, M. G. L.; Hollman, P. C. H.; Venema, D. P. Optimization of a quantitative HPLC determination of potentially anticarci-nogenic flavonoids in vegetables and fruit. J. Agric. Food Chem.
1992, 40, 1591-1598.
(4) Deschner, E. E.; Ruperto, J.; Wong, G.; Newmark, H. L. Quercetin and rutin as inhibitors of azoxymethanol-induced colonic neoplasia. Carcinogenesis 1991, 12, 1193-1196. (5) Aleksandrov, P. N.; Speranskaia, T. V.; Bobkov, I. G.;
Zagor-evskii, V. A.; Zykov, D. A. Effect of rutin and esculamine on models of aseptic inflammation. Farmakol. Toksikol. 1986, 49, 84-86.
(6) Swies, J.; Robak, J.; Dabrowski, L.; Duniec, Z.; Michalska, Z.; Gryglewski, R. J. Antiaggregatory effects of flavonoids in vivo and their influence on lipoxygenase and cyclooxygenase in vitro. Pol. J. Pharmacol. Pharm. 1984, 36, 455-463.
(7) Chung, M. I.; Gan, K. H.; Lin, C. N.; Ko, F. N.; Teng, C. M. Antiplatelet effects and vasorelaxing action of some constitutes of Formosan plants. J. Nat. Prod. 1993, 56, 929-934. (8) Sheu, J. R.; Chao, S. H.; Yen, M. H.; Huang, T. F. In vivo
antithrombotic effect of triflavin, an Arg-Gly-Asp containing peptide on platelet plug formation in mesenteric microvessels of mice. Thromb. Haemost. 1994, 72, 617-621.
(9) Hass, W. K.; Easton, J. D.; Adams, H. P.; Pryse-Philips, W.; Molony, B. A.; Anderson, S.; Kamm, B. A randomized trial comparing ticlopidine hydrochloride with aspirin for the preven-tion of stroke in high-risk patients. New Engl. J. Med. 1989, 321, 501-507.
(10) Gryglewski, R. J.; Korbut, R.; Robak, J.; Swies, J. On the mechanism of antithrombotic action of flavonoids. Biochem. Pharmacol. 1987, 36, 317-322.
(11) Sheu, J. R.; Lee, C. R.; Lin, C. C.; Kan, Y. C.; Lin, C. H.; Hung, W. C.; Lee, Y. M.; Yen, M. H. The antiplatelet activity of PMC, a potent R-tocopherol analogue, is mediated through inhibition of cyclo-oxygenase. Br. J. Pharmacol. 1999, 127, 1206-1212. (12) Sheu, J. R.; Teng, C. M.; Huang, T. F. Triflavin, an RGD-containing antiplatelet peptide, binds to GP IIIa of ADP-stimulated platelets. Biochem. Biophys. Res. Commun. 1992, 189, 1236-1242.
(13) Sheu, J. R.; Lin, C. H.; Peng, C. H.; Huang, T. F. Triflavin, an Arg-Gly-Asp-containing peptide, inhibits the adhesion of tumor cells to matrix protein via binding to multiple integrin receptors
expressed on human hepatoma cells. Proc. Soc. Exp. Biol. Med.
1996, 213, 71-79.
(14) Grynkiewicz, G.; Poenie, M.; Tsien, R. Y. A new generation of Ca2+indicator with greatly improved fluorescence properties.
J. Biol. Chem. 1985, 260, 3440-3450.
(15) Karniguian, A.; Legrand, Y. J.; Caen, J. P. Prostaglandins: specific inhibition of platelet adhesion to collagen and relation-ship with cyclic AMP level. Prostaglandins 1982, 23, 437-457. (16) Sheu, J. R.; Hung, W. C.; Su, C. H.; Lin, C. H.; Lee, L. W.; Lee, Y. M.; Yen, M. H. The antiplatelet activity of Escherichia coli lipopolysaccharide is mediated through a nitric oxide/cyclic GMP pathway. Eur. J. Haematol. 1999, 62, 317-326. (17) Grabarek, J.; Raychowdhury, M.; Ravid, K.; Kent, K. C.;
Newman, P. J.; Ware, J. A. Identification and functional characterization of protein kinase C isozymes in platelets and HEL cells. J. Biol. Chem. 1992, 267, 10011-10017. (18) Siess, W.; Lapetina, E. G. Platelet aggregation induced by alpha
2-adrenoceptor and protein kinase C activation. A novel syner-gism. Biochem. J. 1989, 263, 377-385.
(19) Charo, I. F.; Feinman, R. D.; Detwiler, T. C. Inhibition of platelet secretion by an antagonist of intracellular calcium. Biochem. Biophys. Res. Commun. 1976, 72, 1462-1467.
(20) Kirk, C. J.; Creba, J. A.; Downes, C. P.; Michell, R. H. Hormone-stimulated metabolism of inositol lipids and its relationship to hepatic receptor function. Biochem. Soc. Trans. 1981, 9, 377-379.
(21) Berridge, M. J. Rapid accumulation of inositol trisphosphate reveals that agonists hydrolyse polyphosphoinositides instead of phosphatidylinositol. Biochem. J. 1983, 212, 249-258. (22) Hornby, E. J. Evidence that prostaglandin endoperoxides can
induce platelet aggregation in the absence of thromboxane A2 production. Biochem. Pharmacol. 1982, 31, 1158-1160. (23) McKean, M. L.; Smith, J. B.; Silver, W. J. Formation of
lysophosphatidylcholine in human platelets in response to thrombin. J. Biol. Chem. 1981, 256, 1522-1524.
(24) Walter, U.; Eigenthaler, M.; Geiger, J.; Reinhard, M. Role of cyclic nucleotide-dependent protein kinases and their common substrate VASP in the regulation of human platelets. AdV. Exp. Med. Biol. 1993, 344, 237-249.
(25) McDonald, L. J.; Murad, F. Nitric oxide and cyclic GMP signaling. Proc. Soc. Exp. Biol. Med. 1996, 211, 1-6.
Received for review February 6, 2004. Revised manuscript received April 27, 2004. Accepted April 27, 2004. This work was supported by a grant from the National Science Council of Taiwan (NSC92-2320-B-038-026).