利用人類脫氫表雄甾酮亞硫酸基轉移酵素之序列與結構比對探討其催化機制與功能
108
0
0
全文
(2) 利用人類脫氫表雄甾酮亞硫酸基轉移酵素之序列與結構比 對探討其催化機制與功能 From Sequence and Structure of Human Dehydroepiandrosterone Sulfotransferase to Its Mechanism of Action and Function. 研 究 生:呂陸宜. Student: Lu-Yi Lu. 指導教授:楊裕雄 教授. Advisor: Prof. Yuh-Shyong Yang. 國立交通大學 生物科技學系 碩 士 論 文. A Thesis Submitted to Department of Biological Science and Technology National Chiao Tung University in Partial Fulfillment of the Requirements for the Degree of Master of Science in Biologic Science and Technology May 2006 Hsinchu, Taiwan, Republic of China 中華民國九十五年五月.
(3) 利用人類脫氫表雄甾酮亞硫酸基轉移酵素之序列與結構比對探討 其催化機制與功能 學生: 呂陸宜. 指導教授: 楊裕雄 教授 國立交通大學生物科技學系碩士班 摘要. 本研究利用人類脫氫表雄甾酮亞硫酸基轉移酵素之胺基酸序列與已知 結晶結構來探討其催化機制與功能。亞硫酸基轉移酵素目前大部分已知為 同型二聚體,在生理上主要將生物分子亞硫酸化,進而調控許多生理反應 與功能。在此研究中,首先為了要瞭解四級結構對此酵素活性之影響,因 而藉由定點突變法製備單體之人類脫氫表雄甾酮亞硫酸基轉移酵素與經由 膠體過濾分析,我們成功得到單體的亞硫酸基轉移酵素。並由圓二色偏光 光譜法來觀測單體與雙體在二級與三級結構上之差異。另外,利用酵素動 力學分析探討其催化機制,受質抑制作用,與受質結合能力分析。此外, 藉由熱穩定度與尿素變性分析單體與雙體在構型上的穩定度。另一方面, 亞硫酸基轉移酵素有一個共同特徵:受質抑制反應。藉由交叉比對分析其 結晶結構之二體與三體錯合物,推測可能調控受質抑制反應與受質位向之 胺基酸,進而探討受質抑制反應的起源與機制。在此研究中,我們成功地 瞭解受質抑制反應與受質位向之間相關性。此外,利用定點突變法與其進 一步分析指出,此胺基酸可能在其他亞硫酸基轉移酵素中之相對位置對於 受質抑制反應扮演相類似的調控角色。 i.
(4) From Sequence and Structure of Human Dehydroepiandrosterone Sulfotransferase to Its Mechanism of Action and Function Student: Lu-Yi Lu. Advisor: Prof. Yuh-Shyong Yang. Department of Biological Science and Technology and Institute of Biochemical Engineering, National Chiao Tung University, Hsinchu, Taiwan, ROC ABSTRACT In this research we deduce the human dehydroepiandrosterone sulfotransferase (SULT2A1) catalytic mechanism and function by the comparison of its amino acid sequences and spatially resolved crystal structures. Sulfotransferase (SULT), mostly known to be homodimer in solution, belongs to an enzyme family that is responsible for the sulfonation of biomolecules and then the regulation of many biological functions. In order to study the effect of quaternary structure on the activity of the enzyme, SULT2A1 monomer mutant is prepared by site-directed mutagenesis and confirmed by gel filtration. Further, the differences of secondary and tertiary structures between dimer and monomer, respectively, are investigated by circular dichroism spectrum. Enzyme kinetics, substrate inhibition and substrate binding affinity are facilitated to deduce the catalytic mechanism and actions. Thermal inactivation and conformational stabilities of dimer and monomer are also determined by enzyme assay and urea unfolding analysis individually. Another general feature of sulfotransferase is substrate inhibition. Following the analysis of the structures of binary and ternary complexes of sulfotransferase, we are able to elucidate the specific amino acids responsible for substrate inhibition and binding orientations. And then we propose the origin and mechanism of substrate inhibition and uncover the relationship between the substrate inhibition and binding orientations. Site-directed mutagenesis and resulting analysis confirm that the involvement of specific amino acid residue in substrate inhibition may be a general feature in all cytosolic sulfotransferases.. ii.
(5) ACKNOWLEDGEMENT 終於熬到了這一刻了!雖然辛苦但也著實令人興奮。回想過去從懵懂的專題 生到現在略通一二的碩士畢業生,一路走來真是辛苦啊。回首來時路,要感謝的 人真的太多了,首先要感謝我的指導教授楊裕雄老師,也真多虧有老師當初能收 留一個完全沒有背景知識的專題生,讓我能勉勵自己要汲取更多的知識。而也是 因為老師的指導之下,我的思考訓練才有得以呼吸的空間。更感謝的是老師對我 的信任,能容忍我一次又一次的失敗,適時地勉勵與關懷,創造了一個絕佳的實 驗環境與氣氛,而使學生的時間與空間能更彈性自由。 此外,也要感謝簡慶德老師,黃鎮剛老師,劉明毅老師能擔任我的碩士班口 試委員,對我的論文能提出諸多建議以使其更具完整,其用心與指導,在此感謝 萬分。而一路走來,幫助我的人太多了!國家同步輻射中心的陳俊榮老師,殷程 學長和弘祥學長,清大資工的唐傳義老師以及盈詮,交大生資的PIKI,中研院人 很好的饒博士,清大生科的陳婷學姊,還有熱心的-輔宣同學。真多虧有你們的 幫助,使我的實驗過程中能多元豐富且更順利,再次感謝啦~ LEPE的夥伴們啊,也真多虧有了你們,讓我平淡無奇的碩士班生涯中多增添 了幾分調味。首先要最最感謝的人,就是當初能從pipetman怎麼用開始教的小木 學長。教一個完全沒背景的專題生,這完全就是在考驗您老人家的耐心嘛!但最 讓我受惠的,是小木學長的人生觀與中心思想,從他的身上,我學到了更多書本 上學不到的東西,值得作為人生的準則去追尋。再來就是超厲害的小羊學長(尬酒 尬歌都超強),感覺實驗到他手上就能完全搞定了,雖然我正式踏進來實驗室時他 跟小木學長兩老就雙雙離開了(淚),但對我在專題生時代的訓練,到現在做實驗 的態度與執著上,真的有很深遠的影響!阿後啊,陳平狗,一個當了我六年學長 的學長,也真多虧了他的照顧,真的是亦學長亦友,不過我們的感情絕不是幾句 話就說得完低,在此就介紹完畢。當然,少不了的還有Rich學長啊,三年下來也 吃過他不少的好料啊,不過當然最重要的,除了教我很多人生的道理外,也教了 我很多生物資訊的觀念,超級受用,都可以去開班了啊。還有好人降大勾,一個 我視為最有潛力的研究生,也多虧了有他一路的幫忙,才能讓我的實驗和生活多 iii.
(6) 了幾分味道,很期待看到你的表現啊,加油!還有感謝鄉民團暨煮粥宵夜團的大 晃(鄉長),嘉蔚(鄉民代表兼賢慧組組員),僕僕(麵包超人),淑婷(人超好兼賢慧組 組員)。另外也感謝實驗室的莊博,政哲學長,小米,郁吟學姊(人超好),惠菁學 姊(人超好兼賢慧組組長),好笑又很勁爆的雞姐(原諒我還是喜歡這麼叫你 :)), 人很好的來姐,怡宏學長,很拼很有想法的秉鈞學長,超強的得分王小志(球根本 就是他養的),很有一套的昱樹(兼不是很準的氣象官)。再來就是我視為跟降大勾 一樣厲害的美春(加油阿,期待你的表現!),韋汝,很有喜感的小丸,以及曾經 待過LEPE且很照顧我的小羿,對我很好的斌勾,可愛的摳卡,阿法勾(超強,生 資的魔人兼達人),carol,還有短暫相處過的Quillan,以及優秀的專題生們(未來 實驗室的新血),謝謝大家啦!! 最後感謝我的家人,能永遠在我的背後給我一個最安穩的後盾和避風港。感 謝爸爸和媽媽的栽培及無怨言的支持,沒有你們我決不可能爬上來這研究所的殿 堂。還有哥哥,姐姐,以及小妹的支持,讓我能在求學中無後顧之憂地往前衝! 送給大家我在桌前的座右銘:. 有磨練才能熟練、有投入才能深入、有付出才能傑出! 共勉之~ <( ̄︶ ̄)>~b. 謝謝大家. LE May, 2006. iv.
(7) CONTENTS. PAGE. Abstract (Chinese)................................................................................................................... i Abstract (English) ................................................................................................................... ii Acknowledgement................................................................................................................... iii Contents................................................................................................................................... v Contents of Figures and Tables............................................................................................... vi Abbreviation and Symbol....................................................................................................... ix Chapter 1 Introduction........................................................................................................... 1 Chapter 2 Effects of Quaternary Structure on the Activity of Cytosolic Sulfotransferases.. 19 Chapter 3 Met137 and Tyr238 Modulate Substrate Binding Orientation and Inhibition of Human Dehydroepiandrosterone Sulfotransferase.............................................. 49 Appendix ............................................................................................................................... 90 Autobiography....................................................................................................................... 97. v.
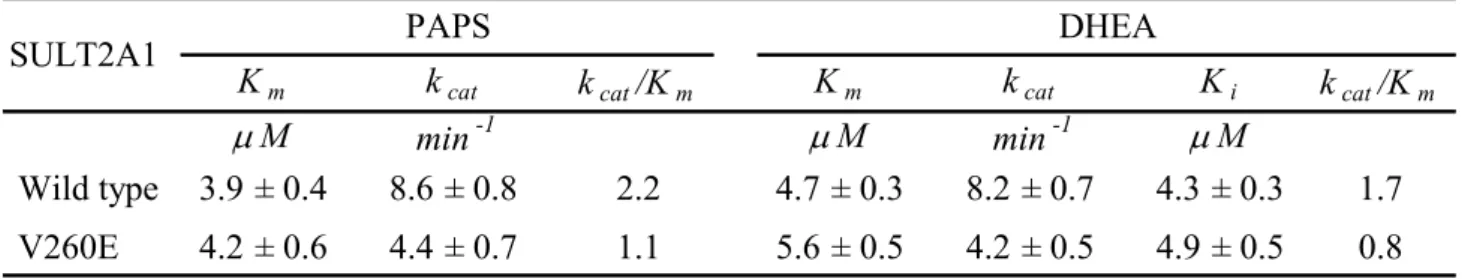
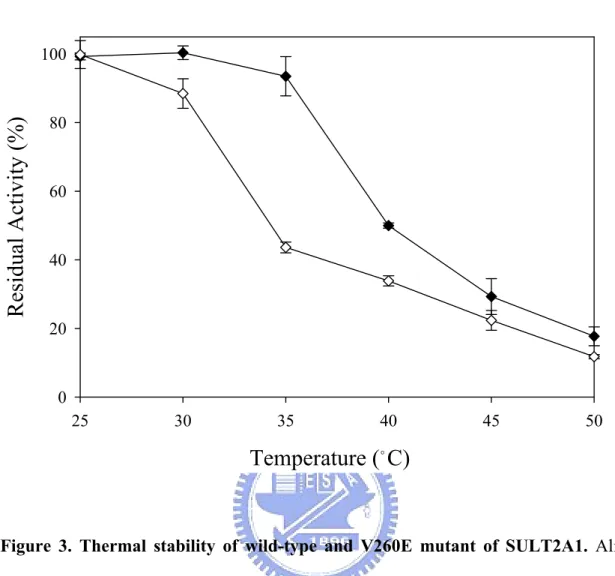
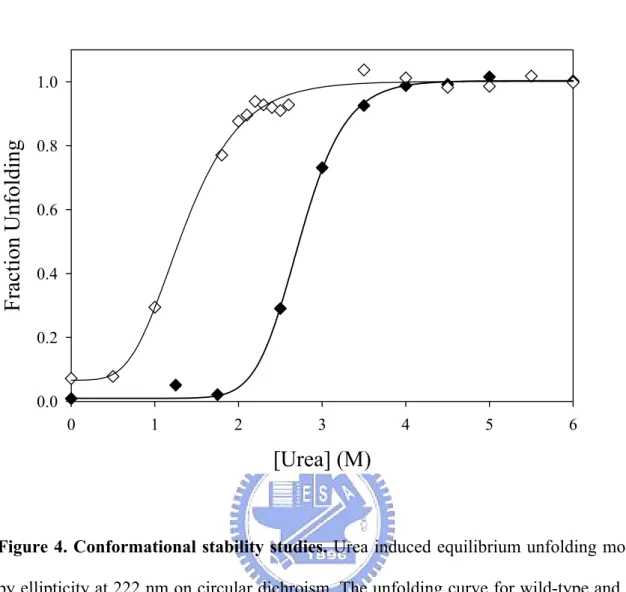
(8) CONTENTS OF TABLES AND FIGURES. PAGE. Chapter 1 Table I. Substrate specificity of human cytosolic sulfotransferases............................... 15. Figure 1. Sulfuryl group transfer in a biological system................................................... 16. Figure 2. Cytosolic sulfotransferase protein classification on the basis of primary amino acid sequence.......................................................................................... 17. Figure 3. Natural level of DHEA with age........................................................................ 18. Chapter 2 Table I. Calibration standard proteins and parameters of gel filtration........................... Table II. Physical and chemical parameters of wild type and monomer mutant of SULT2A1........................................................................................................... 39. 40. Table III Rate constants of PAPS and DHEA with wild-type and V260E mutant of SULT2A1........................................................................................................... 41. Table IV Dissociation constants of PAP and DHEA in wild-type and V260E mutant of SULT2A1.......................................................................................................... Table V. 42. Thermodynamic parameters correlated with free energy of unfolding, △GUNH2O............................................................................................................ 43. Figure 1. Calibration curve of Sephacryl S-200 HR......................................................... 44. Figure 2. Circular dichroism specrtra of wild-type and V260E mutant of SULT2A1...... 45. Figure 3. Thermal stability of wild-type and V260E mutant of SULT2A1...................... 47. Figure 4. Conformational stability studies........................................................................ 48. vi.
(9) Chapter 3 Table I. Primers used for generation of SULT1A1, SULT1A3 and SULT2A1 mutants............................................................................................................... Table II. 72. RMSD values of SULT2A1/DHEA (1J99) and SULT2A1/PAP (1EFH) complex structures............................................................................................. 73. Table III Rate constants of SULT2A1 wild type and mutants using DHEA and ADT as substrates............................................................................................................. 74. Table IV RMSD of SULT2A1/DHEA (1J99) and SULT2A1/ADT (1OV4) complex structures........................................................................................................... Table V.. 75. Dissociation constants of PAP, DHEA, and ADT in wild-type and mutated SULT2A1........................................................................................................... 76. Table VI Rate constants of SULT1A1, SULT1A3 and their mutants............................... 77. Figure 1. Sequence alignment of some cytosolic sulfotransferases.................................. 78. Figure 2. Structures of the loop from residues Tyr-231 to Gly-252 of SULT2A1/PAP, SULT2A1/DHEA and SULT2A1/ADT complexes........................................... 80. Figure 3. Binding modes of DHEA and ADT molecules in SULT2A1............................ 82. Figure 4. Molecular modeling of DHEA and ADT in M137I or M137W of SULT2A1........................................................................................................... 83. Figure 5. Circular dichroism specrtra of wild-type and mutated SULT2A1..................... 84. Figure 6. Multiple sequence and structure alignment of Tyr-238 corresponding residues among some solved-structure sulfotransferases.................................. 86. Figure 7. Schematic illustration of SULT2A1 substrate binding orientation and inhibition modulated by Met-137 and Tyr-238.................................................. 87. Figure 8. SDS-PAGE analysis of M137I and M137W mutants........................................ 88. Figure 9. M137I and M137W mutants cocrystallized with DHEA and ADT................... 89. vii.
(10) Appendix A1.. Primary and secondary structural alignment between SULT1A1 and SULT2A1........................................................................................................... 91. A2.. Tertiary structural alignment between SULT1A1 and SULT2A1...................... 92. A3.. Multiple sequence and secondary structure alignment of partial cytosolic sulfotransferases................................................................................................. viii. 93.
(11) Abbreviation and Symbol. Full Name. ε. Absorption (extinction) coefficient. A280 nm. Absorption at 280 nm. A600 nm. Absorption at 600 nm. ADT. Androsterone. CD. Circular dichroism. DHEA. Dehydroepiandrosterone. DTT. Dithiothreitol. EDC. Ethyl (dimethyl aminopropyl) carbodiimide. k cat. Turnover number. Kd. Dissociation constant. kDa. Kilodalton. Ki. Inhibitory constant of substrate inhibition. Km. Michaelis constant. MALDI-TOF. Matrix-assisted laser desorption ionization time-of-flight. MU. 4-methylumbelliferone. MUS. 4-methylumbelliferyl sulfate. PAPS. 3'-phosphoadenosine 5'-phosphosulfate. PAP. Adenosine 3', 5'-diphosphate. PMSF. Phenylmethylsulfonyl fluoride. p NP. 4-nitrophenol. RP-HPLC. Reversed phase high performance liquid chromatography. SDS. Sodium Dodecyl Sulfate. SULT. Sulfotransferase. V max. Maximum velocity. ix.
(12) CHAPTER 1 Introduction. 1.1 Preface In the post genomic era, great quantities of sequence information are available and most of them are needed to be interpreted for their functional meanings. For example, as the human genomic project is completed in 2003, approximately 20,000-25,000 genes in human DNA were identified and most of them are either unknown or poorly studied. This is following by structural genomics, a field dedicated to a broad understanding of protein structures and functions in relation to gene sequences. The structure of one or more proteins from each family, for a total of about 10000 protein structures in 10 years, will be determined. With these sequence and structure information available, great opportunities have evolved for us to subsequently elucidate the detailed mechanism of enzyme action. Sulfonation is a widespread biological reaction. Sulfotransferase that catalyzes the transfer of a sulfuryl group from 3’-phosphoadenosine 5’-phosphosulfate (PAPS) to a substrate acceptor group is responsible for all the known biological sulfonation. Pathological and toxicological evidences indicate that the abnormal sulfonation leads to inflammations, cancers and infectious diseases (1-3). Understanding the enzyme catalytic mechanism is important for drug design and therapeutic strategy consequently. The use of structure-based sequence alignments and three-dimensional quantitative structure-activity relationship (3D-QSAR) techniques were shown to be very useful to screen for the inhibitors as drug targets in high throughput scale (4-5). In. this. research. we. deduce. the. enzyme. catalytic. mechanism. of. human. dehydroepiandrosterone sulfotransferase (SULT2A1, DHEA-ST) by the comparison of its amino acid sequences and spatially resolved crystal structures. SULT2A1 mainly catalyzes the sulfonation of various steroids and their derivatives, including hydroxysteroids such as. 1.
(13) dehydroepiandrosterone (DHEA), androsterone (ADT), testosterone, estradiol, and many endogenous steroids (6-9). Steroid sulfonation has been recognized as an important process for. maintaining. steroid. hormone. levels. during. their. metabolism.. In. humans,. dehydroepiandrosterone sulfate (DHEAS) is the most prevalent steroid precursor, and is one of the major secretory products of both adult and fetal adrenals (10). To know more about the actions of SULT2A1 to convert the DHEA to DHEAS, this research from sequence and structure of SULT2A1 is studied and then the deduced enzyme mechanism is further confirmed by site-directed mutagenesis and enzymatic analysis.. 2.
(14) 1.2 Sulfotransferase 1.2.1 Sulfonation It has been known for a long time that sulfonation occurs in a biological system. Baumann first described sulfonate conjugation as a pathway in biotransformation in 1876 (11). However, the biochemistry of sulfotransferase enzyme has not been well characterized until the past three decades. Furthermore, the application of molecular biology technology had made it possible to determine the characteristics of these enzymes, whose substrate specificities, regulation, and evolutionary relationships (12). However, the mechanism by which sulfonation takes place was not examined until after the isolation of the active sulfate donor, 3’-phosphoadenosine 5’-phosphosulfate (PAPS), about 80 years later (13). Sulfotransferases. (SULTs). catalyze. the. transfer. of. a. sulfuryl. group. from. 3’-phosphoadenosine 5’-phosphosulfate (PAPS), the universal sulfonate donor molecule, to a variety of amine and hydroxyl substrates as nucleophiles in a process originally called sulfonation (Fig. 1). There are two classes of sulfotransferases: cytosolic sulfotransferases and membrane-associated. sulfotransferases.. Cytosolic. sulfotransferases. sulfonate. small. endogenous and exogenous compounds, such as drugs, steroid hormones, chemical carcinogens, bile acids, and neurotransmitters (14-19). Membrane-associated sulfotransferases, many of which have been implicated recently in crucial biological processes, sulfonate larger biomolecules, suchas carbohydrates and proteins (20-22). Because of the biological importance and medical relevance of sulfotransferases, there is intense interest in the exact functions of these enzymes. Much information is available with regard to the structure, substrate specificity, and kinetic mechanism of cytosolic sulfotransferases, but less information is available on the transition-state structure and design of potent and specific inhibitors. The membrane-associated enzymes are not as well understood, particularly because their important roles in biological processes have only. 3.
(15) recently been uncovered.. 1.2.2 Classification of sulfotransferases The membrane-associated sulfotransferaes are still without a consistent, universal nomenclature scheme and they are named according to their substrate specificity so far. This nomenclature is clarified as required. On the side of cytosolic sulfotransferases, they had often been named after their substrates. However, since the substrate specificities of different sulfotransferases are overlapping, such names can be misleading. To date 10 or 11 human cytosolic sulfotransferases have been characterized and so far the new nomenclature guidelines were applied to 65 sulfotransferase cDNAs and 18 sulfotransferase genes that were characterized from eukaryotic organisms (23). These sequences were evaluated and named on the basis of encoded amino acid sequence identity. Family members share at least 45% amino acid sequence identity whereas subfamily members are at least 60% identical. cDNAs which encode amino acid sequences of at least 97% identity to each other were assigned identical isoform names (Fig. 2).. 1.2.3 Substrate specificity of sulfotransferases Although the sulfotransferase mainly possess PAPS as the universal sulfonate donor molecule, a number of studies have also been conducted on the specificity of the PAPS-binding site (24-25). Moreover, different sulfotransferases exhibit distinct substrate specificity. For example, sulfonation of carbohydrates, peptides, hormones, steroids and neurotransmitters are all catalyzed by different sulfotransferases. However, within a subfamily, sulfotransferase has a broad substrate spectrum for similar substrates. As can be seen in Table I, the diverse substrate specificity is catalyzed by different sulfotransferases which own oneself preferred substrate. For example, in SULT1 family, the preferred. 4.
(16) substrates of sulfotransferase are simple-phenol compounds, such as p-nitrophenol (pNP), dopamine, and thyroid. Sulfotransferase in SULT2 family, however, mainly catalyzes hydroxysteroids, such as cholesterol, androsterone, dehydroepiandrosterone, pregnenolone, and estradiol. Each of the two families, SULT1A1 and SULT2A1, contains diverse substrate binding site and a separate but identical PAPS binding site.. 1.2.4 Structure of cytosolic sulfotransferase Sulfotransferases are a single α/β globular protein with a characteristic five-stranded parallel β-sheet and α-helices flank both sides of the sheet (26-27). Sulfotransferases share a similar structural resemblance to the nucleotide kinases with their secondary structures conserved not only in position but also in connectivity (28). The strand–loop–helix and strand–turn–helix motifs constitute the core PAPS binding site, providing the majority of the enzyme interactions with the PAP molecule. The PSB-loop interacts with the 5’-phosphate of the PAP molecule, whereas helix 6 of the strand–turn– helix unit that runs parallel to the PSB-loop provides interaction with the 3’-phosphate. Not only the structures, but also the amino acid sequences of the phosphate-binding sites are conserved in all sulfotransferases including both the cytosolic and membrane enzymes. (29). Furthermore, although the sulfotransferase enzymes display broad substrate specificity, a given enzyme can often be characterized by a specific substrate. The underlying principle that regulates the characteristic substrate specificity is not well developed. Finally, cytosolic sulfotransferases are generally homodimers in their catalytically active forms in solution. The conserved dimerization motif is found in the cytosolic enzymes (30). Multiple amino acid sequence alignments show that the motif consists of ten residues near the C-terminus and is represented by the consensus sequence KXXXTVXXXE. Structural comparisons and mutagenesis studies were undertaken with mouse SUL T1E1 (a monomer) and human. 5.
(17) SULT1E1 (a dimer) in an attempt to identify a common structural motif. It was found that the mutations V269E and V260E converted the homodimers SULT1E1 and SULT2A1, respectively, into monomers.. 6.
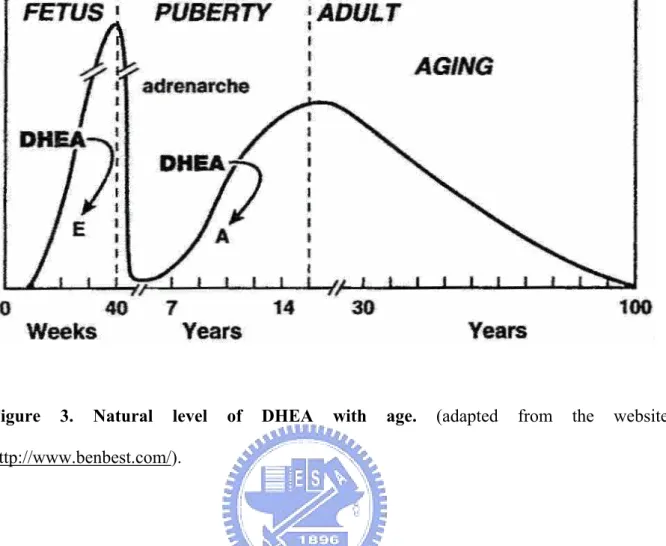
(18) 1.3 Dehydroepiandrosterone (DHEA) and dehydroepiandrosterone 3β-sulfate (DHEAS) 1.3.1 Background Hormones play a plenty of functions physiologically and the abnormal regulation may force the dysfunction and raise the risk, such as the hormone homeostasis and hormone-related disease or metabolism. In this study, I mainly interested in the study of sexual hormone, such as the estradiol, androsterone, testosterone, and dehydroepisterone. For example, the dehydroepisterone (DHEA) plays the profound role and is the precursor of the sexual hormone.The abnormal regulation may cause the irregular secretory amounts of the other hormone as described above. Dehydroepiandrosterone (DHEA) was isolated in urine in 1934, and DHEA 3β-sulfate (DHEAS) was identified 10 years later (31-32). It took another decade to identify DHEA and DHEAS in peripheral blood (33-34). DHEA is one of the hormones produced by the adrenal glands. After being secreted by the adrenal glands, it circulates in the bloodstream as DHEAS and is converted as needed into other hormones. DHEA and DHEAS are the most abundant steroid hormones in the human bloodstream. DHEA is known to be a precursor to the numerous steroid sex hormones (including estrogen and testosterone), which serve well-known functions. While DHEA levels reach their peak in the early morning hours, DHEAS levels show no diurnal variation. It is also one of the most significant age-related biomarkers, which predictably declines with age in even the healthiest of people. Blood levels are highest in the developing fetus, drop sharply after birth, begin climbing again at age 6 to 8 (a time of rapid growth) to a peak at age 25 to 30 and then decline to about 10% of the peak level by age 80 (Fig. 3). Abnormally low levels of DHEA have been reported to be related to a number of diseases, including cancer, diabetes, coronary artery disease, and obesity.. 7.
(19) 1.3.2 Physiological regulation Little is known about how DHEA works in the body and it often has different effects in men, premenopausal women, and postmenopausal women (35-36). Supplementation with DHEA-S has resulted in increased levels of testosterone and androstenedione (37). The conversion of DHEA into testosterone may account for the fact that low blood levels of DHEA have been reported in some men with erectile dysfunction. The findings of a double-blind trial using 50 mg supplements of DHEA taken daily for six months suggests that DHEA may improve erectile function in some men (38).. 1.3.3 DHEA modulates immunity A group of elderly men with low DHEA levels who were given 50 mg of DHEA per day for 20 weeks experienced a significant activation of immune function (39). Postmenopausal women have also shown increased immune functioning in just three weeks when given DHEA in double-blind research (40). DHEA also regulates the systemic lupus erythematosus (SLE), an autoimmune disease. It has been linked to abnormalities in sex hormone metabolism (41). Supplementation with very large amounts of DHEA (200 mg per day) improved clinical status and reduced the number of exacerbations of SLE in a double-blind trial (42). A preliminary trial has confirmed the benefit of 50-200 mg per day of DHEA for people with SLE (43).. 1.3.4 DHEA and cardiopathy Some reports have suggested that DHEA might reduce the risk of heart disease, perhaps by lowering cholesterol levels. In fact, higher levels of DHEA and DHEAS have been associated with cardiovascular risk factors in women, including high blood pressure and smoking (44). Moreover, DHEA has also been reported to lower HDL (45).. 8.
(20) 1.4 Conculsion DHEA is a multifunctional hormone and plays a plenty of function, such as steroid metabolism, cancer, immunity, and aging or more. The sulfated form of DHEA, DHEAS, is primarily in the adrenals, the liver, and small intestines. However, in blood, most DHEA is found as DHEAS with levels that are about 300 times higher than free DHEA. Orally ingested DHEA is converted to its sulfate when passing through intestines and liver. DHEAS is biologically active only after its sulfate group has been split and it becomes DHEA again. From a practical point measurement of DHEAS is preferable to DHEA as levels are more stable. The conversion between DHEA and DHEAS is regulated by SULT2A1 which preferred substrate is DHEA and the regulation of this enzyme and catalytic mechanism is still far from known. In this study we deduce the enzyme mechanism and function from its amino acid sequence and the solved crystal structure. Herein we analyze the quaternary structure, wild-type homodimer and monomer mutant. The monomer mutant, V260E, sufficiently interfere the hydrophobic dimerization interface and the molecular weight is confirmed by gel filtration. Further the circular dichroism spectrum, enzyme kinetics, substrate binding affinity, and the thermal and conformational stability reveal the difference between dimer and monomer of SULT2A1. Moreover, the comparisons of SULT2A1 structure complex also facilitate us to infer the substrate inhibition mechanism and further identify the regulatory amino acid residues. Experimental results and the multiple structural alignment all indicate that the involvement of this amino acid residue in substrate inhibition may be a general feature in all cytosolic sulfotransferases.. 9.
(21) REFERENCES. 1. Hemmerich, S., Verdugo, D., and Rath, V. L. (2004) Strategies for drug discovery by targeting sulfation pathways. Drug Discov. Today 9, 967–975 2. Kauffman, F. C. (2004) Sulfonation in pharmacology and toxicology. Drug Metab. Rev. 36, 823–843 3. Pasqualini, J. R. (2004) The selective estrogen enzyme modulators in breast cancer: a review. Biochim. Biophys. Acta. 1654, 123–143 4. Sharma, V., and Duffel, M. W. (2005) A comparative molecular field analysis-based approach to prediction of sulfotransferase catalytic specificity. Methods Enzymol. 400, 249–263 5. Rath, V. L., Verdugo, D., and Hemmerich, S. (2004) Sulfotransferase structural biology and inhibitor discovery. Drug Discov. Today. 9,1003–1011 6. Falany, C. N. (1997) Enzymology of human cytosolic sulfotransferases. FASEB J. 11, 206–216 7. Weinshilboum, R. M., Otterness, D. M., Aksoy, I. A., Wood, T. C., Her, C., and Raftogianis, R. B. (1997) Sulfation and sulfotransferases 1: sulfotransferase molecular biology: cDNAs and genes. FASEB J. 11, 3–14 8. Chen, G., Banoglu, E., and Duffel, M. W. (1996) Influence of substrate structure on the catalytic efficiency of hydroxysteroid sulfotransferase STa in the sulfation of alcohols. Chem. Res. Toxicol. 9, 67–74 9. Kakuta, Y., Pedersen, L. C., Chae, K., Song, W. C., Leblanc, D., London, R., Carter, C. W., and Negishi, M. (1998) Mouse steroid sulfotransferases: substrate specificity and preliminary X-ray crystallographic analysis. Biochem. Pharmacol. 55, 313–317 10. Chang, H. J., Shi, R., Rehse, P., and Lin, S. X. (2004) Identifying androsterone (ADT) as. 10.
(22) a cognate substrate for human dehydroepiandrosterone sulfotransferase (DHEA-ST) important for steroid homeostasis: structure of the enzyme-ADT complex. J. Biol. Chem. 279, 2689–2696 11. Baumann, E. (1876) Ber. Dtsch. Chem. Ges. pp. 54–58 12. Weinshilboum, R. M., Otterness, D. M., Aksoy, I. A., Wood, T. C., Her, C., and Raftogianis, R. B. (1997) Sulfation and sulfotransferases 1: sulfotransferase molecular biology: cDNAs and genes. FASEB J. 11, 3–14 13. Robbins, P. W., and Lipmann, F. (1956) Identification of enzymatically active sulfate as adenosine-3’-phosphate-5’-phosphatesulfate. J. Am. Chem. Soc. 78, 2652–2653 14. Mulder, G. J., and Jakoby, W. B. (1990) in Conjugation Reactions in Drug Metabolism (Mulder, G. J., ed) pp. 107–161, Taylor and Francis, Ltd., London 15. Falany, C., and Roth, J. A. (1993) in Human Drug Metabolism: From Molecular Biology to Man (Jeffery, E. H., ed) pp. 101–115, CRC Press, Inc. Boca Raton, FL 16. Weinshilboum, R., and Otterness, D. (1994) in Conjugation-Deconjugation Reactions in Drug Metabolism and Toxicity (Kaufmann, F. C., ed) pp. 45–78, Springer-Verlag, Berlin 17. DeBaun, J. R., Miller, E. C., and Miller, J. A. (1970) N-hydroxy-2-acetylaminofluorene sulfotransferase: its probable role in carcinogenesis and in protein-(methion-S-yl) binding in rat liver. Cancer Res. 30, 577–595 18. Glatt, H. (1997) Sulfation and sulfotransferases 4: bioactivation of mutagens via sulfation. FASEB J. 11, 314–321 19. Falany, C. N. (1997) Sulfation and sulfotransferases. Introduction: changing view of sulfation and the cytosolic sulfotransferases. FASEB J. 11, 1–2 20. Habuchi, O. (2000) Diversity and functions of glycosaminoglycan sulfotransferases. Biochem. Biophys. Acta 1474, 115–127 21. Grunwell, J. R., and Bertozzi, C. R. (2002) Carbohydrate sulfotransferases of the. 11.
(23) GalNAc/Gal/GlcNAc6ST family. Biochemistry 41, 13117–13126 22. Kehoe, J. W, and Bertozzi, C. R. (2000) Tyrosine sulfation: a modulator of extracellular protein-protein interactions. Chem. Biol. 7, 57–61 23. Blanchard, R. L., Freimuth, R. R., Buck, J., Weinshilboum, R. M., and Coughtrie, M. W. (2004) A proposed nomenclature system for the cytosolic sulfotransferase (SULT) superfamily. Pharmacogenetics 14, 199–211 24. Hsiao, Y. S., and Yang, Y. S. (2002) A single mutation converts the nucleotide specificity of phenol sulfotransferase from PAP to AMP. Biochemistry 41, 12959–12966 25. Lin, E. S., and Yang, Y. S. (2000) Nucleotide binding and sulfation catalyzed by phenol sulfotransferase. Biochem. Biophys. Res. Commun. 271, 818–822 26. Kakuta, Y., Pedersen, L. G., Carter, C. W., Negishi, M., and Pedersen, L. C. (1997) Crystal structure of estrogen sulphotransferase. Nat. Struct. Biol. 4, 904–908 27. Negishi, M., Pedersen, L. G., Petrotchenko, E., Shevtsov, S., Gorokhov, A., Kakuta, Y., and Pedersen, L. C. (2001) Structure and function of sulfotransferases. Arch. Biochem. Biophys. 390, 149–157 28. Bidwell, L. M., McManus, M. E., Gaedigk, A., Kakuta, Y., Negishi, M., Pedersen, L., and Martin, J. L. (1999) Crystal structure of human catecholamine sulfotransferase. J. Mol. Biol. 293, 521-530 29. Dajani, R., Cleasby, A., Neu, M., Wonacott, A. J., Jhoti, H., Hood, A. M., Modi, S., Hersey, A., Taskinen, J., Cooke, R. M., Manchee, G. R., and Coughtrie, M. W. (1999) X-ray crystal structure of human dopamine sulfotransferase, SULT1A3. Molecular modeling and quantitative structure-activity relationship analysis demonstrate a molecular basis for sulfotransferase substrate specificity. J. Biol. Chem. 274, 37862-37868 30. Petrotchenko, V., Pedersen, L. C., Borches, C. H., Tomer, K. B., and Negishi, M. (2001) The dimerization motif of cytosolic sulfotransferases. FEBS Lett. 490, 39–43. 12.
(24) 31. Butemandt, A., and Dannenbaum, H. (1934) Z. Physiol. Chem. 229, 192–195 32. Munson, P. L., Gallagher, T. F., and Koch, F. C. (1944) J. Biol. Chem. 152, 67–77 33. Baulieu, E. E. (1960) Three sulfate esters of 17-ketosteroids in the plasma of normal subjects and after administration of ACTH. J. Clin. Endocrinol. Metab. 20, 900–904 34. Migeon,. C.. J.,. and. Plager,. J.. E.. (1954). Identification. and. isolation. of. dehydroisoandrosterone from peripheral human plasma. J. Biol. Chem. 209, 767–772 35. Weksler, M. E. (1996) Hormone replacement for men. Br. Med. J. 312, 859–860 36. Ebeling,. P.,. and. Koivisto,. V.. A.. (1994). Physiological. importance. of. dehydroepiandrosterone. Lancet 343, 1479–1481 37. Stomati, M., Rubino, S., Spinetti, A., Parrini, D., Luisi, S., Casarosa, E., Petraglia, F., and Genazzani, A. R. (1999) Endocrine, neuroendocrine and behavioral effects of oral dehydroepiandrosterone sulfate supplementation in postmenopausal women. Gynecol. Endocrinol. 13, 15–25 38. Reiter, W. J., Pycha, A., Schatzl, G., Pokorny, A., Gruber, D. M., Huber, J. C., and Marberger, M. (1999) Dehydroepiandrosterone in the treatment of erectile dysfunction: a prospective, double-blind, randomized, placebo-controlled study. Urology 53, 590–595 39. Khorram, O., Vu, L., and Yen, S. S. (1997) Activation of immune function by dehydroepiandrosterone (DHEA) in age-advanced men. J. Gerontol. A Biol. Sci. Med. Sci. 52, M1–7 40. Casson, P. R., Andersen, R. N., Herrod, H. G., Stentz, F. B., Straughn, A. B., Abraham, G. E., and Buster, J. E. (1993) Oral dehydroepiandrosterone in physiologic doses modulates immune function in postmenopausal women. Am. J. Obstet. Gynecol. 169, 1536–1539 41. Lahita, R. G., Bradlow, H. L., Ginzler, E., Pang, S., and New, M. (1987) Low plasma androgens in women with systemic lupus erythematosus. Arthritis. Rheum. 30, 241–248 42. van. Vollenhoven,. R.. F.,. Engleman,. 13. E.. G.,. and. McGuire,. J.. L.. (1995).
(25) Dehydroepiandrosterone in systemic lupus erythematosus. Results of a double-blind, placebo-controlled, randomized clinical trial. Arthritis. Rheum. 38, 1826–1831 43. van Vollenhoven, R. F., Morabito, L. M., Engleman, E. G., and McGuire, J. L. (1998) Treatment of systemic lupus erythematosus with dehydroepiandrosterone: 50 patients treated up to 12 months. J. Rheumatol. 25, 285–289 44. Johannes, C. B., Stellato, R. K., Feldman, H. A., Longcope, C., and McKinlay, J. B. (1999) Relation. of. dehydroepiandrosterone. and. dehydroepiandrosterone. sulfate. with. cardiovascular disease risk factors in women: longitudinal results from the Massachusetts Women's Health Study. J. Clin. Epidemiol. 52, 95–103 45. Mortola, J. F., and Yen, S. S. (1990) The effects of oral dehydroepiandrosterone on endocrine-metabolic parameters in postmenopausal women. J. Clin. Endocrinol. Metab. 71, 696–704. 14.
(26) Table 1: Substrate specificity of human cytosolic sulfotransferasesa.. SULT. Representative. pNP 2-napthol dopamine 1A1 tyramine minoxidol 3,3'-T2 T3 1A2 1A3. 1B1. 1C1 1C2. 1E1. 2A1. 2B1a. 2B1b. a. pNP dopamine pNP tyramine dopamine pNP T4 T3 rT3 3,3'-T2 pNP dopamine N-OH-2AAF pNP dopamine N-OH-2AAF estradiol DHEA androstenediol pNP estradiol DHEA androstenediol cholesterol DHEA pregnenolone 17α− DHEA pregnenolone 17α-hydroxypregnenolone cholesterol. Km(PAPS) (μM) Km (μM) 1.1 200 130 1.0 6483 2800 0.14 29.1 69(HAST-4) 5.5(HAST-4) 4.1(HAST-4v) 0.2(HAST-4v) ND 1024 1.1 35 3.7 5 23 1.2 ND 141 51 84 8.8 ND 28.6 75.3 ND 47.5 0.025 4.6 0.3 3.2 76.6 2.0 1.0 1.6 0.2 2.3 0.2 0.9 4.4 0.6 1.0. 0.1. 0.1. Adapted from Chapman et al. (2004) Angew. Chem. Int. Ed. Engl. 43, 3526-3548. 15.
(27) NH2 N N. O O -O S O P O OO. XH=OH, NH2, SH, NOH. N N. O O. RXH. OH. -O P O ONH2 N N. O -O P O O-. N N. O RX S OO. O O. OH. -O P O O-. Figure 1. Sulfuryl group transfer in a biological system.. 16.
(28) Figure 2. Cytosolic sulfotransferase protein classification on the basis of primary amino acid sequence. The cladogram depicts the evolutionary relationship of sulfotransferase enzyme superfamily members based on their amino acid sequences. (Adapted from Blanchard et al. (2004) Pharmacogenetics. 14, 199-211).. 17.
(29) Figure. 3.. Natural. level. of. DHEA. with. http://www.benbest.com/).. 18. age.. (adapted. from. the. website.
(30) CHAPTER 2 Effects of Quaternary Structure on the Activity of Human Dehydroepiandrosterone Sulfotransferase. Human dehydroepiandrosterone sulfotransferase (SULT2A1, DHEA-ST) catalyzes the transfer of the sulfuryl group of 3’-phosphoadenosine 5’-phosphosulfate (PAPS) to hydroxysteroids such as dehydroepiandrosterone (DHEA). Most cytosolic sulfotransferases are generally known to be homodimer in solution and the specific amino acid residues responsible for the dimerization have been identified as the KXXXTVXXXE motif (Petrotchenko et al. (2001) FEBS Lett. 490, 39–43). However, the comparisons of the characteristics. and. catalytic. actions. between. dimer. and. monomer. of. cytosolic. sulfotransferases have not been studied in detail so far. To study the effects of quaternary structure on the activity of the enzyme, monomer mutant of SULT2A1 was prepared through site-directed mutagenesis. The mutant V260E of SULT2A1 was sufficient to convert to the monomer by interfering with hydrophobic KTVE motif and confirmed by gel filtration. The circular dichroism spectrum of dimer and monomer mutant reveals the slight conformational change whether in the secondary or tertiary structure. The kinetic constants of dimer and monomer mutant are quite similar whether the PAPS or DHEA are examined, however, the catalytic efficiency, kcat/Km, shows two folds higher in dimer than that in monomer. It indicates that the two subunits in dimer may be both catalytically competent. The binding affinity shows no significantly change whether for PAP in the binary complex (SULT2A1/PAP) or DHEA in the tertiary complex (SULT2A1/PAP/DHEA) between dimer and monomer. Furthermore, the thermal and conformational stability between dimer and monomer mutant all revealed the monomer is more labile than dimer. The effect of the quaternary structure on SULT2A1 may be a model to deduce the other cytosolic sulfotransferases whether the wild type is the homodimer or monomer in solution naturally.. 19.
(31) INTRODUCTION. Sulfotransferases (SULTs) are a large family of enzymes that catalyze the transfer of sulfuryl group from the common 3’-phosphoadenosine 5’-phosphosulfate (PAPS) to numerous endogenous and exogenous compounds. Cytosolic sulfotransferases sulfonate small molecules such as drugs, steroid hormones, chemical carcinogens, bile acids, and neurotransmitters (1-6). Human dehydroepiandrosterone sulfotransferase (SULT2A1, DHEA-ST) catalyzes the sulfonation of various steroids and their derivatives, including hydroxysteroids such as dehydroepiandrosterone (DHEA), androsterone (ADT), testosterone, estradiol, and many endogenous steroids (7-10). Steroid sulfonation has been recognized as an important process for. maintaining. steroid. hormone. levels. during. their. metabolism.. In. humans,. dehydroepiandrosterone sulfate (DHEAS) is the most prevalent steroid precursor, and is one of the major secretory products of both adult and fetal adrenals (11). Cytosolic sulfotransferases consist of around 300 amino acid residues, and most of them are found as homodimer in solution. Previous study (12) performed the cross-linking by ethyl (dimethyl aminopropyl) carbodiimide (EDC) following the separation by C4 reversed phase high performance liquid chromatography (RP-HPLC) column and analyzed by matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF). It was found that the cross-linked peptides were located between one subunit to the other subunit both nearby the carboxyl terminus, and they also exposured on the surface of each subunit. Therefore the mutants V269E and V260E in human SULT1E1 and SULT2A1, respectively, converted the homodimers into monomers. It was further shown that the mutant P269T and E270V performed simultaneously in mouse SULT1E1, a monomer naturally in solution, forced the formation of the dimer. It was concluded that the general motif KXXXTVXXXE (KTVE. 20.
(32) motif) is responsible for the dimerization of cytosolic sulfotransferases. It had also been reported that sulfotransferases are capable of forming not only homodimers but also the heterodimers (13). Moreover, the crystal structures of numerous cytosolic sulfotransferases have recently been solved and they all contain diverse substrate binding site and a separate but identical PAPS binding site (14-19). To date, despite the dimerization motif of cytosolic sulfotransferases had been identified (12), the effect of quaternary structure on the catalysis and function of cytosolic sulfotransferases was still far from known and the protein-protein interaction of cytosolic sulfotransferases has not been well studied. The functional significance of the dimerization process is not known at the present time. In this report, we performed that the mutant V260E of SULT2A1 was sufficient to convert the dimer to a monomer by interfering the hydrophobic KTVE motif and confirmed by gel filtration. To investigate the effects of the quaternary structure on the catalysis and function, the studies of the conformational change, catalytic kinetics, substrate inhibition, substrate binding, thermal and conformational stability are all examined. Furthermore the differences between dimer and monomer of SULT2A1 will be clarified and this may be a model to study the effect of quaternary structure among cytosolic sulfotransferases.. 21.
(33) EXPERIMENTAL PROCEDURES. Materials—PfuTurbo DNA polymerase was purchased from Stratagene, and MUS, MU, PAP, PAPS, dehydroepiandrosterone (DHEA), glutathione (reduced form), PAP agarose and thrombin were purchased from Sigma (St. Louis, MO). Potassium phosphate (dibasic) was obtained from J. T. Baker (U.S.A.). DEAE Sepharose fast flow, Glutathione S-transferase Sepharose fast flow, Sephacryl S-200 HR, and low molecular weight gel filtration calibration kit were obtained from Amersham Pharmacia Biotech Asia Pacific (Hong Kong). All other chemicals were obtained commercially at the highest purity possible. Site-Directed Mutagenesis—The QuikChange site-directed mutagenesis kit from Stratagene was used for the generation of monomer mutant of SULT2A1. Briefly, wild-type SULT2A1 cDNA incorporated in pGEX-2TK prokaryotic expression vector was used as the template in conjunction with specific mutagenic primers. The monomer sense mutagenic oligonucleotide primer sets was 5’-tcacttcacagaggcccaagctg-3’ for SULT2A1. The amplification conditions were 30 cycles of 30 sec at 95 °C, 1 min at 55 °C, and 7 min at 68 °C. The reaction contained 100 ng of DNA template, 0.4 μM of sense and antisense primers, 200 μM of each of the four deoxynucleotide triphosphates (dNTPs), 1×PfuTurbo buffer, and 2.5 units of PfuTurbo DNA polymerase. The PCR products were treated with DpnI restriction enzyme for 1 hour at 37 °C followed by transforming into DH5α. Mutated cDNA sequences were confirmed using an ABI Prism 377 DNA sequencer (Applied Biosystems, Foster City, CA) following the standard protocol. Expression, Purification and Characterization of Wild-type and Monomer Mutant of SULT2A1—The expression and purification of SULT2A1 was described previously (20-21). Competent E. coli BL21 (DE3) cells transformed with pGEX-2TK vector harboring the wild-type or V260E point-mutated SULT2A1 cDNA were grown to A600 nm = 0.8 in 0.5 liter. 22.
(34) of LB medium supplemented with 50 μg/ml ampicillin. After induction with 1 mM isopropyl β-D-thiogalactopyranoside overnight at 25 ℃, the cells were collected by centrifugation and homogenized in an ice-cold lysis buffer containing 20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 125 mM sucrose, and 10% glycerol using a Fisher Scientific Sonic Dismenbrator. The supernatant collected was fractionated by the glutathione sepharose and the bound fusion protein was treated with 15 units of bovine thrombin for 8 hours at 4 ℃. A homogeneous protein was obtained as determined by SDS-polyacrylamide gel electrophoresis (22), Protein concentration of the homogeneous form of wild type and monomer mutant SULT2A1 were estimated on the basis of absorbency at 280 nm (2.38 ml/mg cm-1) (23) using a UV/Vis spectrophotometer (Hitachi UV/Vis-3300, Japan). Size-Exclusion Chromatography and Calibration Curve Preparation—Aliquots of 1ml of various samples was applied on a Sephacryl S-200 HR column that was equilibrated with 20 mM Tris buffer pH 7.5 containing 0.15 M NaCl. Proteins were eluted with the same buffer at a flow rate of 1.0 ml/min. Molecular weights were estimated against calibration standards consisted of ribonuclease A (15.6 kDa), chymotrypsinogen A (19.4 kDa), ovalbumin (47.6 kDa), and albumin (62.9 kDa). Molecular weights of wild type (homodimer) and monomer mutant of SULT2A1 were estimated by gel filtration chromatography (12). Circular Dichroism Measurement—CD measurements were carried out on a Jasco J715 spectropolarimeter and processed data with J-700 standard analysis version 1.33.00 (Jasco, Japan). CD spectra of enzymes at a concentration of 1 mg/ml in 20 mM phosphate buffer (pH 7.0) were measured in a 1 cm light path length quartz cell in the near-UV range (240-320 nm) and 1 mm light path in the far-UV (200-240 nm) at room temperature. Enzyme Assay—The activities of wild-type and monomer mutant of SULT2A1 were determined according to the change of fluorescence based on a coupled-enzyme assay method (24). The fluorescence of MU at 460 nm was measured upon excitation at 355 nm. The. 23.
(35) reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate buffer at pH 7.0, 5 mM 2-mercaptoethanol, 20 μM PAPS, 2 mM MUS, 5.4 μg K65ER68G (25) of rat SULT1A1, SULT2A1 and 5 μM of DHEA at 37 ℃. A linear response was obtained when 1.49 to 14.9 nM (0.1 to 1 μg) SULT2A1 was added in the standard assay condition (24). In the reaction condition, regeneration of PAPS catalyzed by K65ER68G of rat SULT1A1 is much faster than the production of PAP catalyzed by SULT2A1 for approximately 40 to 50 folds. Substrate Binding—The dissociation constants (Kd) of PAP and DHEA towards wild-type and monomer mutant of SULT2A1, respectively, were determined with a spectrofluorimeter (26). The decrease in intrinsic fluorescence of protein at 340 nm was observed upon excitation at 280 nm when an aliquot amount of PAP was added into the mixture consisted of 100 mM potassium phosphate buffer at pH 7.0, 100 nM wild-type or monomer mutant of SULT2A1 and varying concentrations of PAP at 25 ℃ with a final volume of 1.3 ml in a quartz cuvette of 1 cm square cross-section. In the ternary experiments, DHEA was added into the pre-incubated solution containing 0.5 μM wild type or monomer mutant of SULT2A1, 100 mM potassium phosphate at pH 7.0, and 1 μM PAP at 25 °C. Each data point was duplicated, and the difference was within 10%. Thermal Stability—To examine the thermal stability of the wild type and monomer mutant of SULT2A1, aliquots of the protein were treated for 15 minutes at desired temperatures as follows: 25, 30, 35, 40, 45 and 50 °C individually and then were performed by coupled-enzyme assay as described above. Urea-Dependent Unfolding Experiments—For each data point in the urea denaturation experiment, 5 μM wild type or monomer mutant of SULT2A1 in 20 mM potassium phosphate buffer at pH 7.0 and 10% glycerol was treated at desired concentration of urea from 0 to 6 M at 4 ℃ for approximately 24 hours (27-29). Measurements of circular dichroism at 222 nm 24.
(36) of urea-treated enzymes were made in a Jasco J715 spectropolarimeter at room temperature with a cell of 1 mm path length as described above. CD222nm values were converted into fraction of unfolded values to facilitate comparison of denaturation curves. Kinetic Data Analysis—Kinetic experiments were analyzed using nonlinear regression to fit the appropriate equation to the data. In non-inhibitory experiments were individually fit to Michaelis-Menten Equation 1, and substrate inhibition data were fitted to substrate inhibition in Equation 2 (30). The kinetic constants (Km, kcat, and Ki) were obtained from SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL). Data shown represent mean values derived from two determinations. v = V[S]/(Km + [S]). (Eq. 1). v = V[S]/{Km + [S](1 + [S]/Ki)}. (Eq. 2). Analysis of Urea Denaturation Data—This analysis is for a two-state model of denaturation where only the native and the denatured states are populated and it has been found experimentally that the free energy of unfolding of proteins in the presence of denaturant is linearly related to the concentration (27, 31-32). The CD222nm signal, F, plotted as a function of the urea concentration, [Urea], is given by Equation 3, where α is the intercept and β is the slope of the baseline at the native (N) or unfolding (U) state. [Urea]50% is the urea concentration at the midpoint of urea denaturation, and m value is a constant that is proportional to the increase in the degree of exposure of the protein on denaturation. R is the gas constant, 8.314 J mol-1 K-1, and T is the temperature in K. The parameters △GUNH2O , the free energy of denaturation in the absence of urea and is the product of [Urea]50% and m, and m value were obtained by nonlinear regression fitting of the data to Equation 3 with SigmaPlot 2001, V7.0 and Enzyme Kinetics Module, V1.1 (SPSS Inc., Chicago, IL). The [Urea]50% is obtained by Equation 4.. 25.
(37) F=. {(α N + β N [Urea]) + [(α U + β U [Urea]) × exp[(m[Urea] - ΔG UN. ΔG UN. {1 + exp[(m[Urea] - ΔG UN H 2O. H 2O. = m[Urea]50%. )/RT]}. H 2O. )/RT]}. (Eq. 3). (Eq. 4). 26.
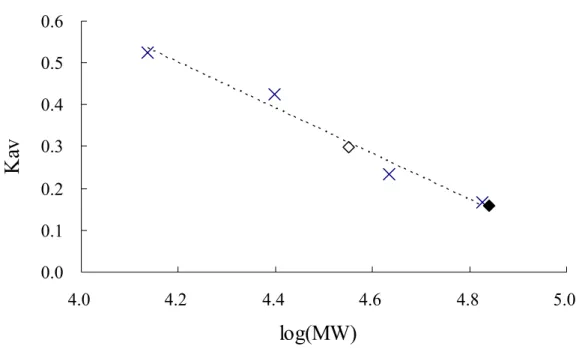
(38) RESULTS. Enzyme Molecular Weight Estimation—The previous study (12) had shown that the dimerization interface of cytosolic sulfotransferases naturally in solution exists and the subunits form the dimer through the highly conserved sequence KXXXTVXXXE nearby the carboxyl terminus. The KTVE motif of each subunit appears to form a zipper-like and anti-parallel interface, and force the formation of the homodimer consisted of two subunits. The main feature of the interface can be described as the complementary hydrophobic interaction and backbone hydrogen bonds in the central portion of motif that are reinforced by an ion pair at each end of the motif. Accordingly, the mutation V260E of SULT2A1 is produced and then expressed. The molecular weights of wild-type and V260E mutant of SULT2A1 was then estimated by gel filtration. The retention volume (Ve) and the parameter (Kav) were shown as the TABLE I. The retention volume of the blue dextran 2000 was 38.1 ml and the Kav could be estimated by the equation: Kav = (Ve – V0) / (Vt – V0). The calibration curve was plotted as the independent variable was log (MW) versus the dependent variable Kav as shown in Fig. 1. The retention volume of the wild type and V260E mutant of SULT2A1 was 52.41 and 63.33 ml, respectively (TABLE II). The corresponding molecular weight according to the retention volume was shown in Fig. 1 and the calculated values were approximately equal to the theoretical molecular weight of dimer and monomer of SULT2A1 (TABLE II).. The estimated molecular weight of wild-type SULT2A1 was approximately. 2-fold than that of V260E mutant. These data indicated that the interface of dimerization was easily disrupted through site-directed mutagenesis to force the formation of monomeric SULT2A1 because the glutamic acid might interfere with the hydrophobic zipper-like KTVE motif as similar in previous study (12). Circular Dichroism Measurement—Analysis by circular dichroism spectrum reveals the. 27.
(39) conformational change in dimer and monomer of SULT2A1 whether in the secondary or the tertiary structure. The CD spectrum in the range of far-UV (200-240 nm) (Fig. 2A) indicated the secondary structure showed the slight difference between dimer and monomer in the 205-225 nm, where is the absorption of α-helix and β-sheet. Accordingly, the mutation V260E of SULT2A1, caused the conformational change towards the secondary structure. Furthermore, the tertiary structure of SULT2A1 was also monitored by the CD spectrum in the range of near-UV (240-320 nm) (Fig. 2B). The range in 270-290 nm also revealed the slight difference of tertiary structure because of the absorption of aromatic residue, such as phenylalanine, tyrosine, and tryptophane. The monomer mutant, V260E, caused the environmental change of the α-helixes, β-sheets, and aromatic residues. It may reveal that there is a bit difference between the dimer and monomer although it may affect the whole protein structure or partial quaternary protein structure located near the KTVE motif. Kinetic Analysis—Rate constants of dimer and V260E mutant of SULT2A1 were shown in TABLE III. Either the PAPS or the DHEA was examined in the experiment. Km for both the dimer and monomer apparently is similar but the catalytic efficiency (kcat/Km) revealed the dimer was higher than monomer for approximately 2 folds. This result indicated the dimer might competent catalytically in each subunit. Furthermore, the substrate inhibition for DHEA was similar either of the dimer and monomer was examined. These results revealed the each subunit of dimer and the monomer might possess the catalytic ability towards both PAPS and DHEA. The catalytic mechanism, such as the substrate inhibition, is also the same for dimer and monomer of SULT2A1. Dissociation Constants of PAP and DHEA in Binary and Ternary Complex of Wild-type and V260E Mutant of SULT2A1—So far the crystal structures of cytosolic sulfotransferases all indicate that they all contain the diverse substrate binding site and a separate but identical PAPS binding site (14-19). The dissociative ability of the SULT2A1/PAP complex should be. 28.
(40) identical to other cytosolic sulfotransferases. The monomer of SULT2A1, however, was still far from known that the dissociation constants towards the PAPS and DHEA, even the comparison with the dimer of SULT2A1. In this study we performed the dissociation constants of PAP in the binary complex (SULT2A1/PAP) and the DHEA in the ternary complex (SULT2A1/PAP/DHEA) (TABLE IV). Whether the dimer or monomer was examined, the Kd values towards PAP in the binary complex and DHEA in the ternary complex revealed no significant differences. Thermal Stability Dissociation Constants of PAP, DHEA and ADT in Binary and Ternary Dead-end Complex of SULT2A1—Thermal inactivation of SULT2A1 was studied over the temperature range of 25–50 ℃ after the incubation at each desired temperature for 15 min (Fig. 3). It was observed that the monomer mutant of SULT2A1 lost only 10% of the basal activity of the control after thermal treatment at 30 ℃ for 15 min. The obvious decreased activities towards dimer and monomer are at the temperature of 40 and 35 ℃, respectively. Also, there is a slight deviation to the previous data reported in the literatures that the tolerance of thermal inactivation was higher at the temperature of 43 ℃ (33) while the dimer of SULT2A1 was examined. It was obvious that the monomer of SULT2A1 was more labile than dimer because the temperature of 50 % thermal inactivation was approximately 35 ℃ for monomer and 40 ℃ for dimer, respectively. Urea-Dependent Unfolding Experiments—The dimer and monomer of SULT2A1 were examined under the unfolding induction by urea from 0-6 M at 4 ℃ for approximately 24 hours. Typical normalized urea-induction unfolding curves were shown in Fig. 4 and the data from such transition curves were fitted to Equation 3 and 4 to yield values for [Urea]50%, m value, and △GUNH2O for dimer and monomer mutant of SULT2A1 (TABLE V) (all errors are calculated from the best fit of the data and are not standard deviations from repetitive runs).. 29.
(41) Based on these △GUNH2O values for urea denaturation monitored by CD222nm in TABLE V, the thermodynamic stabilities of monomer is more labile than dimer of SULT2A1. Related parameters of urea-induced denaturation are m value, the slope of the transition, and [Urea]50%, the urea concentration at the midpoint of denaturations given in TABLE V. The [Urea]50% was obviously higher in dimer than in monomer mutant.. 30.
(42) DISCUSSION. The fact that specific protein-protein interaction can be mediated by short segments of polypeptide chains and it is pivotally important for many physiological processes. For example, upon phosphorylation, ERK2 (a MAP-kinase) forms a homodimer that translocates into the nucleus (this dimerization is mediated by a flexible loop of ERK2 molecule) (34). The key contact surface consists of 12 residues that constitute the hydrophobic zipper-like dimer interface enforced by ion pairs at each end of the zipper (35-36). So far the dimerization interface of cytosolic sulfotransferase had been identified as the KTVE motif (12) and these highly conserved amino acid residues form a zipper-like and anti-parallel interface, and force the formation of the homodimer consisted of two subunits. In this study we mutated the Val-260 to glutamic acid to interfere the hydrophobic interaction and caused the charge-charge repulsion. The V260E mutant of SULT2A1 forces the dissociation of the two subunits of dimer to form the monomer and confirmed by gel filtration as shown in Fig. 1 and TABLE II. The secondary and tertiary structures reveal slight difference between dimer and monomer by the measurement of circular dichroism spectrum (Fig. 2). However, the conformational change may result from the mutation itself because the glutamic acid (negative charge residue) is more hydrophilic than the valine (neutral and more hydrophobic). The site-directed point mutation (V260E) may cause partial conformational change so that there is the slight difference between dimer and monomer. Furthermore, it may also result from the protein-protein interaction between the two subunits of dimer of SULT2A1. Consequently the monomer loses the ability to approach with each other and the secondary and tertiary structure observed by CD spectrum is in fact the true ones without the interference by the other subunit.. 31.
(43) The enzyme kinetics reveal the Km and Ki values are identical whether the dimer or monomer is examined (TABLE III). It indicates the catalytic mechanism may be the same even the substrate inhibition reaction. The kcat/Km, however, is 2 folds higher in dimer than in monomer of SULT2A1. It reveals both the subunits of dimer are competent catalytically and it makes more senses. Moreover, previous study indicated that the binding study with human SULT1E1 (homodimer) is shown that the stoichiometry of PAP or PAPS binding is one ligand with one enzyme (26). It was conflicting that the dimer is half active and the rest one is inactive. Furthermore, the substrate binding study (TABLE IV) also reveals that the substrate binding affinity is identical either in PAP or DHEA between dimer and monomer. The each subunit of dimer does not affect the release of the PAP and DHEA, and it could also interpret that the substrate inhibition does not attribute to dimerization of SULT2A1 (37) although the origin of substrate inhibition had been proposed the formation of ternary dead-end complex (26, 38). The Ki values for either of dimer and monomer (TABLE III) indicates that substrate inhibition is not the result of dimerization and the release of DHEA does not significantly alter (TABLE IV). Thermal and conformational stability reveal the monomer is more labile and the thermal tolerance is lower than dimer approximately for 5 ℃ while the enzyme activity is inactive for 50 % (Fig. 3). The conformational stability examined by urea unfolding experiment shows the monomer is denatured under lower concentration of urea (Fig. 4). It may be attributed to the conformational change as described above but it is not clear to know that whether the protein-protein interaction forces the dimer more stable than monomer. Consequently, we have prepared the monomer of SULT2A1 by site-directed mutagenesis and confirmed by gel filtration. The conformational change, enzyme kinetics, thermal inactivation and conformational stability experiments indicate the monomer and dimer possess slight differences and it is potential to be the model to study the quaternary structure. 32.
(44) of cytosolic sulfotransferases and further to pursue the significance and the physiological function.. 33.
(45) REFERENCES. 1. Mulder, G. J., and Jakoby, W. B. (1990) in Conjugation Reactions in Drug Metabolism (Mulder, G. J., ed) pp. 107–161, Taylor and Francis, Ltd., London 2. Falany, C., and Roth, J. A. (1993) in Human Drug Metabolism: From Molecular Biology to Man (Jeffery, E. H., ed) pp. 101–115, CRC Press, Inc. Boca Raton, FL 3. Weinshilboum, R., and Otterness, D. (1994) in Conjugation-Deconjugation Reactions in Drug Metabolism and Toxicity (Kaufmann, F. C., ed) pp. 45–78, Springer-Verlag, Berlin 4. DeBaun, J. R., Miller, E. C., and Miller, J. A. (1970) N-hydroxy-2-acetylaminofluorene sulfotransferase: its probable role in carcinogenesis and in protein-(methion-S-yl) binding in rat liver. Cancer Res. 30, 577–595 5. Glatt, H. (1997) Sulfation and sulfotransferases 4: bioactivation of mutagens via sulfation. FASEB J. 11, 314–321 6. Falany, C. N. (1997) Sulfation and sulfotransferases. Introduction: changing view of sulfation and the cytosolic sulfotransferases. FASEB J. 11, 1–2 7. Falany, C. N. (1997) Enzymology of human cytosolic sulfotransferases. FASEB J. 11, 206–216 8. Weinshilboum, R. M., Otterness, D. M., Aksoy, I. A., Wood, T. C., Her, C., and Raftogianis, R. B. (1997) Sulfation and sulfotransferases 1: sulfotransferase molecular biology: cDNAs and genes. FASEB J. 11, 3–14 9. Chen, G., Banoglu, E., and Duffel, M. W. (1996) Influence of substrate structure on the catalytic efficiency of hydroxysteroid sulfotransferase STa in the sulfation of alcohols. Chem. Res. Toxicol. 9, 67–74 10. Kakuta, Y., Pedersen, L. C., Chae, K., Song, W. C., Leblanc, D., London, R., Carter, C. W., and Negishi, M. (1998) Mouse steroid sulfotransferases: substrate specificity and. 34.
(46) preliminary X-ray crystallographic analysis. Biochem. Pharmacol. 55, 313–317 11. Chang, H. J., Shi, R., Rehse, P., and Lin, S. X. (2004) Identifying androsterone (ADT) as a cognate substrate for human dehydroepiandrosterone sulfotransferase (DHEA-ST) important for steroid homeostasis: structure of the enzyme-ADT complex. J. Biol. Chem. 279, 2689–2696 12. Petrotchenko, E. V., Pedersen, L. C., Borchers, C. H., Tomer, K. B., and Negishi, M. (2001) The dimerization motif of cytosolic sulfotransferases. FEBS Lett. 490, 39–43 13. Kiehlbauch, C. C., Lam, Y. F., and Ringer, D. P. (1995) Homodimeric and heterodimeric aryl. sulfotransferases. catalyze. the. sulfuric. acid. esterification. of. N-hydroxy-2-acetylaminofluorene. J. Biol. Chem. 270, 18941–18947 14. Gamage, N. U., Duggleby, R. G., Barnett, A. C., Tresillian, M., Latham, C. F., Liyou, N. E., McManus, M. E., and Martin, J. L. (2003) Structure of a human carcinogen-converting enzyme, SULT1A1. Structural and kinetic implications of substrate inhibition. J. Biol. Chem. 278, 7655–7662 15. Lu, J. H., Li, H. T., Liu, M. C., Zhang, J. P., Li, M., An, X. M., and Chang, W. R. (2005) Crystal structure of human sulfotransferase SULT1A3 in complex with dopamine and 3'-phosphoadenosine 5'-phosphate. Biochem. Biophys. Res. Commun. 335, 417–423 16. Kakuta, Y., Pedersen, L. G., Carter, C. W., Negishi, M., and Pedersen, L. C. (1997) Crystal structure of estrogen sulphotransferase. Nat. Struct. Biol. 4, 904–908 17. Shevtsov, S., Petrotchenko, E. V., Pedersen, L. C., and Negishi, M. (2003) Crystallographic analysis of a hydroxylated polychlorinated biphenyl (OH-PCB) bound to the catalytic estrogen binding site of human estrogen sulfotransferase. Environ. Health Perspect. 111, 884–888 18. Rehse, P. H., Zhou, M., and Lin, S. X. (2002) Crystal structure of human dehydroepiandrosterone sulphotransferase in complex with substrate. Biochem. J. 364,. 35.
(47) 165–171 19. Lee, K. A., Fuda, H., Lee, Y. C., Negishi, M., Strott, C. A., and Pedersen, L. C. (2003) Crystal structure of human cholesterol sulfotransferase (SULT2B1b) in the presence of pregnenolone and 3'-phosphoadenosine 5'-phosphate. Rationale for specificity differences between prototypical SULT2A1 and the SULT2BG1 isoforms. J. Biol. Chem. 278, 44593–44599 20. Chang, H. J., Zhou, M., and Lin, S. X. (2001) Human dehydroepiandrosterone sulfotransferase: purification and characterization of a recombinant protein. J. Steroid Biochem. Mol. Biol. 77, 159–165 21. Sakakibara, Y., Takami, Y., Nakayama, T., Suiko, M., and Liu, M. C. (1998) Localization and functional analysis of the substrate specificity/catalytic domains of human M-form and P-form phenol sulfotransferases. J. Biol. Chem. 273, 6242–6247 22. Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680–685 23. Gill, S. C., and von Hippel, P. H. (1989) Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 182, 319–326 24. Chen, W. T., Liu, M. C., and Yang, Y. S. (2005) Fluorometric assay for alcohol sulfotransferase. Anal. Biochem. 339, 54–60 25. Yang, Y. S., Marshall, A. D., McPhie, P., Guo, W. X., Xie, X., Chen, X., and Jakoby, W. B. (1996) Two phenol sulfotransferase species from one cDNA: nature of the differences. Protein Expr. Purif. 8, 423–429 26. Zhang, H., Varlamova, O., Vargas, F. M., Falany, C. N., and Leyh, T. S. (1998) Sulfuryl transfer: the catalytic mechanism of human estrogen sulfotransferase. J. Biol. Chem. 273, 10888–10892 27. Jackson, S. E., Moracci, M., elMasry, N., Johnson, C. M., and Fersht, A. R. (1993) Effect. 36.
(48) of cavity-creating mutations in the hydrophobic core of chymotrypsin inhibitor 2. Biochemistry 32, 11259–11269 28. Wallace, L. A., Sluis-Cremer, N., and Dirr, H. W. (1998) Equilibrium and kinetic unfolding properties of dimeric human glutathione transferase A1-1. Biochemistry 37, 5320–5328 29. MacDonald, R. I., and Pozharski, E. V. (2001) Free energies of urea and of thermal unfolding show that two tandem repeats of spectrin are thermodynamically more stable than a single repeat. Biochemistry 40, 3974–3984 30. Cornish-Bowden, A. (1995) Analysis of Enzyme Kinetic Data, pp. 118–122, Oxford University Press, Oxford 31. Tanford, C. (1968) Protein denaturation. Adv. Protein Chem. 23, 121–282 32. Pace, C. N. (1986) Determination and analysis of urea and guanidine hydrochloride denaturation curves. Methods Enzymol. 131, 266–279 33. Kudlacek, P. E., Clemens, D. L., Halgard, C. M., and Anderson, R. J. (1997) Characterization of recombinant human liver dehydroepiandrosterone sulfotransferase with minoxidil as the substrate. Biochem. Pharmacol. 53, 215–221 34. Khokhlatchev, A. V., Canagarajah, B., Wilsbacher, J., Robinson, M., Atkinson, M., Goldsmith, E., and Cobb, M. H. (1997) Phosphorylation of the MAP kinase ERK2 promotes its homodimerization and nuclear translocation. Cell 93, 605–615 35. Scheufler, C., Brinker, A., Bourenkov, G., Pegoraro, S., Moroder, L., Bartunik, H., Hartl, F. U., and Moarefi, I. (2000) Structure of TPR domain-peptide complexes: critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell 101, 199–210 36. Cobb, M. H., and Goldsmith, E. J. (2000) Dimerization in MAP-kinase signaling. Trends Biochem. Sci. 25, 7–9 37. Pedersen, L. C., Petrotchenko, E. V., and Negishi, M. (2000) Crystal structure of. 37.
(49) SULT2A3, human hydroxysteroid sulfotransferase. FEBS Lett. 475, 61–64 38. Duffel, M. W., and Jakoby, W. B. (1981) On the mechanism of aryl sulfotransferase. J. Biol. Chem. 256, 11123–11127. 38.
(50) TABLE I Calibration curve determination of molecular weight on Sephacryl S-200 HR.. a. 4.83. Vea 51.77. Kav 0.17. 43000. 4.63. 57.26. 0.23. Chymotrypsinofen A. 25000. 4.40. 72.83. 0.42. Ribonuclease A. 13700. 4.14. 81.14. 0.53. MW. log(MW). Albumin. 67000. Ovabumin. b. Ve represented the retention volume of eluted protein under the condition as indicated under. “Experimental Procedures.” b. Kav was calculated by the equation: Kav = (Ve – V0) / (Vt – V0). The Vt was the column. volume as 120 ml and the V0 was the retention volume of blue dextran 2000 (2000 kDa), whose retention volume was 38.1 ml.. 39.
(51) TABLE II Physical and chemical parameters of wild type and monomer mutant of SULT2A1.. SULT2A1. a. Molecular weighta Theoretical pIa. Veb. Wild type. 68994. 5.5. 52.41. V260E. 34541. 5.3. 63.33. The molecular weight and theoretical isoelectric point (pI) were determined by the. ProtParam on the EXPASY server (http://tw.expasy.org/tools/protparam.html). b. The Ve indicated the retention volume as described above and determined by Sephacryl. S-200 HR.. 40.
(52) TABLE III Rate constants of PAPS and DHEA with wild-type and V260E mutant of SULT2A1a.. SULT2A1. PAPS Km. μM. a. k cat. DHEA k cat /K m. Wild type. 3.9 ± 0.4. min 8.6 ± 0.8. V260E. 4.2 ± 0.6. 4.4 ± 0.7. Km. μM. -1. 2.2 1.1. k cat -1. Ki. k cat /K m. μM. 4.7 ± 0.3. min 8.2 ± 0.7. 4.3 ± 0.3. 1.7. 5.6 ± 0.5. 4.2 ± 0.5. 4.9 ± 0.5. 0.8. The reaction mixture with a final volume of 1 ml consisted of 100 mM potassium phosphate. buffer at pH 7.0, 5 mM 2-mercaptoethanol, 20 μM PAPS, 2 mM MUS, 5.4 μg K65ER68G of rat SULT1A1, wild type or V260E mutant of SULT2A1 and 5 μM of DHEA at 37 ℃. A linear response was obtained when 14.9 nM (1 μg) SULT2A1 was added in the standard assay condition.. 41.
(53) TABLE IV Dissociation constants of PAP and DHEA in wild-type and V260E mutant of SULT2A1a.. Binary complexb. Ternary complexc. PAP. DHEA. nM. μM. Wild type. 40.7 ± 8.2. 0.8 ± 0.05. V260E. 36.5 ± 9.7. 0.9 ± 0.1. SULT2A1. a. Dissociation constants were determined with spectrofluorimeter as indicated under. “Experimental Procedures.” b. The dissociation constants of binary complex was determined with PAP ( 5 to 365 nM) and. SULT2A1 (100 nM). c. The dissociation constants of ternary complex were determined with DHEA (0.1 to 50 μM). and SULT2A1 (0.5 μM) in the presence of PAP (1 μM).. 42.
(54) TABLE V Thermodynamic parameters correlated with free energy of unfolding, △GUNH2O.. H2O c. [Urea]50%a. mb. M. kcal mol-2. kcal mol-1. Wild type. 2.7. 2.3 ± 0.3. 6.3 ± 0.8. V260E. 1.1. 1.5 ± 0.2. 1.6 ± 0.5. SULT2A1. △GUN. a. [Urea]50% is the urea concentration at the midpoint of urea denaturation.. b. m value is a constant that is proportional to the increase in the degree of exposure of the. protein on denaturation. c. △GUNH2O is the free energy of denaturation in the absence of urea, and is the product of. [Urea]50% and m value.. 43.
(55) 0.6 0.5. Kav. 0.4 0.3 0.2 0.1 0.0 4.0. 4.2. 4.4. 4.6. 4.8. 5.0. log(MW). Figure 1. Calibration curve of Sephacryl S-200 HR. A molecular weight calibration curve, which defined the relationship between the elution volumes of a set standards and the logarithm of their respective molecular weights, was determined with Sephacryl S-200 HR. Calibration standards (crisscross) used were ribonuclease A (15.6 KDa), chymotrypsinogen A (19.4 KDa), ovalbumin (47.6 KDa), and Albumin (62.9 KDa). The wild type and V260E mutant were closed and open diamonds, respectively. The Kav is obtained from the equation Kav = (Ve – V0) / (Vt – V0) as described above.. 44.
(56) A. 2. 0. -1. 6. [θ] (10 deg cm /dmol). 1. -2. -3. -4 200. 210. 220. 230. 240. 300. 320. Wavelength (nm) B. 2. 0. 4. [θ] (10 deg cm /dmol). 2. -2. -4. 240. 260. 280. Wavelength (nm). 45.
(57) Figure 2. Circular dichroism specrtra of wild-type and V260E mutant of SULT2A1. CD measurements were carried out on a Jasco J715 spectropolarimeter and processed data with J-700 standard analysis version 1.33.00 (Jasco, Japan). CD spectra of enzymes at a concentration of 1 mg ml-1 in the phosphate buffer (pH 7.0) were measured in a (A) 1 mm light path length quartz cell in the far-UV range (200-240 nm) and (B) 1 cm light path in the near-UV (240-320 nm) at room temperature individually. The closed and open diamonds indicated the wild type and V260E mutant of SULT2A1, respectively.. 46.
數據



+7
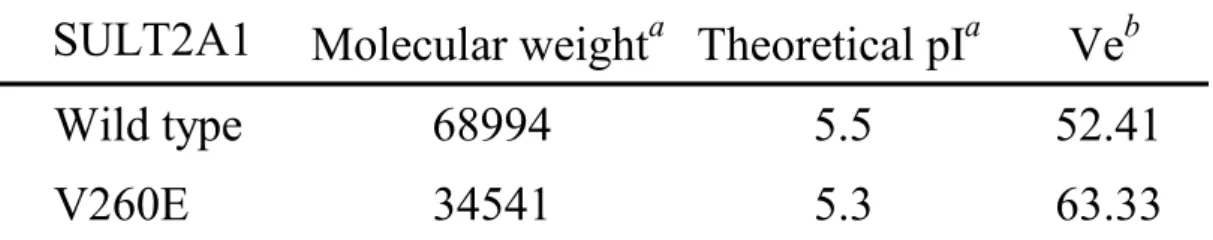

Outline
相關文件
The ontology induction and knowledge graph construction enable systems to automatically acquire open domain knowledge. The MF technique for SLU modeling provides a principle model
• To enhance teachers’ knowledge and understanding about the learning and teaching of grammar in context through the use of various e-learning resources in the primary
Xianggang zaji (miscellaneous notes on Hong Kong) was written by an English and translated into Chinese by a local Chinese literati.. Doubts can therefore be cast as to whether
vs Functional grammar (i.e. organising grammar items according to the communicative functions) at the discourse level2. “…a bridge between
(b) reviewing the positioning of VPET in the higher education system in Hong Kong, exploring the merits of developing professional vocational qualifications at the degree
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
In this paper, we build a new class of neural networks based on the smoothing method for NCP introduced by Haddou and Maheux [18] using some family F of smoothing functions.
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
