Novel pterostilbene derivative ANK-199 induces autophagic cell
death through regulating PI3 kinase class III/Beclin 1/Atg-related
proteins in cisplatin-resistant human oral cancer CAR cells
MIN-TSANG HSIEH1,2, HAO-PING CHEN3, CHI-CHENG LU4, JO-HUA
CHIANG5, TIAN-SHUNG WU5, DAIH-HUANG KUO6, LI-JIAU HUANG7,
SHENG-CHU KUO7, JAI-SING YANG8
1 School of Pharmacy, 7Graduate Institute of Pharmaceutical
Chemistry, 8Department of Pharmacology, China Medical University,
Taichung 404; 2 Chinese Medicinal Research and Development
Center, China Medical University Hospital, Taichung
404;3Department of Biochemistry, Tzu Chi University, Hualien 970; 4Department of Food Science and Biotechnology, National Chung
Hsing University, Taichung 402; 5Department of Chemistry, National
Cheng Kung University, Tainan 701; 6Department of Pharmacy and
Graduate Institute of Pharmaceutical Technology, Tajen University, Pingtung, Taiwan, ROC
Abstract. Pterostilbene is a natural analogue of phenolic phytoalexin and an effective chemopreventive agent against multiple types of cancer cells. Recently, a series of pterostilbene derivatives were designed and synthesized in our laboratory as novel anticancer agents. Here, we studied the antitumor activity and the molecular mechanism of the pterostilbene derivative ANK-199 in cisplatin-resistant human oral cancer CAR cells. ANK-199 increased cell death and determined by MTT assay in a concentration- and time-dependent response. On the contrary, ANK-199 exerts low cytotoxicity against normal oral cells. ANK-199 induced the cell autophagy markers, the autophagic vacuoles and acidic vesicular organelle (AVO), which were manifested by monodansylcadaverine (MDC) and acridine orange (AO) staining, as well as cleavage of LC3. Autophagy related signal molecules were up-regulated in ANK-199-treated CAR cells. The levels of Atg7, Atg12, Beclin 1 and LC-3-II mRNA were up-regulated by real-time PCR assay. Moreover, ANK-199 significantly enhanced 26 gene expressions and suppressed 96 gene expressions, which were determined by cDNA microarray. Based on the results, we identified numerous differentially expressed genes related to regulation of cellular component biogenesis, actin cytoskeleton organization, regulation of protein complex assembly, regulation of actin filament-based process, regulation of cytoskeleton organization, regulation of actin polymerization or depolymerization, regulation of actin filament length and cell signaling pathways. In conclusion, ANK-199 triggered autophagy death through the PI3 kinase class III/Beclin 1 and Atgs signaling pathway in CAR cells. Our data demonstrated that ANK-199 may be potentially efficacious in the treatment of cisplatin-resistant human oral cancer.
cisplatin-resistant CAR human oral cancer cells Introduction
Pterostilbene is a natural stilbenoid compound of phenolic phytoalexin analogue and found in narra tree, grape, blueberries, etc. . It has been studied for its biologic effects, including anticancer activity with low intrinsic toxicity , anti-inflammatory , anti-oxidative effect , regulation of neutrophil function and against free radical mediated oxidative damage . The mechanisms of the anticancer action have been investigated and include pro-apoptosis, pro-autophagy, telomerase inhibition, DNA damage, anti-angiogenesis, anti-metastasis, and immuno-stimulatory effects . Recently, pterostilbene induces apoptosis in pancreatic cancer cells , breast cancer MCF-7 cells , docetaxel-induced multiple drug resistance (MDR) lung cancer cells , osteosarcoma cells , prostate cancer PC-3 and LNCaP cells , leukemia K562 cells, MDR and BCR-ABL-expressing leukemia cells , colon cancer cells , hepatocellular carcinoma cells and, and gastric carcinoma cells . However, it is also reported that pterostilbene induces autophagic death in leukemia HL60 and MOLT4 cells , lung cancer cells , colon cancer HT29 cells , breast cancer MCF-7 cells , bladder cancer cells , and vascular endothelial cells .
Three major processes lead to cell death, which is divided into apoptosis, autophagy and necrosis . Autophagic cell death plays an important role in normal physiology in animals . During the cells experience damage, nutrient starvation, pathogen infection and aging, the autophagic death is required for the support of cellular survival or death . Autophagy causes a specific morphological and biochemical modification. The cytoplasm and phagophore of various organelles are sequestered by a membrane to form an autophagosomes . The autophagosome fuses with the lysosome then forming autophagolysosome, eventually resulting in
degradation of the captured proteins or organelles by lysosomal enzymes . The autophagy-related proteins (Atgs) are involved in autophagic cell death . The membrane nucleation is mediated by a phosphatidylinositol 3-kinase (PI3K) class III, Beclin 1 (the mammalian ortholog of yeast ATG6), Rubicon and Atg 14 . The production of autophagosome is necessary for the employment of Atg16 L1-Atg12-Atg7-Atg5 complex and microtubule-associated protein 1 light chain 3 type II (LC3 II) (membrane-bound form) . Once cells undergo autophagic cell death, an autophagosomal marker LC3-II increases from the conversion of LC3-I . Hence, induction of autophagic death for cancer cells is one of the best strategies in chemotherapy .
To improve the anticancer efficacy and safety of pterostilbene (Figure 1A), our laboratory designed and synthesized a novel pterostilbene derivative ANK-199 (Figure 1B). The aim of this study is to discover whether the anti-oral cancer activity of ANK-199 mediates through the direct cytotoxic effect and to understand the molecular mechanisms in cisplatin-resistant CAR human oral cancer cells. This study is focused on the autophagic death-induced by ANK-199 in CAR. Our results demonstrate that ANK-199 inhibits cells viability and induces autophagy through the PI3 kinase class III/ Beclin 1/Atgs signals.
Materials and Methods
Chemicals and reagents. Dimethyl sulfoxide (DMSO), methyladenine (MA),
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), monodansylcadaverine (MDC), cisplatin, β-actin antibody, and Tween20 were obtained from Sigma-Aldrich Corp. (St. Louis, MO, USA). Fetal bovine serum (FBS), L-glutamine, Penicillin/streptomycin, Dulbecco’s Modified Eagle Medium (DMEM), acridine orange (AO), and trypsin-EDTA were purchased from Life
Technologies (Carlsbad, CA, USA). The primary antibodies (anti-Atg 5, anti-Atg 7, anti-Atg 12, anti-Atg 14, anti-Atg 16L1, anti-Beclin 1, anti-PI3K class III, anti-LC-3-II, and anti-Rubicon) were obtained from Cell Signaling Technology (Danvers, MA, USA), and the horseradish peroxidase (HRP)-conjugated secondary antibodies against rabbit or mouse immunoglobulin for Western blotting were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). ANK-199 [4-(3,5-dimethoxystyryl)phenyl 3-hydroxy-2-(hydroxymethyl)-2-methylpropanoate] was synthesized and supplied by Dr. Sheng-Chu Kuo.
Cell culture. The human oral cancer cell line CAL 27 was obtained from the
American Type Culture Collection (ATCC, Manassas, VA, USA). CAR, a cisplatin-resistant cell line, was established by clonal selection of CAL 27 using 10 cycles of 1 passage treatment with 10-80 μM of cisplatin followed by a recovery period of another passage . CAR cells were cultivated in DMEM supplemented with 10% FBS, 100 μg/ml streptomycin, 100 Units/ml penicillin, 2 mM L-glutamine and 80 μM cisplatin. Human normal gingival fibroblasts cells (HGF) and human normal oral keratinocyte cells (OK) were kindly provided by Dr. Tzong-Ming Shieh (Department of Dental Hygiene, China Medical University). HGF and OK cells were cultivated in DMEM as previously described for our study.
Cell viability and morphological examination. CAR cells (1x104 cells) into a 96-well
plate were incubated with 0, 25, 50, 75 and 100 μM of ANK-199 for 24, 48 and 72 h. For incubation with the autophagy inhibitor, cells were pretreated with 3-MA (10 mM) for 1 h, followed by treatment with or without ANK-199 (50 and 75 μM) for 48 h. After washing the cells, DMEM containing MTT (0.5 mg/ml) of was added to detect viability as previously described . The cell viability was expressed as % of the
control. Cell morphological examination of autophagic vacuoles was determined utilizing a phase-contrast microscope.
Detections of autophagic vacuoles by MDC and acidic vesicular organelles (AVO) with AO staining. CAR cells were seeded on sterile coverslips in tissue culture plates
with a density of 5x104 cells/per coverslip. After 0, 50, 75 μM of ANK-199 treatment
for 48 h, cells were stained with either 1 μg/ml AO or 0.1 mM MDC at 37 °C for 10 mins. Cells were immediately visualized by fluorescence microscopy (Nikon, Melville, NY, USA) for the detection of autophagic vacuoles and AVO occurrences.
Autophagy assay by LC3B-GFP imaging and nuclear stain. The induction of
autophagy was detected with the Premo Autophagy Sensor (LC3B-GFP) BacMam 2.0 kit (Molecular Probes/Life Technologies). CAR cells were seeded on sterile coverslips in tissue culture plates with a density of 1x104 cells/per coverslip. After
CAR cells were transfected with LC3B-GFP in accordance with the manufacturer’s protocol, cells were treated with 0, 50 and 75 μM of ANK-199 for 48 h. Cells were then fixed on ice with 4% paraformaldehyde, and the slides were mounted and analyzed by a fluorescence microscope as described elsewhere . After treatments, cells were stained with 4’,6-diamidino-2-phenylindole (DAPI, Molecular Probes/Life Technologies) and photographed using a fluorescence microscope .
Western blotting analysis. CAR cells (1x107 cells/75-T flask) were treated with
ANK-199 (50 and 75 μM) for 48 h. At the end of incubation, the total proteins were prepared, and the protein concentration was measured by using a BCA assay kit (Pierce Chemical, Rockford, IL, USA). Equal amounts of cell lysates were run on 10% SDS-polyacrylamide gel electrophoresis and further employed by
immunoblotting as described by Lai et al. .
Real-time PCR analysis. CAR cells at a density of 5 × 106 in T75 flasks were
incubated with or without 50 and 75 μM of ANK-199 for 24 h. Cells were collected, and total RNA was extracted by the QIAGEN RNeasy Mini Kit (QIAGEN Inc., Valencia, CA, USA). Each RNA sample was individually reverse-transcribed using the High Capacity cDNA Reverse Transcription Kits (Applied Biosystems, Foster City, CA, USA). Quantitative PCR was assessed for amplifications with 2X SYBR Green PCR Master Mix (Applied Biosystems), as well as forward and reverse primers for Atg7, Atg12, Beclin 1 and LC3 II gene. (Human
ATG7-F-CAGCAGTGACGATCGGATGA; Human
ATG7-R-GACGGGAAGGACATTATCAAACC; Human
ATG12-F-TGTGGCCTCAGAACAGTTGTTTA; Human
ATG12-R-CGCCTGAGACTTGCAGTAATGT; Human
BECN1-F-GGATGGTGTCTCTCGCAGATTC; Human
BECN1-R-GGTGCCGCCATCAGATG; Human LC3-II-F-CCGACCGCTGTAAGGAGGTA; Human LC3-II-R-AGGACGGGCAGCTGCTT) Applied Biosystems 7300 Real-Time PCR System was run in triplicate, and each value was expressed in the comparative threshold cycles (CT) method for the housekeeping gene GAPDH.
cDNA microarray analysis. CAR cells (5 × 106 per T75 flask) were incubated with or
without 75 μM of ANK-199 for 24 h. Cells were scraped and collected by centrifugation. The total RNA was subsequently isolated as mentioned above, and the purity were assessed at 260 nm and 280 nm using a Nanodrop (ND-1000; Labtech. International). Each sample (300 ng) was amplified and labeled using the GeneChip WT Sense Target Labeling and Control Reagents (900652) for Expression Analysis.
Hybridization was performed against the Affymetrix GeneChip Human Gene 1.0 ST array. The arrays were hybridised for 17 h at 45 °C and 60 rpm. Arrays were subsequently washed (Affymetrix Fluidics Station 450), stained with streptavidin-phycoerythrin(GeneChip Hybridization, Wash, and Stain Kit, 900720), and scanned on an Affymetrix GeneChip Scanner 3000. Resulting data was analyzed by using Expression Console software (Affymetrix) with default RMA parameters. Genes regulated by ANK-199 were determined with a 1.5-fold change. For detection of significantly over-represented GO biological processes, the DAVID functional annotation clustering tool (http://david.abcc.ncifcrf.gov) was used (DAVID Bioinformatics Resources 6.7). Enrichment was determined at DAVID calculated Benjamini value < 0.05. Significance of over-expression of individual genes was determined.
Statistical analysis. All the statistical results were expressed as the mean ± S.E.M. of
triplicate samples. Statistical analyses of data were done using one-way ANOVA followed by Student’s t-test, and *** p < 0.001 was considered significant.
Results
ANK-199 exhibits cytotoxicity and inhibits viable CAR cells. Cells were treated with
ANK-199 (0, 25, 50, 75 and 100 μM) for 24, 48 and 72 h. ANK-199 concentration-and time-dependently decreased cell viability of CAR cells (Figure 2A). The half maximal (50%) inhibitory concentration (IC50) for a 24, 48 and 72-h treatment of
ANK-199 in CAR cells were 106.21±3.21, 73.25±4.20 and 32.58±2.39 μM. To examine whether ANK-199 induced apoptosis in CAR cells, cells were treated with or without ANK-199 (50 and 75 μM) for 48 h. Our results showed that no evidence of DNA fragmentation was observed in treated cells (data not shown). Therefore,
ANK-199 did not induce apoptosis in CAR cells. Next, we examine whether autophagic cell death is involved in ANK-199-induced cell death. Cells were treated with 50 μM ANK-199 for 24, 48 and 72 h and then examined the autophagic vacuoles occurrence. As shown in Figure 2B, ANK-199 induced the formation of autophagic vacuoles in CAR cells in a time-dependent manner. Importantly, ANK-199 has less toxic (no viability impact and morphological traits) in normal oral cell lines, including HGF (IC50 > 100 μM) (Figure 3A) and OK cells (IC50 > 100 μM) (Figure 3B). Thus,
ANK-199-induced cell death of CAR cells proceeds through autophagic death instead of apoptosis.
ANK-199 induces autophagic cell death in CAR cells. We further confirmed the
formation of autophagosome vesicles in ANK-199-treated CAR cells. We detected the autophagic cell death caused by ANK-199 using MDC staining, which is popular fluorescent marker that preferentially accumulates in autophagic vacuoles . Cells treated with 50 and 75 µM of ANK-199 for 48 h clearly showed autophagic vacuoles, while few autophagic vacuole was observed in the control group (Figure 4A). We also found that the MDC intensity enhanced in a dose-dependent response. We also confirmed the ANK-199-troggered autophagic cell death using AO staining. In Figure 4B, AO staining of ANK-199-treated CAR cells clearly showed AVOs within the cytoplasm compared to control. Microtubule-associated protein 1 light-chain 3 (LC3) is an autophagic membrane marker for the detection of early autophagosome formation . We examined the LC3 distribution in 199-treated CAR cells. ANK-199 treatment enhanced the punctate pattern of LC3-GFP in autophagic CAR cells (Figure 4C). In addition, CAR cells were treated with ANK-199 (50 and 75 μM) for 48 h and thereafter analyzed DNA condensation. In Figure 4D, no effect of DNA
condensation was observed in CAR cells treated with ANK-199 for 48 h. Our results suggest that ANK-199-induced cell death in CAR cells is dependent on the induction of autophagic death.
ANK-199 up-regulates the autophagy-associated protein levels in CAR cells. Induced
autophagic cell death is associated with the important protein levels, such as Atg complex (Atg5, Atg7, Atg12, Atg14, and Atg16 L1), Beclin-1, PI3K class III, Rubicon and LC-3. We investigated the protein levels in ANK-199-treated CAR cells. As shown in Figure 5, ANK-199 at 50 and 75 μM increased the protein levels of Atg5, Atg7, Atg12, Atg14, and Atg16 L1), Beclin-1, PI3K class III and LC-3, but it decreased the protein level of Rubicon in CAR cells. Our results imply that ANK-199 induced autophagic cell death in CAR cells through interfering with the kinase class III/Beclin 1/Atg-associated signals pathway.
ANK-199 stimulates the autophagy-associated mRNA levels in CAR cells. We also
investigated the autophagy-associated gene of mRNA levels in ANK-199-treated CAR cells. As shown in Figure 6, we demonstrated that ANK-199 increased the level of Atg7 gene (Figure 6A), Atg12 gene (Figure 6B), Beclin-1 gene (Figure 6C) and
LC-3II gene (Figure 6D) expression in CAR cells
3-MA protects against autophagy in ANK-199-treated CAR cells. 3-MA, an inhibitor
of PI3K kinase class III, is presented to potently inhibit autophagy-dependent protein degradation and suppress the formation of autophagosomes . In this study, CAR cells were pretreated with 3-MA and then exposed to 50 or 75 μM of ANK-199 before measuring the levels of autophagic vacuoles and cell viability. Our results showed that 3-MA can inhibit autophagic vacuoles (Figure 7A) and is able to reduce cell
viability (Figure 7B) in ANK-199-treated CAR cells. Based on the data, we suggest that ANK-199-induced autophagy in CAR cells is mediated through interfering with the kinase class III.
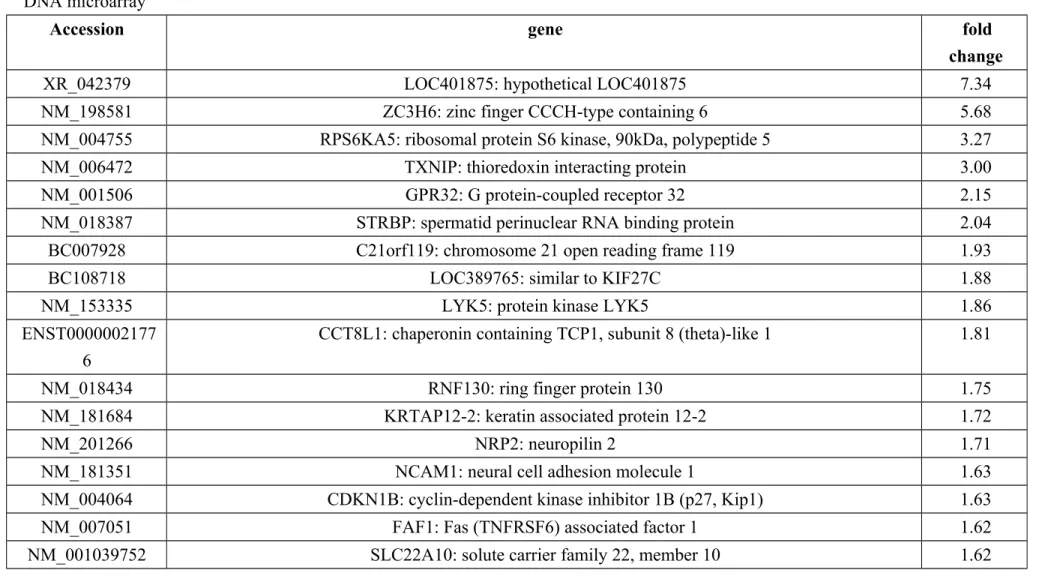
Microarray analysis. The analysis of cDNA microarray was analyzed to examine the
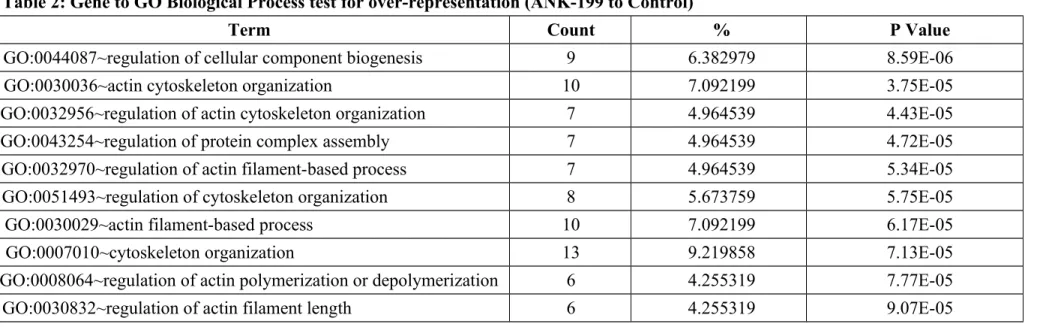
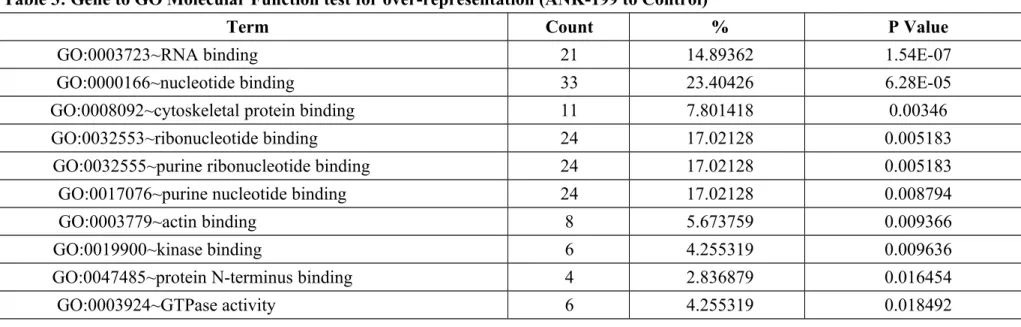
expressed genes in ANK-199-treated CAR cells. In Table 1, we showed that the transcripts of 26 genes were up-regulated, while these of 96 genes were down-regulated in ANK-199-treated CAR cells, respectively. Our results in Table 2 showed that ANK-199 regulated the expression of important genes and the biological process include cellular component biogenesis, actin cytoskeleton organization, regulation of protein complex assembly, regulation of actin filament-based process, regulation of cytoskeleton organization, regulation of actin polymerization or depolymerization, regulation of actin filament length. In Table 3, Gene to GO Molecular Function test for over-representation in ANK-199-treated CAR cells include RNA binding (21 genes); nucleotide binding (33 genes); cytoskeletal protein binding (11 genes); ribonucleotide binding (24 genes); purine ribonucleotide binding (24 genes); purine nucleotide binding (24 genes); actin binding (8 genes); kinase binding (6 genes); protein N-terminus binding (4 genes); GTPase activity (6 genes).
Discussion
In Taiwan, betel-quid and areca-nut chewing are a very important factor related to oral cavity carcinoma, which is the fourth-most common cause of cancer death in males . The therapy of HNSCC is combination with surgery, radiotherapy and/or chemotherapy . The clinical success of cisplatin-based chemotherapy, mostly in combination with 5-fluorouracil (5-FU) or taxane (paclitaxel and docetaxel), is however limited by multi-factorial intrinsic or acquired resistance . Natural products
powerful to prevent or therapy cancer are obvious with their high anticancer activity and low toxicity in normal cells . Pterostilbene, a naturally occurring dimethylether analogue of resveratrol, has been demonstrated to have anticancer property . The human clinical trial demonstrated that pterostilbene is safe in doses up to 250 mg/day . It was reported that pterostilbene suppresses cancer cell growth through alteration of the cell cycle, induction of apoptosis and autophagy or inhibition of cell metastasis and anti-angiogenesis in in vitro models . In in vivo tests, pterostilbene inhibits tumorigenesis and metastasis with minor toxicity . Pterostilbene deserves more investigation in preclinical studies or clinical trial as a potential anticancer agent . Recently, a series of pterostilbene derivatives were synthesized as less toxic anticancer candidates. Our results demonstrated that ANK-199 has greater cytotoxicity than that of the other compounds in human cisplatin-resistant CAR cells. In this study, we also investigated the molecular mechanism of autophagic induction by ANK-199 in CAR cells. ANK-199 caused the anti-prolifernation effects of 10-80% at 25, 75 and 100 μM for 24, 48 and 72 h on CAR cells, respectively (Figure 2). In addition, ANK-199 has much less cytotoxicity in the normal oral cells (HGF and OK) than that in CAR cells (Figure 3). This is a novel finding regarding that ANK-199 represents a promising candidate as an anti-oral cancer drug with low toxicity to normal cells.
It was report that accumulation of autophagic vacuoles and autophagy induced by pterostilbene occurred in HL60 human leukemia cells . Chen et. al. showed that pterostilbene-mediated chemopreventive effects in vivo were a result of the inhibition of epidermal growth factor receptor (EGFR) pathways and induction of apoptosis and autophagy during urethane-induced lung tumorigenesis. Chakraborty et. al. reported that pterostilbene induced autophagy in MCF-7 cells at 72 h treatment. Zhang et. al.
also demonstrated that pterostilbene promotes autophagy in vascular endothelial cells, and their group suggest pterostilbene could serve as a potential lead compound for developing a class of autophagy regulator as autophagy-related diseases therapy. In the present study, ANK-199 induced growth inhibitory effects in CAR cells and caused autophagic characteristics, for example, autophagic vesicle formation (Figure 4A), acidic vesicular organelles (Figure 4B), cleavage of LC3 (Figure 4C) and elevated protein levels of autophagic proteins, Atg complex, Beclin-1, PI3K class III and LC-3-II (Figure 5). ANK-199 also increased the mRNA levels of autophagic genes Atg 7, Atg 12, Beclin-1 and LC-3-II (Figure 6). Importantly, ANK-199 cells were pre-treated with 3-MA (a specific inhibitor of PI3K kinase class III) followed by treating with ANK-199, growth inhibitory effects and autophagic vesicle formation were significantly decreased compared with the ANK-199 alone treatment group (Figure 7). We used the cDNA microarray was analyzed to examine the expressed genes in CAR cells after treatment with 199, and the results showed that ANK-199 induced cell autophagic cell death may through cellular component biogenesis and actin cytoskeleton regulation (Table 1-3). Our results did not rule out ANK-199 might be involved in apoptotic or other signaling pathways after longer exposure (more than 72 h), but this study showed that CAR cells induced autophagic cell death after the 24 and 48 h treatment with ANK-199. The main reason of ANK-199-triggered autophagy is: (i) ANK-199 is a pterostilbene derivative; (ii) ANK-199 keeps the pharmacologic effect of pterostilbene.
Overall, the molecular signaling pathway could be summarized in Figure 8. Our results demonstrated that the PI3 kinase classIII/Beclin 1/Atgs protein regulation promotes autophagic cell death in ANK-199-treated CAR cells. These findings imply that ANK-199 may be used as a novel therapeutic reagent for the treatment of oral
cancer.
Acknowledgements
This work was supported part by a research grant from the National Science Council of the Republic of China (NSC ) awarded to SHENG-CHU KUO
References
1. McCormack D and McFadden D: Pterostilbene and cancer: current review. J Surg Res 173: e53-61, 2012.
2. Hong BH, Wu CH, Yeh CT and Yen GC: Invadopodia-associated proteins blockade as a novel mechanism for 6-shogaol and pterostilbene to reduce breast cancer cell motility and invasion. Mol Nutr Food Res 57: 886-895, 2013.
3. Mena S, Rodriguez ML, Ponsoda X, Estrela JM, Jaattela M and Ortega AL: Pterostilbene-induced tumor cytotoxicity: a lysosomal membrane permeabilization-dependent mechanism. PLoS One 7: e44524, 2012.
4. Mak KK, Wu AT, Lee WH, et al: Pterostilbene, a bioactive component of blueberries, suppresses the generation of breast cancer stem cells within tumor microenvironment and metastasis via modulating NF-kappaB/microRNA 448 circuit. Mol Nutr Food Res 57: 1123-1134, 2013.
5. Chen RJ, Tsai SJ, Ho CT, et al: Chemopreventive effects of pterostilbene on urethane-induced lung carcinogenesis in mice via the inhibition of EGFR-mediated pathways and the induction of apoptosis and autophagy. J Agric Food Chem 60: 11533-11541, 2012.
6. Huang CS, Ho CT, Tu SH, et al: Long-term ethanol exposure-induced hepatocellular carcinoma cell migration and invasion through lysyl oxidase activation are attenuated by combined treatment with pterostilbene and curcumin
analogues. J Agric Food Chem 61: 4326-4335, 2013.
7. Hsu CL, Lin YJ, Ho CT and Yen GC: The inhibitory effect of pterostilbene on inflammatory responses during the interaction of 3T3-L1 adipocytes and RAW 264.7 macrophages. J Agric Food Chem 61: 602-610, 2013.
8. McCormack D, McDonald D and McFadden D: Pterostilbene ameliorates tumor necrosis factor alpha-induced pancreatitis in vitro. J Surg Res 178: 28-32, 2012. 9. McCormack D and McFadden D: A review of pterostilbene antioxidant activity
and disease modification. Oxid Med Cell Longev 2013: 575482, 2013.
10. Perecko T, Drabikova K, Lojek A, et al: The effects of pterostilbene on neutrophil activity in experimental model of arthritis. Biomed Res Int 2013: 106041, 2013.
11. Acharya JD and Ghaskadbi SS: Protective effect of Pterostilbene against free radical mediated oxidative damage. BMC Complement Altern Med 13: 238, 2013.
12. Tippani R, Prakhya LJ, Porika M, Sirisha K, Abbagani S and Thammidala C: Pterostilbene as a Potential Novel Telomerase Inhibitor: Molecular Docking Studies and its In Vitro Evaluation. Curr Pharm Biotechnol 2014.
13. Pan MH, Chiou YS, Chen WJ, Wang JM, Badmaev V and Ho CT: Pterostilbene inhibited tumor invasion via suppressing multiple signal transduction pathways in human hepatocellular carcinoma cells. Carcinogenesis 30: 1234-1242, 2009. 14. Elango B, Devibalan S, Sekar TV, Palanisamy R, Ramasamy P and Mohanram
RK: Therapeutic potential of pterostilbene against pancreatic beta-cell apoptosis through Nrf2 mechanism. Br J Pharmacol 2014.
15. Chakraborty A, Bodipati N, Demonacos MK, Peddinti R, Ghosh K and Roy P: Long term induction by pterostilbene results in autophagy and cellular differentiation in MCF-7 cells via ROS dependent pathway. Mol Cell Endocrinol
355: 25-40, 2012.
16. Hsieh MJ, Lin CW, Yang SF, et al: A combination of pterostilbene with autophagy inhibitors exerts efficient apoptotic characteristics in both chemosensitive and chemoresistant lung cancer cells. Toxicol Sci 137: 65-75, 2014.
17. Liu Y, Wang L, Wu Y, et al: Pterostilbene exerts antitumor activity against human osteosarcoma cells by inhibiting the JAK2/STAT3 signaling pathway. Toxicology 304: 120-131, 2013.
18. Lin VC, Tsai YC, Lin JN, et al: Activation of AMPK by pterostilbene suppresses lipogenesis and cell-cycle progression in p53 positive and negative human prostate cancer cells. J Agric Food Chem 60: 6399-6407, 2012.
19. Tolomeo M, Grimaudo S, Di Cristina A, et al: Pterostilbene and 3'-hydroxypterostilbene are effective apoptosis-inducing agents in MDR and BCR-ABL-expressing leukemia cells. Int J Biochem Cell Biol 37: 1709-1726, 2005. 20. Nutakul W, Sobers HS, Qiu P, et al: Inhibitory effects of resveratrol and
pterostilbene on human colon cancer cells: a side-by-side comparison. J Agric Food Chem 59: 10964-10970, 2011.
21. Pan MH, Chang YH, Badmaev V, Nagabhushanam K and Ho CT: Pterostilbene induces apoptosis and cell cycle arrest in human gastric carcinoma cells. J Agric Food Chem 55: 7777-7785, 2007.
22. Siedlecka-Kroplewska K, Jozwik A, Boguslawski W, et al: Pterostilbene induces accumulation of autophagic vacuoles followed by cell death in HL60 human leukemia cells. J Physiol Pharmacol 64: 545-556, 2013.
23. Chen RJ, Ho CT and Wang YJ: Pterostilbene induces autophagy and apoptosis in sensitive and chemoresistant human bladder cancer cells. Mol Nutr Food Res 54: 1819-1832, 2010.
24. Zhang L, Cui L, Zhou G, Jing H, Guo Y and Sun W: Pterostilbene, a natural small-molecular compound, promotes cytoprotective macroautophagy in vascular endothelial cells. J Nutr Biochem 24: 903-911, 2013.
25. Inoue H and Tani K: Multimodal immunogenic cancer cell death as a consequence of anticancer cytotoxic treatments. Cell Death Differ 21: 39-49, 2014.
26. Lemasters JJ, Nieminen AL, Qian T, et al: The mitochondrial permeability transition in cell death: a common mechanism in necrosis, apoptosis and autophagy. Biochim Biophys Acta 1366: 177-196, 1998.
27. Melendez A and Neufeld TP: The cell biology of autophagy in metazoans: a developing story. Development 135: 2347-2360, 2008.
28. Klionsky DJ and Emr SD: Autophagy as a regulated pathway of cellular degradation. Science 290: 1717-1721, 2000.
29. Huang WW, Tsai SC, Peng SF, et al: Kaempferol induces autophagy through AMPK and AKT signaling molecules and causes G2/M arrest via downregulation of CDK1/cyclin B in SK-HEP-1 human hepatic cancer cells. Int J Oncol 42: 2069-2077, 2013.
30. Lin C, Tsai SC, Tseng MT, et al: AKT serine/threonine protein kinase modulates baicalin-triggered autophagy in human bladder cancer T24 cells. Int J Oncol 42: 993-1000, 2013.
31. Wang N, Pan W, Zhu M, et al: Fangchinoline induces autophagic cell death via p53/sestrin2/AMPK signalling in human hepatocellular carcinoma cells. Br J Pharmacol 164: 731-742, 2011.
32. Kang R, Zeh HJ, Lotze MT and Tang D: The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ 18: 571-580, 2011.
K7 modulates Rubicon-mediated inhibition of autophagosome maturation. J Virol 87: 12499-12503, 2013.
34. Hwang S, Maloney NS, Bruinsma MW, et al: Nondegradative role of Atg5-Atg12/ Atg16L1 autophagy protein complex in antiviral activity of interferon gamma. Cell Host Microbe 11: 397-409, 2012.
35. Gao W, Chen Z, Wang W and Stang MT: E1-like activating enzyme Atg7 is preferentially sequestered into p62 aggregates via its interaction with LC3-I. PLoS One 8: e73229, 2013.
36. Ricci MS and Zong WX: Chemotherapeutic approaches for targeting cell death pathways. Oncologist 11: 342-357, 2006.
37. Gosepath EM, Eckstein N, Hamacher A, et al: Acquired cisplatin resistance in the head-neck cancer cell line Cal27 is associated with decreased DKK1 expression and can partially be reversed by overexpression of DKK1. Int J Cancer 123: 2013-2019, 2008.
38. Chang PY, Peng SF, Lee CY, et al: Curcumin-loaded nanoparticles induce apoptotic cell death through regulation of the function of MDR1 and reactive oxygen species in cisplatin-resistant CAR human oral cancer cells. Int J Oncol 43: 1141-1150, 2013.
39. Yang JS, Hour MJ, Huang WW, Lin KL, Kuo SC and Chung JG: MJ-29 inhibits tubulin polymerization, induces mitotic arrest, and triggers apoptosis via cyclin-dependent kinase 1-mediated Bcl-2 phosphorylation in human leukemia U937 cells. J Pharmacol Exp Ther 334: 477-488, 2010.
40. Wang J, Kim TH, Ahn MY, et al: Sirtinol, a class III HDAC inhibitor, induces apoptotic and autophagic cell death in MCF-7 human breast cancer cells. Int J Oncol 41: 1101-1109, 2012.
18beta-glycyrrhetinic acid in PC12 cells via modulation of the PI3K/Akt pathway. J Agric Food Chem 57: 754-761, 2009.
42. Lai KC, Lu CC, Tang YJ, et al: Allyl isothiocyanate inhibits cell metastasis through suppression of the MAPK pathways in epidermal growth factorstimulated HT29 human colorectal adenocarcinoma cells. Oncol Rep 31: 189-196, 2014.
43. Tsai SC, Yang JS, Peng SF, et al: Bufalin increases sensitivity to AKT/mTOR-induced autophagic cell death in SK-HEP-1 human hepatocellular carcinoma cells. Int J Oncol 41: 1431-1442, 2012.
44. Gao M, Yeh PY, Lu YS, et al: OSU-03012, a novel celecoxib derivative, induces reactive oxygen species-related autophagy in hepatocellular carcinoma. Cancer Res 68: 9348-9357, 2008.
45. Liu SY, Lu CL, Chiou CT, et al: Surgical outcomes and prognostic factors of oral cancer associated with betel quid chewing and tobacco smoking in Taiwan. Oral Oncol 46: 276-282, 2010.
46. Sanjiv K, Su TL, Suman S, et al: The novel DNA alkylating agent BO-1090 suppresses the growth of human oral cavity cancer in xenografted and orthotopic mouse models. Int J Cancer 130: 1440-1450, 2012.
47. Yu FS, Yang JS, Yu CS, et al: Safrole induces apoptosis in human oral cancer HSC-3 cells. J Dent Res 90: 168-174, 2011.
48. Lo WL, Kao SY, Chi LY, Wong YK and Chang RC: Outcomes of oral squamous cell carcinoma in Taiwan after surgical therapy: factors affecting survival. J Oral Maxillofac Surg 61: 751-758, 2003.
49. Sun Q, Sakaida T, Yue W, Gollin SM and Yu J: Chemosensitization of head and neck cancer cells by PUMA. Mol Cancer Ther 6: 3180-3188, 2007.
angiogenesis: a potential source for investigational new agents to treat cancer-Part 1. Curr Oncol 13: 14-26, 2006.
51. Riche DM, McEwen CL, Riche KD, et al: Analysis of safety from a human clinical trial with pterostilbene. J Toxicol 2013: 463595, 2013.
Table 1: List of genes with more than 1.5-fold changes in mRNA levels in CAR cells after ANK-199 (50 μM) 24 h-treatment identified by DNA microarray
Accession gene fold
change
XR_042379 LOC401875: hypothetical LOC401875 7.34
NM_198581 ZC3H6: zinc finger CCCH-type containing 6 5.68
NM_004755 RPS6KA5: ribosomal protein S6 kinase, 90kDa, polypeptide 5 3.27
NM_006472 TXNIP: thioredoxin interacting protein 3.00
NM_001506 GPR32: G protein-coupled receptor 32 2.15
NM_018387 STRBP: spermatid perinuclear RNA binding protein 2.04
BC007928 C21orf119: chromosome 21 open reading frame 119 1.93
BC108718 LOC389765: similar to KIF27C 1.88
NM_153335 LYK5: protein kinase LYK5 1.86
ENST0000002177 6
CCT8L1: chaperonin containing TCP1, subunit 8 (theta)-like 1 1.81
NM_018434 RNF130: ring finger protein 130 1.75
NM_181684 KRTAP12-2: keratin associated protein 12-2 1.72
NM_201266 NRP2: neuropilin 2 1.71
NM_181351 NCAM1: neural cell adhesion molecule 1 1.63
NM_004064 CDKN1B: cyclin-dependent kinase inhibitor 1B (p27, Kip1) 1.63
NM_007051 FAF1: Fas (TNFRSF6) associated factor 1 1.62
NM_198993 STAC2: SH3 and cysteine rich domain 2 1.62 NM_018257 PCMTD2: protein-L-isoaspartate (D-aspartate) O-methyltransferase domain containing 2 1.62
NM_000901 NR3C2: nuclear receptor subfamily 3, group C, member 2 1.62
BC093665 FAM92B: family with sequence similarity 92, member B 1.56
NM_016609 SLC22A17: solute carrier family 22, member 17 1.55
NM_181605 KRTAP6-3: keratin associated protein 6-3 1.55
XM_938903 LOC649839: similar to large subunit ribosomal protein L36a 1.53
BC021739 LOC554201: hypothetical LOC554201 1.53
ENST0000032924 4
LOC100132169: similar to hCG1742852 1.50
NM_021109 TMSB4X: thymosin beta 4, X-linked -1.50
NM_019896 POLE4: polymerase (DNA-directed), epsilon 4 (p12 subunit) -1.51
NM_015475 FAM98A: family with sequence similarity 98, member A -1.51
NM_006136 CAPZA2: capping protein (actin filament) muscle Z-line, alpha 2 -1.51
NM_001128619 LUZP6: leucine zipper protein 6 -1.51
NM_014248 RBX1: ring-box 1 -1.52
NM_024755 SLTM: SAFB-like, transcription modulator -1.53
NM_003168 SUPT4H1: suppressor of Ty 4 homolog 1 (S. cerevisiae) -1.53
NM_017892 PRPF40A: PRP40 pre-mRNA processing factor 40 homolog A (S. cerevisiae) -1.53 NM_004776 B4GALT5: UDP-Gal:betaGlcNAc beta 1,4- galactosyltransferase, polypeptide 5 -1.53
NM_002090 CXCL3: chemokine (C-X-C motif) ligand 3 -1.53
NM_000937 POLR2A: polymerase (RNA) II (DNA directed) polypeptide A, 220kDa -1.54
NM_001349 DARS: aspartyl-tRNA synthetase -1.55
NM_003348 UBE2N: ubiquitin-conjugating enzyme E2N (UBC13 homolog, yeast) -1.56
NM_001614 ACTG1: actin, gamma 1 -1.56
NM_053024 PFN2: profilin 2 -1.56
NM_003010 MAP2K4: mitogen-activated protein kinase kinase 4 -1.57
NM_001099771 A26C1B: ANKRD26-like family C, member 1B -1.57
NM_006000 TUBA4A: tubulin, alpha 4a -1.58
NM_133494 NEK7: NIMA (never in mitosis gene a)-related kinase 7 -1.59
NM_173647 RNF149: ring finger protein 149 -1.59
NM_182917 EIF4G1: eukaryotic translation initiation factor 4 gamma, 1 -1.59
NM_002599 PDE2A: phosphodiesterase 2A, cGMP-stimulated -1.62
NM_005066 SFPQ: splicing factor proline/glutamine-rich (polypyrimidine tract binding protein associated) -1.64
NM_001039479 KIAA0317: KIAA0317 -1.64
NM_001127649 PEX26: peroxisomal biogenesis factor 26 -1.64
NM_015153 PHF3: PHD finger protein 3 -1.64
NM_007189 ABCF2: ATP-binding cassette, sub-family F (GCN20), member 2 -1.64
NM_007126 VCP: valosin-containing protein -1.64
NM_012234 RYBP: RING1 and YY1 binding protein -1.65
NR_004845 LOC644936: cytoplasmic beta-actin pseudogene -1.65
NM_014795 ZEB2: zinc finger E-box binding homeobox 2 -1.66
NM_001099692 EIF5AL1: eukaryotic translation initiation factor 5A-like 1 -1.67 NM_138689 PPP1R14B: protein phosphatase 1, regulatory (inhibitor) subunit 14B -1.67 NM_015665 AAAS: achalasia, adrenocortical insufficiency, alacrimia (Allgrove, triple-A) -1.67
NM_001099692 EIF5AL1: eukaryotic translation initiation factor 5A-like 1 -1.68
NM_001099692 EIF5AL1: eukaryotic translation initiation factor 5A-like 1 -1.68
NM_002154 HSPA4: heat shock 70kDa protein 4 -1.68
NM_013451 FER1L3: fer-1-like 3, myoferlin (C. elegans) -1.70
NM_000303 PMM2: phosphomannomutase 2 -1.71
NM_002795 PSMB3: proteasome (prosome, macropain) subunit, beta type, 3 -1.71
NM_001363 DKC1: dyskeratosis congenita 1, dyskerin -1.71
NM_001102 ACTN1: actinin, alpha 1 -1.73
NM_004299 ABCB7: ATP-binding cassette, sub-family B (MDR/TAP), member 7 -1.73
NM_005857 ZMPSTE24: zinc metallopeptidase (STE24 homolog, S. cerevisiae) -1.74
NM_152265 BTF3L4: basic transcription factor 3-like 4 -1.74
NM_020409 MRPL47: mitochondrial ribosomal protein L47 -1.74
NM_006148 LASP1: LIM and SH3 protein 1 -1.76
BC065192 C2orf12: chromosome 2 open reading frame 12 -1.76
NM_001414 EIF2B1: eukaryotic translation initiation factor 2B, subunit 1 alpha, 26kDa -1.78 NM_003222 TFAP2C: transcription factor AP-2 gamma (activating enhancer binding protein 2 gamma) -1.78
NM_007350 PHLDA1: pleckstrin homology-like domain, family A, member 1 -1.79
ENST0000024257 7
NM_032830 CIRH1A: cirrhosis, autosomal recessive 1A (cirhin) -1.79
NM_152265 BTF3L4: basic transcription factor 3-like 4 -1.80
NM_176816 CCDC125: coiled-coil domain containing 125 -1.80
NM_001039690 CTF8: chromosome transmission fidelity factor 8 homolog (S. cerevisiae) -1.83 NM_001127257 SLC39A10: solute carrier family 39 (zinc transporter), member 10 -1.84
XM_001716411 LOC128322: hypothetical LOC128322 -1.86
NM_002370 MAGOH: mago-nashi homolog, proliferation-associated (Drosophila) -1.90
NM_138578 BCL2L1: BCL2-like 1 -1.91
NM_003580 NSMAF: neutral sphingomyelinase (N-SMase) activation associated factor -1.92
NM_015922 NSDHL: NAD(P) dependent steroid dehydrogenase-like -1.92
NM_001797 CDH11: cadherin 11, type 2, OB-cadherin (osteoblast) -1.93
NM_021242 MID1IP1: MID1 interacting protein 1 (gastrulation specific G12 homolog (zebrafish)) -1.93
NM_005968 HNRNPM: heterogeneous nuclear ribonucleoprotein M -1.95
NM_012338 TSPAN12: tetraspanin 12 -1.97
NM_014953 DIS3: DIS3 mitotic control homolog (S. cerevisiae) -2.02
NM_003130 SRI: sorcin -2.11
NM_018993 RIN2: Ras and Rab interactor 2 -2.12
NM_004093 EFNB2: ephrin-B2 -2.13
NM_032256 TMEM117: transmembrane protein 117 -2.14
NM_005415 SLC20A1: solute carrier family 20 (phosphate transporter), member 1 -2.15 NM_017872 THG1L: tRNA-histidine guanylyltransferase 1-like (S. cerevisiae) -2.20
NM_153618 SEMA6D: sema domain, transmembrane domain (TM), and cytoplasmic domain, (semaphorin) 6D -2.27 NM_014604 TAX1BP3: Tax1 (human T-cell leukemia virus type I) binding protein 3 -2.27
NM_033505 SELI: selenoprotein I -2.30
NM_003666 BLZF1: basic leucine zipper nuclear factor 1 -2.35
NM_002714 PPP1R10: protein phosphatase 1, regulatory (inhibitor) subunit 10 -2.50 NM_002714 PPP1R10: protein phosphatase 1, regulatory (inhibitor) subunit 10 -2.50 NM_002714 PPP1R10: protein phosphatase 1, regulatory (inhibitor) subunit 10 -2.50
NR_003003 SCARNA17: small Cajal body-specific RNA 17 -2.51
NR_002738 SNORD57: small nucleolar RNA, C/D box 57 -2.57
NM_006080 SEMA3A: sema domain, immunoglobulin domain (Ig), short basic domain, secreted, (semaphorin) 3A
-2.57
NM_009587 LGALS9: lectin, galactoside-binding, soluble, 9 -2.65
NM_003234 TFRC: transferrin receptor (p90, CD71) -2.85
NM_138966 NETO1: neuropilin (NRP) and tolloid (TLL)-like 1 -2.97
NM_001098272 HMGCS1: 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 1 (soluble) -2.97
NM_006350 FST: follistatin -4.21
NM_024090 ELOVL6: ELOVL family member 6, elongation of long chain fatty acids (FEN1/Elo2, SUR4/Elo3-like, yeast)
-4.46
NM_005328 HAS2: hyaluronan synthase 2 -5.00
NM_001753 CAV1: caveolin 1, caveolae protein, 22kDa -5.04
Table 2: Gene to GO Biological Process test for over-representation (ANK-199 to Control)
Term Count % P Value
GO:0044087~regulation of cellular component biogenesis 9 6.382979 8.59E-06
GO:0030036~actin cytoskeleton organization 10 7.092199 3.75E-05
GO:0032956~regulation of actin cytoskeleton organization 7 4.964539 4.43E-05
GO:0043254~regulation of protein complex assembly 7 4.964539 4.72E-05
GO:0032970~regulation of actin filament-based process 7 4.964539 5.34E-05
GO:0051493~regulation of cytoskeleton organization 8 5.673759 5.75E-05
GO:0030029~actin filament-based process 10 7.092199 6.17E-05
GO:0007010~cytoskeleton organization 13 9.219858 7.13E-05
GO:0008064~regulation of actin polymerization or depolymerization 6 4.255319 7.77E-05
Table 3: Gene to GO Molecular Function test for over-representation (ANK-199 to Control)
Term Count % P Value
GO:0003723~RNA binding 21 14.89362 1.54E-07
GO:0000166~nucleotide binding 33 23.40426 6.28E-05
GO:0008092~cytoskeletal protein binding 11 7.801418 0.00346
GO:0032553~ribonucleotide binding 24 17.02128 0.005183
GO:0032555~purine ribonucleotide binding 24 17.02128 0.005183
GO:0017076~purine nucleotide binding 24 17.02128 0.008794
GO:0003779~actin binding 8 5.673759 0.009366
GO:0019900~kinase binding 6 4.255319 0.009636
GO:0047485~protein N-terminus binding 4 2.836879 0.016454
Figure legends
Figure 1. Chemical structures of (A) pterostilbene and (B) ANK-199.
Figure 2. Effect of ANK-199 on CAR cell viability and morphological examination. CAR cells were treated with or without various concentrations of ANK-199 for 24, 28 and 72 h. (A) Cell viability was measured by MTT assay. Data are presented as the mean ± S.E.M (n =3). ***p < 0.05 vs vehicle control. (B) Cells were photographed with a phase-contrast microscope.
Figure 3. Effect of ANK-199 on normal oral cells. (A) Normal human gingival fibroblasts cells (HGF) and (B) normal human oral keratinocyte cells (OK) cells after exposure to various concentrations of ANK-199 for 72 h were determined by MTT assay. The data shown represent the mean ±S.E.M . (n =3).
Figure 4. Effect of ANK-199 on autophagic death in CAR cells. Cells in the presence and absence of ANK-199 challenge for 48 h were harvested to investigate. (A) The autophagolysosome marker was probed by MDC. (B) AO staining was used to detect AVO. (C) LC3B expression was determined by Premo Autophagy Sensor kit. (D) DNA condensation (an apoptotic characteristic) was tested utilizing DAPI staining.
Figure 5. Effect of ANK-199 on autophagy-regulated signaling in CAR cells. Immunoblot analysis of the molecules of vehicle control or ANK-199 treatment at 50 and 75 μM of CAR cells for 48 h showed autophagy-modulated signal molecules (Atg5, Atg7, Atg12, Atg14, Atg16 L1, Beclin 1, PI3K classIII, Rubicon, LC-3) as described in Materials and Methods. The anti-β-cctin mAb was used as an internal
control for loading.
Figure 6. Effect of ANK-199 on autophagy-related gene expression in CAR cells. Total RNA was extracted from CAR cells after ANK-199 treatment, and real-time PCR was performed for different specific primers, including (A) Atg7, (B) Atg12, (C)
Beclin-1 and (D) LC3II gene levels. GAPDH is as an internal control gene. The results
are given as mean ± S.E.M. (n = 3), *p < 0.05 and ***p < 0.001 vs vehicle control.
Figure 7. Effect of 3-MA on autophagic death in ANK-199-treated CAR cells. Cells were pretreated with 10 mM 3-MA and then exposed to 50 or 75 μM of ANK-199 for 48 h. (A) The autophagic vacuole formation was assessed using MDC staining, and (B) MTT assay was applied to estimate cell viability. All values are expressed as mean ± S.E.M. (n = 3). ***p < 0.001 vs vehicle control or ANK-199 treatment groups.
Figure 8. Proposed schematic molecular signaling for ANK-199-triggered autophagy in cisplatin-resistant human oral cancer CAR cells.