A Computational Model of the Canine
Pulmonary Veins Sleeves
Chu-Pin Lo
1, Tzyy-Leng Horng
2∗, Hsiang-Ning Luk
3,
Hui-Chun Tien
1, Juan-Ming Yuan
1, and Daniel Lee
4 1Department of Applied Mathematics, Providence University, Taichung, Taiwan2Department of Applied Mathematics, Feng Chia University, Taichung, Taiwan 3Department of Anesthesiology, Taichung Veterans General Hospital, Taichung, Taiwan
4Department of Mathematics, Tunghai University, Taichung, Taiwan
July 4, 2006
Abstract
The cellular electrophysiological properties of the pulmonary vein sleeves (PVS), the intersection between pulmonary veins and left atrium, were re-ported few decades ago. However, the clinical significance of pulmonary vein sleeves was not found until recently. In patients with atrial fibrilla-tion, it was found that ectopic foci existed in the pulmonary vein sleeves. Therefore, electrophysiological studies on PVS have drawn a lot of at-tention recently. For example, in the experiments of dogs and rabbits, besides the fast response type of action potential, various pacemaker-like automaticities and triggered activities, e.g., early afterdepolarizations (EAD), delayed afterdepolarizations (DAD), have been demonstrated in the PVS under physiological or pathophysiological situations (see [Hocini et al., 2002], [Ehrlich et al., 2003], [Hojo et al., 2003], [Wang et al., 2003, 2005], [Po et al., 2005], [Chen et al., 2000, 2001, 2002-a, 2002-b, 2002-c, 2003-a, 2003-b, 2003-c, 2004-a, 2004-b]).
In this study, we have developed a mathematical model which fits well the experiments for the canine PVS in [44]. Using this model, besides the normal response of the PVS cells we have also successfully reproduced the experimentally observed frequency responses and ischemia/hyperkalemia phenomena.
Keywords :Pulmonary vein sleeves, Action potential
1
Introduction
Pulmonary veins (PV) are the great vessels connected between lung and left atrium and transport oxygenated blood to the heart. There are 4 pulmonary
∗Corresponding author. Tel: +886-4-24517250 ext. 5126. E-mail: [email protected]
veins in human beings. At the intersection between pulmonary veins and left atrium, there is a short segment covered with striated cardiac muscle (not smooth muscle) and called pulmonary vein sleeve (PVS). Complex anatomical architecture of the mammalian PVS, due to the intermingled circular and lon-gitudinal muscular bundles, might contribute to formation of ectopic focus/foci during atrial tachyarrhythmias. Any changes in the action potentials parame-ters of PV sleeves myocytes are prone to cause abnormal impulse formation and conduction. Arrhythmias can be generated by the mechanisms such as abnor-mal automaticity, triggered activity and reentry. On the cellular mechanisms responsible for the normal and abnormal electrophysiological properties of PV sleeves myocytes, various transmembrane ionic currents and intracellular ionic activities are involved.
The role of pulmonary vein (PV) sleeves in humans is not clear until recently. It has been demonstrated in a pioneer work from Haissaguerre et al. [19] that ectopic focus/foci originated in pulmonary vein (PV) have been mapped and ablated in patients with focal atrial fibrillation. Atrial tachyarrhythmias could therefore be successfully terminated and sinus rhythm could be resumed. A great attention on the role of PV in initiating and sustaining focal atrial fibril-lation therefore has been drawn.
In literature, fundamental characterization of PV sleeves has been docu-mented in rodents ([42], [Cheung, 1981-a], [Cheung, 1981-b]), dogs and rabbits ([2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [16], [23], [24], [39], [44], [45]). In addition to fast-response action potentials, spontaneous activities and triggered activities, such as early afterdepolarizations (EAD) and delayed afterdepolar-izations (DAD) have also been shown.
In Chen et al. series of papers , it has been reported that a high incidence of pacemaker cells activity, EAD, DAD, and so-called high frequency irregular rhythms of the multicellular preparations or isolated single myocytes from PV sleeves from normal healthy dogs and rabbits. Based on the cellular electrophys-iological data, Chen et al. therefore made a conclusion that PV sleeves contained multiple types of cardiomyocytes with distinct electrophysiological characteris-tics and implied a role of PV in initiating atrial fibrillation. However, the other five international independent research groups have presented a contradictory conclusion. Namely, no abnormal automaticity or triggered activity was ever found in dogs and rabbits pulmonary veins sleeves under normal experimental conditions. Therefore, this controversy regarding the important electrophysio-logical properties of pulmonary veins sleeves need a thorough re-examination in details.
Since the pioneering work of Hodgkin and Huxley [ ] in describing quantita-tively the ionic currents of the squid giant axon, mathematical models have pro-vided one important and useful approach to study the cardiac cells. Mathemat-ical algorithms and equations are derived to describe versatile action potentials configurations, transmembrane ionic currents and channels, pumps, exchangers and intracellular ions activities under normal and abnormal conditions. In lit-erature, there have appeared many mathematical models for the various types of cells in heart of different species. However there are no suitable
computa-tional models for the PV sleeves until now. Since the important role of the PV sleeves and the debate mentioned above, it is urgent and valuable to build up a mathematical model for the PV sleeves.
In the present paper, based on the experimental data in [44], we have estab-lished a mathematical model to explore various canine PVS electrophysiological characteristics, under both normal and pathological conditions.
2
Materials and Methods
2.1
Establishment of the canine PV sleeves mathematical
model
In the literature, there are no specific mathematical models for the PV. We will modify the mathematical model for the canine right atrium in [27] and [40] to serve our purpose. As stated in [16] and [29], there are no differences in kinetic or voltage-dependent properties of currents between the right atrium (RA), left atrium (LA), and PV. The only difference is the magnitude of some ion current densities, which then causes characteristic changes of the action potential. In summary, LA myocytes have larger rapid delayed rectifier current
IKr than do RA. Therefore, the action potential duration (APD) is shorter in
the LA versus RA and the effective refractory period is also shorter in the LA compared with RA, resulting in faster repolarization in LA. On the other hand, PV has greater rapid and slow delayed rectifier currents, IKr, IKs, respectively
and smaller inward rectifier current IK1, transient outward K+current Ito, and
L-type Ca2+ current I
Ca. Hence, PV has shorter APD, more positive resting
membrane potential (RMP), and slower phase 0 upstroke velocity, compared to LA.
Since the differences of electrophysiological properties between RA and PV are not essential from the kinetical viewpoint as mentioned above, we will modify the model in [27], [40] by mainly adjusting the current densities according to the experimental data in [44].
2.2
Parameters of the model
All of the model equations of this study are the same as in [40], except for some revisions in the appendix. All of the parameters in our model are chosen in Table 1 of [40] except those indicated belows. The initial data are taken from Table 1 of [27]. Since the experimental samples in [44] are superfused with the normal Tyrode solution composed of (mM), N aCl 130, KCl 4, CaCl21.8, M gCl20.5,
N aHCO3 12, and glocose 5, the extracellular ionic concentrations are fixed as:
potassium concentration [K+]
o= 4 mM, chloride concentration [Cl−]o= 138.6
mM, sodium concentration [N a+]
o= 142 mM, calcium concentration [Ca2+]o=
1.8 mM. As explained in [27], the maximal conductance of the transient outward
K+ current, g
In order to fit the experimental results in [44], we have reasonably revised some ionic current conductances in Table 1 of [40]; see the section “Results” below.
2.3
ODE numerical method
Here we used MATLAB function ODE15S as the ODE solver. ODE15S is a stiff multistep variable order solver based on the numerical differentiation formulas (NDFs). Optionally, it uses the backward differentiation formulas (BDFs, also known as Gear’s method) that are usually less efficient. It turns out to be very cost efficient for our current stiff problem.
3
Results and Discussion
We used the current-clamp mode with stimulus current intensity Istim= −2900
pA, duration=2 ms to record the action potential and various current densities.
To describe the action potential more precisely, usually the following parame-ters will be measured: the action potential amplitude(AP A), resting membrane potential(RM P ), action potential duration (AP D) at 50% and 90% of repo-larization (AP D50 and AP D90 , respectively), and the maximal velocity of
depolarization of phase 0 (Vmax).
3.1
Frequency response of the action potential
Using our canine PV model with rapid delayed rectifier current conductance and inward rectifier current conductance gKr= 0.055872, gK1= 0.06 (nS/pF),
respectively, we repeated the frequency response experiment as in [44]. The computer simulation results under the pacing rates (Hz): 0.05, 0.1, 0.5, 1, 2, 3, 4, are shown in Figure 1, 2, 3, and Table 1. We then conclude that the action potentials become shorter (AP D &), depolarized (RM P %) and depressed (Vmax&) as the frequency increases, which are consistent with the experimental
results in [44](see Fig. 3 therein).
3.2
Ischemia/hyperkalemia
Myocardial ischemia results from a withdrawal of oxygen from myocardial tis-sue (due to inefficient or absent perfusion), resulting in disturbances to aerobic respiration and ATP production. Alterations in intracellular ATP can alter the activty of membrane pumps, and thus the distribution of critical ions, e.g.,
N a+and K+. As the intracellular stores of ATP diminish due to reduced aerobic
respiration, N a+ /K+ pumps responsible for ion distribution also demonstrate
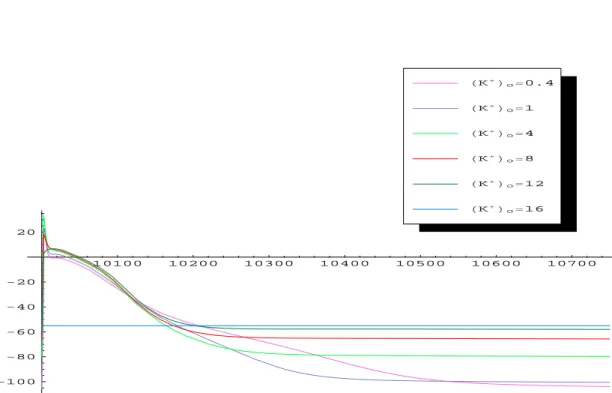
reduced activity. Though normally this pump acts to relocate sodium out of the cell and potassium into the cell, a lethargic pump performs this process ineffi-ciently. Thus, there is an extracellular accumulation of the potassium, referred to as “hyperkalemia”. We mimicked the hyperkalemia situation by varying
the extracellular potassium concentration as in [44]. The obtained results are shown in Figures 4, 5, 6, 7 and Table 2. Clearly, with the increased extracellular potassium, RMP becomes more positive, increasing sodium channel inactivation and reducing the inward sodium current. This dominating effect is somewhat mitigated by the increased RMP being closer to the action potential thresh-old. Thus, moderate increases (0.4 mM to 4 mM) in potassium may actually increase upstroke velocity Vmax (this is termed “superconduction”), while large
increases in extracellular potassium concentrations (>4mM) begin to inactivate the sodium current and decrease upstroke velocity. In summary, our simu-lation results have demonstrated the classical characteristics of ischemic cells: depolarization of resting potential, reduced action potential upstroke, reduced conduction velocity, and reduced action potential duration, which were consis-tent with the experimental results in [44] (see Fig. 4 therein).
4
Conclusion
In this study, we have successfully reproduced the key characteristics of the ex-periments in [44] about frequency response and varying extracellular potossium concentration. As a robust computational model, this model should be tested by more pathophysiological experiments, e.g., ischemia/hypoxia, drugs effect ... Maybe new currents will be included to fit new experimental data, e.g., ATP related potassium current... In particular, the reasons leading to the automatic-ity phenomena and triggered waves, the main conflict in the literature, should be explored in the future works.
50 100 150 200 250 -80 -60 -40 -20 20 4Hz 3 Hz 2 Hz 1 Hz 0.5 Hz 0.1 Hz 0.05 Hz
Fig.1 Frequency response of action potential under different pacing rates
1 2 3 4 Hz 195 200 205 210 APD90 HmsL 1 2 3 4 Hz 90 95 100 105 110 115 APA HmVL 1 2 3 4 Hz -80 -78 -76 -74 -72 RMPHmVL 1 2 3 4 Hz 42.5 47.5 50 52.5 55 57.5 V max HVsL
Fig. 2 Parameters of action potentials under different pacing rates 1 2 3 4 Hz 20 40 60 80 100 V max H%L
Fig. 3 Vmax versus frequency (normalized)
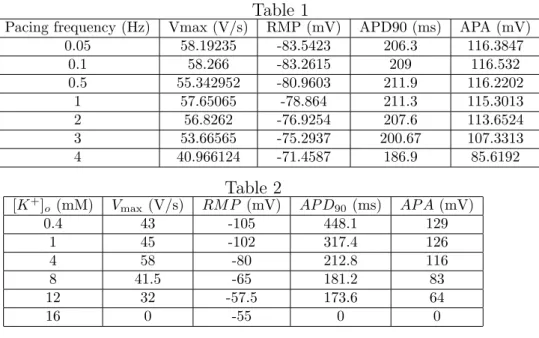
Table 1
Pacing frequency (Hz) Vmax (V/s) RMP (mV) APD90 (ms) APA (mV)
0.05 58.19235 -83.5423 206.3 116.3847 0.1 58.266 -83.2615 209 116.532 0.5 55.342952 -80.9603 211.9 116.2202 1 57.65065 -78.864 211.3 115.3013 2 56.8262 -76.9254 207.6 113.6524 3 53.66565 -75.2937 200.67 107.3313 4 40.966124 -71.4587 186.9 85.6192
Table 2
[K+] o (mM) Vmax (V/s) RM P (mV) AP D90(ms) AP A (mV) 0.4 43 -105 448.1 129 1 45 -102 317.4 126 4 58 -80 212.8 116 8 41.5 -65 181.2 83 12 32 -57.5 173.6 64 16 0 -55 0 010100 10200 10300 10400 10500 10600 10700 -100 -80 -60 -40 -20 20 HK+Lo=16 HK+Lo=12 HK+Lo=8 HK+Lo=4 HK+Lo=1 HK+Lo=0.4
2.5 5 7.5 10 12.5 15 @K +D o HmML 100 200 300 400 APD90 HmsL
2.5 5 7.5 10 12.5 15 @K +D o HmML -100 -80 -70 -60 RMP HmvL Fig. 6 RMP versus [K+] o
2.5 5 7.5 10 12.5 15 @K +D o HmML 20 40 60 80 100 120 APA HmvL
Fig. 7 APA versus [K+] o
2.5 5 7.5 10 12.5 15 @K +D o HmML 10 20 30 40 50 Vmax HvsL
Fig. 8 Vmax versus [K+]o
Appendix
In this study, all of the model equations are the same as in [40], except the followings. The equation labelled “rA*” in this appendix means the revision of the equation “A*” in [40]. Note that the revised equations (rA2), (rA3), (rA4), (rA5), (rA20), (rA29), (rA30), are due to the explanations in [27] or unit consistency. For (rA74),(rA75),(rA76), refer to [26]. The other errors can be easily seen from the basic conservation principle and the unit consistency.
dV
dt = −Iion− Istim
Cm (rA1)
Iion= IN a+ IK1+ Ito+ IKur,d+ IKr+ IKs+ ICa+ ICl,Ca+ Ip,Ca+ IN aCa+ IN aK+ Ib,N a+
Ib,Ca+ Ib,Cl, where Ib,Cl= 0.0018(V − ECl) (see (2), (4) of [27])
(rA2)
d[N a+]
i
dt =
Cm(−3IN aK−3IN aCa−Ib,N a−IN a+CTN aCl)
F Vi ,
d[K+]
i
dt =
Cm(2IN aK− IK1− Ito− IKur,d− IKr− IKs) − Istim
F Vi (rA4) d[Cl−] i dt = Cm(ICl,Ca+ Ib,Cl+ CTN aCl) F Vi (rA5) d[Ca2+] i dt =
Cm(2IN aCa−Ip,Ca−ICa−Ib,Ca)
2F Vi +
Vup(Iup,leak−Iup)+VrelIrel
Vi −d[Ca2+]T rpn dt − d[Ca2+] Cmdn dt (rA6) d[Ca2+] rel dt = Itr− Irel− d[Ca2+] Csqn dt (rA8) IK1= r [K+]o 5.4 gK1(V − EK) 1 + exp[0.07(V + 80)] (rA19) Ito= 0.04956Oa3Oi(V − EK) (rA20) IKur,d= 0.75(0.00855 + 0.0779 1 + exp(V +11 −16 ) )u3 aui(V − EK) (rA29) gKur,d= 0.75(0.00855 + 0.0779 1 + exp(V +11 −16 ) ) (rA30) IKr= r [K+]o 5.4 gKrxr(0.07 + 0.58 1 + exp[(V + 15)/22.4])(V − EK) (rA39) 1 [Cmdn]max d[Ca2+] Cmdn dt = 200[Ca 2+] i(1 − [Ca 2+] Cmdn [Cmdn]max ) − 0.476 [Ca2+] Cmdn [Cmdn]max (rA74) 1 [T rpn]max d[Ca2+] T rpn dt = 784[Ca 2+] i(1 −[Ca 2+] T rpn [T rpn]max ) − 0.392 [Ca2+] T rpn [T rpn]max (rA75) 1 [Csqn]max d[Ca2+] Csqn dt = 0.48[Ca 2+] rel(1 −[Ca 2+] Csqn [Csqn]max ) − 0.4 [Ca2+] Csqn [Csqn]max (rA76)
Acknowledgements
The study was supported, in part, by the grants from Providence Univer-sity and Taichung Veterans General Hospital (PU-93-11100-5162-006-007) and Taiwan National Science Council (NSC 93-2115-M-126-002- and NSC 94-2115-M-126-002-). Also, we thank the support of the National Center for Theoretical Science (NCTS) of Taiwan.
References
[1] Carmeliet E, ”Calcium channel-antagonists and the cardiovascular system”,
Acta Cardiol, 41:133-46, 1986
[2] Chen SA, Chen YJ, Yeh HI, Tai CT, Chen YC, Lin CI, ”Pathophysiol-ogy of the pulmonary vein as an atrial fibrillation initiator”, Pacing Clin
Electrophysion, 26:1576-82, 2003-a
[3] Chen SA, Tai CT, Yeh HI, Chen YJ, Lin CI, ”Controversies in the mech-anisms and ablation of pulmonary vein atrial fibrillation”, Pacing Clin
Electrophysiol, 26:1301-7, 2003-b
[4] Chen YC, Chen SA, Chen YJ, Chang MS, Chan P, Lin CI, ”Effects of thyroid hormone on the arrhythmogenic activity of pulmonary vein car-diomyocytes”, J Am Coll Cardiol, 39:366-72, 2002-a
[5] Chen YC, Chen SA, Chen YJ, Tai CT, Chan P, Lin CI, ”Effect of ethanol on the electrophysiological characteristics of pulmonary vein cardiomyocytes”,
Eur J Pharmacol, 483:215-22, 2004-a
[6] Chen YC, Chen SA, Chen YJ, Tai CT, Chen P, Lin CI, ”T-type calcium current in electrical activity of cardiomyocytes isolated from rabbit pul-monary vein”, J Cardiovasc Electrophysiol, 15:567-71, 2004-b
[7] Chen YJ, Chen SA, Chang MS, Lin CI, ”Arrhythmogenic activity of cardiac muscle in pulmonary veins of the dog: implication for the gensesis of atrial fibrillation”, Cardiovasc Res, 48:265-73, 2000
[8] Chen YJ, Chen SA, Chen YC et al., ”Effects of rapid atrial pacing on the ar-rhythmogenic activity of single cardiomyocytes from pulmonary veins: im-plication in initiation of atrial fibrillation”, Circulation, 104:2849-54, 2001 [9] Chen YJ, Chen SA, Chen YC, Yeh HI, Chang MS, Lin CI, ”Electrophysiol-ogy of single cardiomyocytes isolated from rabbit pulmonary veins: impli-cation in initiation of focal atrial fibrillation”, Basic Res Cardiol, 97:26-34, 2002-b
[10] Chen YJ, Chen YC, Chan P, Lin CI, Chen SA, ”Temperature regulates the arrhythmogenic activity of pulmonary vein cardiomyocytes”, J Biomed
Sci, 10:535-43, 2003-c
[11] Chen YJ, Chen YC, Yeh HI, Lin CI, Chen SA, ”Electrophysiology and arrhythmogenic activity of single cardiomyocytes from canine superior vena cava”, Circulation, 105:2679-85, 2002-c
[12] Chen DW, ”Electrical activity of the pulmonary vein and its interaction with the right atrium in the guinea-pig”, J Physiol, 314:455-56,1981 (a) [13] Chen DW, ”Pulmonary vein as an ectopic focus in digitalis-induced
[14] Chugh A, Ozaydin M, Scharf C et al, ”Mechanism of immediate recur-rences of atrial fibrillation after restoration of sinus rhythm”, Pacing Clin
Electrophysiol, 27:77-82, 2004
[15] De Bakker JM, Ho SY, Hocini M, ”Basic and clinical electrophysiology of pulmonary vein ectopy”, Cardiovasc Res, 54:287-94, 2002
[16] Ehrlich JR, Cha TJ, Zhang L, Charties D, Melnyk P, Hohnlose SH, Nattel S, ”Cellular electrophysiology of canine pulmonary vein cardiomyocytes: action potential and ionic current properties”, J Physiol, 551:801-13, 2003 [17] Fenton FH, Cherry EM, Hastings HM, Evans SJ, ”Multiple mechanisms of spiral wave breakup in a model of cardiac electrical activity”, Chaos, 12:852-892, 2002
[18] Garfinkel A, Kim H, Voroshilovsky O, et al., ”Preventing ventricular fibril-lation by flattening cardiac restitution”, Proc Natl Acad Sci USA, 97:6061-6, 2000
[19] Haissaguerre M, Jais P, Shah DC et al, ”Spontaneous initiation of atrial fibrillation by ectopic beats originating in the pulmonary veins”, N Engl J
Med, 339:659-66,1998
[20] He B, Modeling and imaging of bioelectrical activity :principles and appli-cations, Kluwer Academic/Plenum Publishers, 2004.
[21] Hille B, Ion channels of excitable membranes, 3rd edition. Sinauer Asso-ciates, Inc., 2001.
[22] Hiraoka M, Ikeda K, Sano T, ”The mechanism of barium0induced auto-maticity in ventricular muscle fibers”, Adv Myocardiol, 1:255-66, 1980 [23] Hocini M, Ho SY, Kawara T et al, ”Electrical conduction in canine
pul-monary veins: electrophysiological and anatomic correlation”, Circulation, 105:2442-8, 2002
[24] Honjo H, Boyett MR, Niwa R, lnada S, Yamamoto M, Mitsui K, Horiuchi T, Shibata N, Kamiya K, Kodama I.” Pacing-induced spontaneous activity in myocardial sleeves of pulmonary veins after treatment with-ryanodine”,
Circulation, 107:1937-43, 2003
[25] Imoto Y, Ehara T, Matsuura H, ”Voltage- and time-dependent block of iK1 underlying Ba+-induced ventricular automaticity”, Am J Physiol,
252:H325-33, 1987
[26] Keener J and Sneyd J., Mathematical Physiology, in “interdisciplinary Ap-plied Mathematics 8” (J.E. Marsden, L. Sirovich, and S. Wiggins editors), New York, Spinger-Verlag, 1998.
[27] Kneller J, Ramirez RJ, Chartier D, Courtemanche M, and Nattel S, ”Time-dependent transients in an ionically based mathematical model of the ca-nine atrial action potential”, Am J Physiol Heart Circ Physiol, 282: H1437-H1451, 2002
[28] Kumagai K, Ogawa M, Noguchi H, et al, ”Electrophysiological properties of pulmonary veins assessed using a multielectrode basket catheter”, J Am
Coll Cardiol, 43:2281-9, 2004
[29] Li D, Zhang L, Kneller J, and Nattel S, ”Potential ionic mechanism for repolarization differences between canine right and left atrium”, Circ Res, 88:1168-1175, 2001
[30] Luo CH, Rudy Y, ”A dynamic model of the cardiac ventricular action potential. I. Simulations of ionic currents and concentration changes”, Circ
Res, 74:1071-96, 1994
[31] Luo CH, Rudy Y, ”A dynamic model of the cardiac ventricular action potential. II. Afterdepolarizations, triggered activity, and potentiation”,
Circ Res, 74:1097-133, 1994
[32] Mansour M, Ruskin J, Keane D, ”Initiation of atrial fibrillation by ectopic beats originating from the ostium of the inferior vena cava”, J Cardiovasc
Electrophysiol, 13:1292-5, 2002
[33] Morillo CA, Klein GJ, Jones DL, Guiraudon GM. ”Chronic rapid atrial pacing:structural, functional, and electrophysiologic characteristics of a new model of sustained atrial fibrillation”, Circulation, 91:1588-1595,1995 [34] Nattel S, Li D, Yue L, ”Basic mechanisms of atrial fibrillation—very new
insights into very old ideas”, Annu Rev Physiol, 62:51-77, 2000
[35] Nattel S, Li D. ”lonic remodeling in the heart: pathophysiological signifi-cance and new therapeutic opportunities for atrial fibrillation”, Circ Res, 87:440-7, 2000
[36] Omichi C, Chou CC, Lee MH, Chang CM, Lai AC, Hayashi H, Zhou S, Milyauchi Y, Okuyama Y, Hamabe A, Hwang C, Fishbein MC, Lin SF, Karagueuzian HS, Chen PS. ”Demonstration of electrical and anatomic connections between Marshall bundles and left atrium in dogs: Implications on the generation of P waves on surface Electrocardiogram”, J Cardiovasc
Electrophysiol, 13:1283-91, 2002
[37] Oral H, Knight BP, Tada H, Ozaydin M, Chugh A, Hassan S, Scharf C, Lin SW, Greenstein R, Pelosi F Jr, Strickberger SA, Moragy F, ”Pulmonary vein isolation for paroxysmal and persistent atrial fibrillation”, Circulation, 105:1077-81, 2002
[38] Oral H, Ozaydin M, Tada H, et al, ”Mechanistic significance of intermittent pulmonary vein tachycardia in patients with atrial fibrillation”, J
Cardio-vasc Electrophysiol, 13:645-50, 2002
[39] Po SS, Li Y, Tang D, Liu H, Geng N, Jackman WM, Scherlag B, Lazzara R, Patterson E, ”Rapid and stable re-entry within the pulmonary vein as a mechanism initiating paroxysmal atrial fibrillation”, Journal of the
American College of Cardiology, 45:1871-7, 2005
[40] Ramirez RJ, Nattel S, and Courtemanche M, ”Mathematical analysis of canine atrial action potentials: rate, regional factors, and electrical remod-eling”, Am J Physiol Heart Circ Physiol, 279: H1767-H1785, 2000
[41] Schram G, Pourrier M, Melnyk P, Nattel S. ”Differential distribution of cardiac ion channel expression as a basis for regional speciallization in elec-trical function”, Circ Res, 90:939-50, 2002
[42] Tasaki H. ”Electrophysiological study of the striated muscle cells of the extrapulmonary vein of the guinea-pig”, Jpn Circ J, 33:1087-98, 1969 [43] Tsai CF, Tai CT, Chen SA, ” Pulmonary vein ablation: role in preventing
atrial fibrillation”, Curr Opin Cardiol, 18:39-46, 2003
[44] Wang TM, Chiang CE, Sheu JR, Tsou CH, Chang HM, Luk HN, ”Homoge-nous distribution of fast response action potentials in canine pulmonary vein sleeves: a contradictory report”, Int J Cardiol, 89:187-95, 2003 [45] Wang TM, Luk HN, Sheu JR, Wu HP, Chiang CE, ”Inducibility of
ab-normal automaticity and triggered activity in myocardial sleeves of canine polmonary veins”, Int J Cardiol, 104:59-66, 2005
[46] Wang YG, H¨user J, Blatter LA, Lipsius SL, ”Withdrawal of acetylcholine elicits Ca2+-induced delayed afterdepolarizations in cat atrial myocytes”,
Circulation, 96:1275-1281, 1997
[47] Weiss JN, Garfinkel A, Karagueuzian HS, Qu Z, Chen PS, ”Chaos and the transition to ventricular fibrillation: a new approach to antiarrhythmic drug evaluation”, Circulation, 99:2819-2826,1999
[48] Weiss JN, Chen PS, Qu Z, Karagueuzian HS, Lin SF, Garfinkel A, ”trical restitution and cardiac fibrillation”, Journal of Cardiovascular
Elec-trophysiology, 13:292-5, 2002
[49] Wijffels MC, Kirchhof CJ, Dorland R, Allessie MA, ”Atrial fibrillation begets atrial fibrillation, A study in awake chronically instrumented goats”,
Circulation, 92:1954-68,1995
[50] Wu TJ, Ong JJ, Chang CM, Doshi RN, Yashima M, Huang HL, Fish-bein MC, Ting CT, Karagueuzian HS, Cheng PS. ”Pulmonary veins and ligament of marshall as sources of rapid activations in a canine model of sustained atrial fibrillation”, Circulation, 103;1157-1163, 2001.
![Fig. 5 APD 90 versus [K + ] o](https://thumb-ap.123doks.com/thumbv2/9libinfo/7492367.115351/9.892.208.829.220.660/fig-apd-versus-k-o.webp)
![Fig. 7 APA versus [K + ] o](https://thumb-ap.123doks.com/thumbv2/9libinfo/7492367.115351/11.892.205.812.208.641/fig-apa-versus-k-o.webp)
![Fig. 8 V max versus [K + ] o](https://thumb-ap.123doks.com/thumbv2/9libinfo/7492367.115351/12.892.207.827.207.686/fig-v-max-versus-k-o.webp)
