國立高雄大學電機工程學系(研究所)
碩(博)士論文
多階層基因集遺傳演算法
Hierarchical Gene-Set Genetic Algorithms for Optimization
研究生:吳明泰撰
指導教授:洪宗貝
Hierarchical Gene-Set Genetic Algorithms for
Optimization
Advisor: Dr.(Professor) Tzung-Pei Hong Institute of Electrical Engineering National University of Kaohsiung
Student: Min-Thai Wu Institute of Electrical Engineering National University of Kaohsiung
ABSTRACT
In this paper, gene sets, instead of individual genes, are used in the genetic process to speed up convergence. A gene-set mutation operator is proposed, which can make several neighboring genes to simultaneously mutate. A gene-set crossover operator is also designed to choose the crossover points at the boundary of gene sets. The proposed gene-set mutation and crossover operators will cause a larger diversity than the conventional ones. A hierarchical gene-set genetic algorithm is then proposed, which uses adjustable gene-set lengths to find final solutions. Different phases of populations use different gene-set lengths to perform the genetic operations. The gene-set length is shortened in half in each phase until the length is 1. In addition, effectively avoiding local optimal trapping has been very important to GAs. One modified gene-set genetic algorithm is thus proposed to further help the escape of local optimums. An escape operation, as well as the gene-set mutation operation, is designed for gene-set genetic algorithms to increase the probability of finding global optima. The property that a longer gene set will cause a larger diversity is formally shown. The modified gene-set genetic algorithm can consider both the escape from local optima and the search for global optima. Since the modified approach will generate a smaller number of offspring for a larger gene-set size, it is extended by using dynamic mutation rates to increase the number of offspring when the gene-set size is large. Experiments on three problems are also made to show the effectiveness of the proposed gene-set genetic algorithms.
Keywords: genetic algorithm, chromosome, gene, gene set, crossover, mutation, escape operation.
多階層基因集遺傳演算法
指導教授:洪宗貝 博士(教授) 國立高雄大學電機工程研究所 學生:吳明泰 國立高雄大學電機工程研究所 摘要 在本論文中,在運行基因演算法的過程中,使用基因集來取代傳統單一集因 的運作來加速整個演算法的表現及效能。使用基因集的突變運算被提出,它可以 使得多數個臨近的基因在同一個時間點上發生突變,另一個使用基因集的交配運 算同時被提出,在選擇交配的位置是,取決於基因集為單位。被提出的突變和交 配的運算可以比原先使用單一基因為單位的運算得到更大的差異性。結合基因集 的突變和交配運算,一個多階層遺傳演算法被提出,它使用一個可以被調整的基 因集長度來搜尋最後的結果。在不同的時期中,族群使用不同的基因集長度去執 行基因演算法。經過每一個時期,基因集長度縮短為原先的一半,直到基因集長 度還原為 1 為止。有效的避開區域最佳值對於基因演算法是非常重要的,在另外 修正的兩個基因集演算法中,一個被設計的跳脫運算被提出,它類似一種突變的 效果,可以有效提高演算法搜尋到全域最佳值的機率,在本論文中,將會推論在 較長的基因集運算可以對於子代造成較大的差異性變化,跳脫運算便是基於此特 徵所設計。跳脫運算可以同時減少運算時間及提高搜尋到較佳的解的機率。當基 因集長度較長的時候,所產生的後代數目會有比較少的問題,在其中之一個修正 的基因集演算法中,當基因集長度較長的時候,我們使用了一個動態調整突變率 的方式來提高後代的數目,它可以幫助演算法得到較好的解答,在實驗中,使用 了三個不同特性的方程式來呈現多階層基因集遺傳法的效能和表現。 關鍵字:遺傳演算法、染色體、基因、基因集、突變運算、交配運算、跳脫運算。Acknowledgements
對於這份論文的完成,非常感謝我的指導教授,洪宗貝博士。對
於整個論文的構思、研究、討論及演算法的撰寫,洪老師總是細心給
予指導和正確的建議,並對於我個人沒有考慮的各種方面問題提出質
疑和給予中肯的想法,在之後整個論文的修改上,犧牲了許多洪老師
的私人時間,且教導予我許多撰寫論文的技巧及需注意的事項,在碩
士班兩年的生活,非常感謝洪老師的認真教學,使我成長許多。
同時,我要感謝這兩年來所以指導我的教授們,包括我所參與的
AI Meeting 的林文揚教授以及計算機組的吳志宏教授,除此之外,擔
任口試委員的曾新穆教授及潘正祥教授,均對本篇論文提出相當適當
的建議。還有這兩年來共同在研究室共同努力的學長姐和同學學弟
們,像是詠騏學長在論文閱讀方面的教導、與浚瑋、國能的互相討論
和激勵、俊豪學長的鼓勵,還有其他人的幫忙,如 AI Meeting 的研究
夥伴。研究生的生涯,多了你們增添不少的色彩與回憶。
另外感謝的是家庭的支持,總是鼓勵容易心情低落的我,和一直
支援我繼續升學的決定,總是為我打氣,讓我可以順利完成碩士學
位,謝謝你們。
感謝所有幫助過我的人,致上最深的謝意。
Index
Chapter 1 Introduction ... 5
Chapter 2 Review of Related Works... 8
Chapter 3 The Gene-Set Representation and Operators... 13
3.1 Gene-Set Representation... 13
3.2 The Proposed Gene-Set Mutation Operator... 14
3.3 The Proposed Gene-Set Crossover Operator ... 18
Chapter 4 The Hierarchical Gene-Set Genetic Algorithm... 22
Chapter 5 Theoretical Foundation for the Modified Gene-set Genetic Algorithm ... 25
Chapter 6 A Modified Gene-Set Genetic Algorithm ... 36
Chapter 7 Theoretical Foundation for the Modified Gene-set Genetic Algorithm with Dynamic Mutation Rates ... 40
Chapter 8 A Modified Gene-Set Genetic Algorithm with Dynamic Mutation Rates ... 43
Chapter 9 Experimental Results ... 48
Chapter 1
Introduction
Genetic algorithms (GAs) have become increasingly important to solving difficult
problems because they can provide feasible solutions in a limited amount of time.
They were first proposed by Holland in 1975 [21] and have been successfully applied
to the fields of optimization [7][13][22][38][39][40][41][45][47], machine
learning[6][17][35], neural networks [1][20][30][36][44][46], and fuzzy logic
controllers [28][52], among others [9][19][27][33][34].
When genetic algorithms are used to solve a problem, a representation that
describes the problem states must first be defined. An initial population is then
generated and three genetic operations (crossover, mutation, and reproduction) are
performed to generate the next generation. In the past, a gene was usually regarded as
the basic unit for performing crossover and mutation operations. In this paper, gene
sets, instead of individual genes, are used in the genetic process. The operation length
can thus be logically thought of as the number of gene sets, which is much shorter
representation is a little different from the current one, the original crossover and
mutation operators also need to be modified. A hierarchical gene-set genetic algorithm
is then proposed, which uses adjustable gene-set lengths to find final solutions. At
beginning, a larger gene-set length is set and the genetic process is run for a fixed
number of generations. Then the gene-set length is shortened in half and the genetic
process is run again for a fixed number of generations. The same process is repeated
until the gene-set length is 1. In this way, the proposed genetic algorithm can search
more flexibly in the solution space. Effectively avoiding local optimal trapping has
been an import research topic in GA. The mutation operation is thus critical to the
success of genetic algorithms since it can diversify the search directions and helps GA
escape from local optima. In the other two modified gene-set genetic algorithms,
another escape operation, as well as the mutation operation, is designed for gene-set
genetic algorithms to increase the probability of finding global optima. The property
that a longer gene set causes a larger diversity is first shown. An escape operation
based on the property is designed and a modified gene-set genetic algorithm with the
escape operation is proposed. Using the escape operation, the proposed algorithm can
help the search of global optima if the current search direction is correct and can help
the escape of local optimum if it is not. In the one of modified gene-set genetic
the number of offspring when the gene-set size is large. Different mutation rates are
used for different length of gene sets. Longer gene sets will be allowed larger
mutation rates, thus being able to amend the shortcoming of the generated offspring
due to the longer gene-set size and evolve better solutions. Experiments on three
problems are made to show the effectiveness of the genetic algorithm.
The paper is organized as follows. Genetic algorithms are reviewed in Chapter 2.
The gene-set genetic representation and the gene-set crossover and mutation operators
are introduced in Chapter 3. The hierarchical gene-set genetic algorithm is proposed
in Chapter 4. The property that a longer gene set causes a larger diversity is formally
proven in Chapter 5. A modified gene-set genetic algorithm with the escape operation
is proposed in Chapter 6. The relation of the number of offspring, the mutation rate
and the length of gene sets is shown in Chapter 7. A modified gene-set genetic
algorithm with dynamic mutation rates is proposed in Chapter 8. Experimental results
for showing the performance of the proposed algorithm are described in Chapter 9.
Chapter 2
Review of Related Works
When genetic algorithms are used to solve a problem, a representation that
describes the problem’s states must first be defined. Several chromosome
representations have been proposed and commonly used, such as binary strings,
real-value vectors, permutations, finite–state representation, and parse-tree
representation. Binary strings [4][5][16][19][23] are the standard and the most
commonly used representation of solutions for genetic algorithms. They use only the
two symbols 0 and 1 to represent a chromosome. Real-valued vectors [16] are another
popular representation used in GA. Each position in a chromosome is a real value.
Real-value vectors are especially useful for solving real-value optimization problems.
Permutations are a popular representation for some combinatorial optimization
problems [15][28][39][42][45][50]. They encode the set of objects into numbers and
then arrange them into a chromosome. As to the finite-state representation [2][14][22],
it first constructs a state transition table according to the given problems, and then
evolve according to the transitive table. This method is used for the environment in
the relation. In addition, the parse-tree representation [3][8][26][29][34][40] is often
used for evolving executable structures, such as a program. Each chromosome is
represented by a parse tree.
After a representation has been chosen for a problem, the genetic process begins.
An initial population is generated and three genetic operations (crossover, mutation,
and reproduction) are performed to generate the next generation. The simple genetic
algorithm uses a single crossover operator and a single mutation operator throughout
the entire genetic process. The same procedure is then repeated until the termination
criterion is satisfied. The simple genetic algorithm is described as follows.
The Simple Genetic Algorithm:
Step 1: Define a suitable representation for the problem to be solved.
Step 2: Create an initial population of N individuals for evolution.
Step 3: Define a suitable fitness function for the individuals.
Step 4: Perform genetic operations (crossover and mutation) to generate possible
offspring.
Step 5: Evaluate the fitness value of each individual.
Step 7: If the termination criterion is not satisfied, go to Step 4; otherwise, stop the
algorithm.
The simple genetic algorithm uses several parameters such as population size,
crossover probability pcrossover and mutation probability pmutation. The crossover
operator usually exchanges some bits between two individuals with probability
pcrossover. Valuable information of the parents can thus be shared among the offspring. The mutation operator is applied to each single individual with a small probability
pmutation. The mutation operator preserves a reasonable level of population diversity and prevents premature convergence to local optima. The values of pcrossover and
pmutation are known to critically affect the behavior and performance of the genetic algorithm[47].
Some researches about dynamical adjustment of crossover and mutation rates
were thus proposed. Srinivas and Patnaik proposed the adaptive genetic algorithm
(AGA), in which pcrossover and pmutation are varied according to the fitness values of the
solutions [47]. Fogarty used a varying mutation rate and demonstrated that a mutation
rate decreasing exponentially over generations has superior performance [13]. Davis
defined five fixed operators to replace the original crossover and mutation operators
probabilities. Hong et al. proposed a dynamic genetic algorithm (DGA), which
simultaneously used more than one crossover and mutation operators to generate the
next generation. The crossover and mutation ratios changed along with the evaluation
results of the respective offspring in the next generation [21]. This paper tries to improve the performance of the genetic algorithms from a different aspect. Gene sets,
instead of individual genes, are used and dynamically adjusted in the genetic process
to speed up convergence.
Some common crossover operators are multiple-point crossover[46], uniform
crossover[48], one-point crossover, substring crossover, etc. They are briefly
described as follows.
1. Multi-point crossover method: This method defines a mask to determine
which bits should be exchanged between two individuals. For the positions which are
1 on the mask, the parents exchange the corresponding bits.
2. Uniform crossover method: This method defines a mask to determine which
bits should be exchanged between two individuals. Bit values of 1 and 0, however,
alternative with each other on the mask.
3. One-point crossover method: There is only one bit with value 1 on the mask.
That is, the operator randomly selects a single bit within two parents to perform
4. Substring crossover method: This method changes arbitrary substrings
between two individuals. Lengths and positions of these substrings are chosen at
random, but are the same for both individuals.
Some common mutation operators are swapping mutation, inversion mutation,
and bit-change mutation, among others. They are briefly described as follows :
1. One-point mutation method: This method changes one bit of a chromosome.
2. Swapping mutation method: This method exchanges arbitrary two bits in a
chromosome.
3. Inversion mutation method: This method changes the order of the bits in an
arbitrary interval of a chromosome.
4. Bit-change mutation method: This method changes the bit value 0 to 1 and 1
Chapter 3
The Gene-Set Representation and Operators
In this paper, the gene-set genetic algorithm is proposed to speed up the
convergence of the genetic process. The representation is first described below.
3.1 Gene-Set Representation
A gene set is a set of genes, which combines two or more neighboring genes
together into a unit and operates like a composite gene. Figure 1 shows the gene-set
representation. n genes a gene-set gn-m+1gn-m+2…gn … … … gm+1gm+2…g2m g1g2…gm
Figure 1: gene-set representation
In Figure 1, there are totally n genes and each gene set consists of m genes. m
and n are usually 2’s exponential expressions. Let n = 2a and m = 2b. The number of
example in which 8 gene sets are formed from a chromosome with 32 genes.
When gene sets, instead of individual genes, are used in the genetic process, the
operation length can be logically thought of as the number of gene sets, which is
much shorter than the total gene length. The convergence speed is thus faster than that
in the original GA, and the execution time needed can be reduced. The experimental
results described in Section 5 will show the effect. Note that the gene-set
representation can also be easily extended to other gene types in addition to binary
bits.
3.2 The Proposed Gene-Set Mutation Operator
Since the gene-set representation is a little different from current one, the genetic
operations must also be modified or re-designed. The gene-set mutation operation is
first proposed here. As mentioned above, a gene set is a unit which is operated as a
whole. Assume each gene set in a chromosome is chosen for mutation with the
probability of pmu. If a gene set has been chosen for mutation, the proposed mutation 0001 1101 1111 1100 1011 1011 1100 1000
probability except itself. For binary-bit genes, the probability can be easily derived as
1/(2m-1), where m is the length of a gene set. Figure 3 shows an example of the
proposed mutation operation for a 4-bits gene set. Assume the gene set “0000” has
been chosen for mutation. It may be converted into one of the other 15 different gene
Note that the conventional single-point mutation operator sets a mutation rate for
each gene. That is, each gene may be mutated from 0 to 1 or vice verse according to
the given mutation rate. In this way, the probability for several neighboring genes to
simultaneously mutate will be much smaller than that for a single gene. For example,
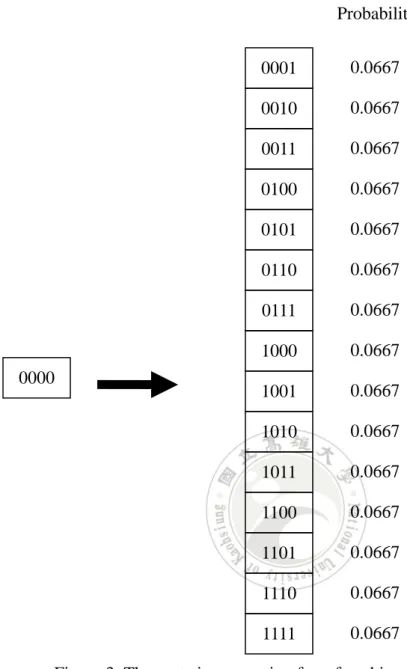
assume the four genes “0000” are to be mutated and the mutation rate is 0.04. The 0000 0001 0010 0011 0100 0101 0110 0111 1000 1001 1010 1011 1100 1101 1110 1111 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 0.0667 Probability
fifteen possible mutation results with their probability by the conventional
single-point mutation operator are shown in Figure 4.
Note that the proposed gene-set mutation operator will assign the same 0000 0001 0010 0011 0100 0101 0110 0111 1000 1001 1010 1011 1100 1101 1110 1111 0.2349 0.2349 0.0098 0.2349 0.0098 0.0098 0.0004 0.2349 0.0098 0.0098 0.0004 0.0098 0.0004 0.0004 0.00002 Possibility
Figure 4: The probability distribution from the conventional single-point mutation operator
probability to all the possible offspring. It will thus have a larger diversity than the
conventional one.
3.3
The Proposed Gene-Set Crossover Operator
In this paper, a gene-set substring crossover operator is proposed for generating
possible offspring. The conventional substring crossover operator exchanges arbitrary
substrings between two individuals. In the proposed gene-set substring crossover
operator, the crossover points at two parents must be the same and are randomly
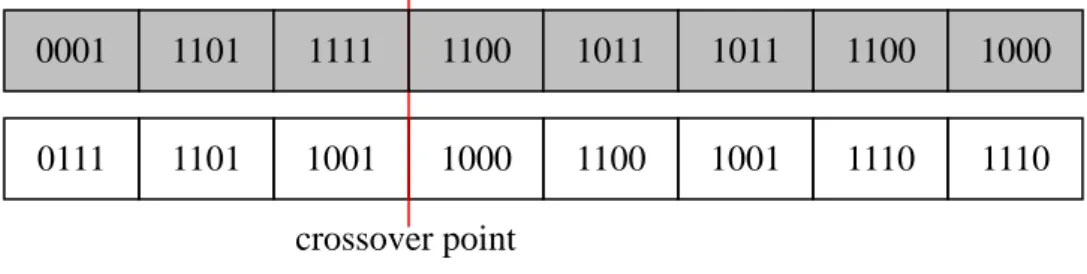
chosen at the boundary of gene sets. An example for the proposed gene-set crossover
operator is shown in Figure 6, where the crossover point is chosen at the end of the
third gene set. The two parent chromosomes in Figure 6(a) will crossover to produce
Note that the average length of the shorter substring in a parent chromosome by
the gene-set substring crossover operator will be a little longer than that of the
conventional one. For example in a 32-bit chromosome, the gene-set substring
crossover will set the crossover point at the boundary. There are thus seven possible
crossover points. The average length of the shorter substring for the proposed
crossover operation is calculated as (2×(4+8+12)+16)/7, which is 9.1428. The average
length of the shorter substring by the original substring crossover operation is (2×
(1+2+3+…+15)+16)/31, which is 8.2580. It is easily observed that the average length
of the shorter substring by the proposed crossover operator is longer than that by the
conventional one. The proposed crossover operator will thus in average cause wider 0001 1101 1111 1100 1011 1011 1100 1000 0111 1101 1001 1000 1100 1001 1110 1110 1100 1011 1011 1100 1000 0111 1101 1001 1000 1100 1001 1110 1110 0001 1101 1111
Figure 6: A crossover example for chromosomes with 32 bits and 4-bit genes crossover point
(a) The parent chromosomes before crossover
discrepancy between offspring and parents.
Formally, let the length of each chromosome is n and the length of each gene set
is m. When both n and m are two’s exponential expressions, the number of gene sets
will be even. In this case, the average length (AL1)of the shorter substring by the
proposed crossover operator is calculated as follows:
AL1 = [(m+2m+…+((n/2)-m)+n/2+((n/2)-m)+((n/2)-2m)+…+m)]/(n/m-1) = [2×((m+n/2-m)×((n/2)/m-1)/2)+n/2]/(n/m-1) = [(m+n/2-m)×((n/2)/m-1)+n/2]/(n/m-1) = [(n/2×((n/2)/m-1)+n/2)]/ (n/m-1) = ((n/2)2/m)/((n/m)-1) =(n/2)2/(n-m).
Similarly, the average length (AL2) by the original crossover operator is
calculated as follows: AL2 = [1+2+3+…+(n/2-1)+n/2+(n/2-1)+(n/2-2)+…+1]/(n-1) = [2×((1+n/2-1)×(n/2-1)/2)+n/2]/(n-1) = [(1+n/2-1)×(n/2-1)+n/2]/(n-1) = [n/2×(n/2-1)+n/2]/(n-1) = (n/2)2/(n-1).
Theorem: Let n be the length of each chromosome, m be the length of each gene
set. Assume n and m are two’s exponential expressions, m<n and gcd(m,n)=m. If m>1,
the average length of the shorter substring by the proposed gene-set substring
crossover operator is longer than that by the original substring crossover operator.
Proof: According to the above derivation, the average length of the shorter
substring by the proposed crossover operator is (n/2)2/(n-m), and is (n/2)2/(n-1) by the
Chapter 4
The Hierarchical Gene-Set Genetic Algorithm
A genetic process is rough when the gene-set length is large as the entire large
gene set is processed as a whole. It is thus suitable to the beginning of a genetic
process. On the contrary, the genetic process can be refined if the length is set at a
small value. A hierarchical gene-set genetic algorithm is thus proposed here to
effectively and efficiently find nearly optimal solutions to a problem. At beginning, a
larger gene-set length is set, and the genetic process is run for a fixed number of
generations (or until convergence). Then the gene-set length is shortened in half and
the genetic process is run again for a fixed number of generations (or until
convergence). The same process is repeated until the gene-set length is 1. In this way,
the proposed genetic algorithm can search more flexibly in the encoding space. A
simple example is shown in Figure 7, where the gene-set length is initially set at 4.
The genetic process for each gene-set length is run for n generations. After n
1 ~ n Generations n+1 ~ 2n Generations 2n+1 ~ 3n Generations
1011
1111
1000
1010
11 10 11 11 00 10 10 10 1 1 0 1 1 1 1 1 0 0 0 1 0 1 0 1The proposed hierarchical gene-set algorithm is described below.
The hierarchical gene-set genetic algorithm:
INPUT: A population size P, a chromosome size w, an initial gene-set length l, a
mutation rate rm,, a crossover rate rc, and a number N of generations in each
phase.
OUTPUT: A nearly optimal solution.
Step 1: Define a suitable chromosome representation with size w for the problem to be
solved.
Step 2: Define a suitable fitness function for evaluating the individuals.
Step 3: Randomly generate a population of P individuals.
Step 4: Set k = l, where k is used to keep the gene-set length currently being Figure 7: An example for gradually adjusting gene-set length
Step 5: Set n = 1, where n is used to count the number of generations.
Step 6: Execute gene-set crossover and mutation operations on the population.
Step 7: Evaluate the fitness value of each individual.
Step 8: Select the superior P individuals according to their fitness values.
Step 9: Set n = n+1.
Step 10: If n < N, repeat Steps 6 to 10; otherwise, do the next step.
Step 11: set k = k/2.
Step 12: If k < 1, stop the algorithm and output the best chromosome in the current
generation as the result; otherwise, go to Step 5.
After the process of the hierarchical gene-set genetic algorithm, a nearly optimal
Chapter 5
Theoretical Foundation for the Modified
Gene-set Genetic Algorithm
In this section, the property that a longer gene set causes a larger diversity is
formally shown. Both the crossover and the mutation operations in a long gene set
will behave better than in a short one. The derivation for the crossover operation is
first given below.
Lemma 1: Let the length of each chromosome be n and the length of each gene
set be m. When both n and m are two’s exponential expressions, the number of gene
sets will be even if n>m, and will be 1 if n=m.
Proof: Let n=2a and m=2b, a≧b. The number of gene sets is thus 2a-b, which is
even when a>b, and is 1 when a=b. ■
Note that in implementation, m is set smaller than n; otherwise, the gene-set
operation will generate a new totally random chromosome.
Lemma 2: Let the length of each chromosome be n and the length of each gene
set be m, n>m. The average length (AL) of the shorter substring by the gene-set
substring crossover operator is (n/2)2/(n-m).
Proof: By Lemma 1, when n>m, the number of gene sets is even. Thus:
AL = [(m+2m+…+((n/2)-m)+n/2+((n/2)-m)+((n/2)-2m)+…+m)]/(n/m-1) = [2×((m+n/2-m)×((n/2)/m-1)/2)+n/2]/(n/m-1) = [(m+n/2-m)×((n/2)/m-1)+n/2]/(n/m-1) = [(n/2×((n/2)/m-1)+n/2)]/ (n/m-1) = ((n/2)2/m)/((n/m)-1) = (n/2)2/(n-m). ■
Theorem 1: Let n be the length of each chromosome, and m1 and m2 be two
gene-set sizes. Assume n, m1 and m2 are all two’s exponential expressions. If m1> m2,
the average length of the shorter substring by the gene-set substring crossover
operator for m1 is longer than that for m2.
Proof: According to Lemma 2, the average length of the shorter substring by the
m2. If m1> m2, then n-m1<n-m2. Thus, (n/2)2/(n-m1) >(n/2)2/(n-m2). ■
According to Theorem 1, long gene-set sizes will get larger average substring
change by crossover operations than short ones, thus causing larger diversity. The
derivation for the mutation operation is given below.
Lemma 3: Let the length of each gene set be m, which is a two’s exponential
expression. When a gene set has been mutated, the number of all possible mutation
results is 2m-1.
Proof: When a set has been mutated, the number of combinations for the gene set to change j bits isCmj , j≧1. The number of combinations is thus:
1 2 0 0 1 − = − =
∑
∑
= = m m m j m j m j m j C C C . ■Lemma 4: Let the length of each gene set be m, which is a two’s exponential
expression. When a gene set has been mutated, the average number of changed bits in
the gene set is
2 1 2 2 m m m ⋅ − .
Proof: When a gene set G has been mutated, the number of combinations for the gene set to change j bits is Cmj , m≧j≧1. Its probability Pj is thus
(
2 −1)
=∑
= m m j m m k m j C Cand k=0 is excluded. Since the length m≧1, the average number of changed bits
(ACB) for G is derived as follows:
( )
(
(
)
)
(
(
)
)
(
(
)
(
)
(
)
)
(
)
(
)
(
)
⋅ = = − = = − = = ⋅ − = ⋅ ⋅ − = ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ ⋅ − = ⎟ ⎟ ⎟ ⎠ ⎞ ⎜ ⎜ ⎜ ⎝ ⎛ ⎟ ⎠ ⎞ ⎜ ⎝ ⎛ ⋅ + ⋅ − = ⎟ ⎟ ⎟ ⎠ ⎞ ⎜ ⎜ ⎜ ⎝ ⎛ ⋅ − = − ⋅ − + − ⋅ = − = − =∑
∑
∑
∑
∑
∑
2 1 2 2 2 2 1 2 1 2 1 2 1 2 2 1 2 1 1 2 1 1 2 1 2 1 2 / 1 2 0 2 0 2 0 2 0 0 1 m m C m C m C m C m C j m C j C j C j G ACB m m m m m j m j m m j m j m m j m m j m j m m j m m j m m m j m j m m j m j m m j ■Lemma 5: Let two neighboring gene sets G1 and G2 exist, each with length of m,
which is two’s exponential expression. When the mutation rate for a gene set is Pm
and when at least one of the two gene sets has mutated, the conditional probability P1
for only G1 (or G2) to mutate is (1-Pm)/(2-Pm). The conditional probability P2 for both
G1 and G2 to simultaneously mutate is Pm/(2-Pm).
Proof: When the mutation rate for a gene set is Pm, the probability for only G1 to
mutate is Pm×(1- Pm), for only G2 is Pm×(1- Pm), and for both G1 and G2 is Pm×Pm.
When at least one of the two neighboring gene sets has mutated, the conditional
probability for only G1 (or only G2) to mutate is thus:
(
)
(
m)
m(
m(
(
m)
)
)
Pm P P P P P P P P P P P − − = + − ⋅ − = + − − 2 1 1 2 1 1 2 1 2 .The conditional probability for both G1 and G2 to mutate is thus:
(
)
(
(
)
)
m m m m m m m m m m m P P P P P P P P P P P − = + − ⋅ ⋅ = + − 2 1 2 1 2 2 2 . ■Lemma 6: Let two neighboring gene sets G1 and G2 exist, each with length of m,
which is two’s exponential expression. When the mutation rate for a gene set is Pm
and when at least one of the two gene sets has mutated, the average number of
changed bits in the two neighboring gene sets is
(
P)
m m m m m m ⋅ − ⋅ ⋅ − + 1 2 2 2 2 1 2 2 2 .Proof: When the mutation rate for a gene set is Pm and when at least one of the
two gene sets has mutated, the conditional probability for G1 (or G2) is P1, and for
both G1 and G2 is P2 according to Lemma 5. The average number of changed bits in
the two gene sets is derived as follows:
(
)
( )
( )
(
( )
( )
)
( )
( ) (
)
(
)
(
) (
)
(
)
(
−)
⋅ ⋅ − ⋅ ⋅ + = ⋅ − ⋅ + ⋅ − = ⋅ − ⋅ + ⋅ − ⋅ − ⋅ − = ⋅ − ⋅ − ⋅ − ⋅ − = ⋅ − ⋅ − + − = ⋅ − ⋅ + = ⋅ − ⋅ + = ⋅ + ⋅ = + ⋅ + ⋅ + ⋅ = + m P m P m P m P m P P P m P P m P P G ACB P G ACB P G ACB G ACB P G ACB P G ACB P G G ACB m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m m 1 2 2 2 2 1 2 1 2 2 2 1 2 2 1 1 2 2 2 1 2 1 2 1 2 2 2 1 1 2 2 2 1 2 1 2 2 2 1 1 2 2 2 1 1 2 2 2 1 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 1 2 1 1 2 1 1 2 1 2 2 1 1 1 2 1■ Lemma 7: If
(
)
m m m P 2 2 1 2 ⋅ − +﹤1 for any positive integer m ,then Pm < m
m 2 1 2 − . Proof:
(
)
m m m P 2 2 1 2 ⋅ − + <1, Æ(
)
m m m P 2 2 1 2 + < − ⋅ , Æ2mPm <2m −1, Æ m m m P 2 1 2 − < . ■Theorem 2: Let a gene set G of length 2m consist of two sub gene sets G1 and G2,
each with a length of m, which is a two’s exponential expression.When the mutation
rate Pm for a gene set is less than m m
m ⋅ −1 2 2 2 2
and when the genes of length 2m has
been mutated, the average number of changed bits for 2m as a gene set unit is larger
than that for m as a gene set unit.
Proof: According to Lemma 4, the average number of changed bits for mutating
G is m m m ⋅ −1 2 2 2 2
. According to Lemma 6, the average number of changed bits for
mutating G1 and G2 is
(
(
)
)
m P P m m m m m m ⋅ − ⋅ ⋅ − + ⋅ 1 2 2 2 2 1 2 2 2. When Pm is less than m
m 2 1 2 − ,
(
)
(
)
m m m m P P 2 2 1 2 ⋅ − + ⋅﹤ 1 by Lemma 7. Thus, when Pm is less than m
m 2 1 2 − ,
(
)
(
P)
m P m m m m m m ⋅ − ⋅ ⋅ − + ⋅ 1 2 2 2 2 1 2 2 2 < m m m ⋅ −1 2 2 2 2. The average number of changed bits for 2m as
Corollary: Let m be a two’s exponential expression, when the mutation rate Pm
for a gene set is less than 0.5 and when the genes of length 2m has been mutated, the
average number of changed bits for a gene set unit of 2m is larger than that for a gene
set unit of m, m≧1.
Proof: According to Theorem 2, when Pm is less than m
m
2 1 2 −
and when the
genes of length 2m has been mutated, the average number of changed bits for a gene
set unit of 2m is larger than that for a gene set unit of m. Since the function m
m
2 1 2 −
is
strictly increasing and m≧1, so the minimum value of the function appears when
m=1. Thus, m m 2 1 2 − ≦ 0.5 2 1 2 1 1 = −
. Therefore, when Pm is less than 0.5, the average
number of changed bits for a gene set unit of 2m is larger than that for a gene set unit
of m, m≧1. ■ Lemma 8: When Pm1> Pm2,
(
)
(
)
(
(
)
n)
m m n m m P P P P 2 2 1 1 1 1 1 1− − > − − for anyn≧2.Proof: We use the mathematical induction to prove it. When n=2, the following
formulas hold:
(
)
(
)
(
) (
2)
2 2 2 1 1 2 2 1 2 1 2 1 1 1 2 1 1 2 2 2 2 1 1 m m m m m m m m m m m m m P P P P P P P P P P P P P − ⋅ − − = − = − − , and(
)
(
)
(
) (
2)
2 2 2 1 1 2 2 1 2 1 2 2 2 2 2 2 2 2 2 2 2 1 1 m m m m m m m m m m m m m P P P P P P P P P P P P P − ⋅ − − = − = − − .When Pm1> Pm2, Pm1Pm22 =Pm1Pm2Pm2 <Pm1Pm1Pm2 =Pm21Pm2. Thus,
(
) (
) (
) (
2)
2 2 2 1 1 2 2 1 2 1 2 2 2 2 1 1 2 2 1 2 1 2 2 2 2 2 2 m m m m m m m m m m m m m m m m P P P P P P P P P P P P P P P P − ⋅ − − > − ⋅ − − .Then we assume the conclusion holds for n = k and prove that it will also hold
for n = k+1. Thus when n = k, the following formula holds:
(
)
(
)
(
(
)
k)
m m k m m P P P P 2 2 1 1 1 1 1 1− − > − − . It implies Pm1−Pm1⋅(
1−Pm2)
k >Pm2 −Pm2⋅(
1−Pm1)
k.The following formula can be derived for n = k+1:
(
)
(
)
(
(
(
)
(
)
(
(
)
)
)
)
(
(
)
(
)
(
) (
(
)
)
1)
2 1 1 2 2 1 1 1 2 1 1 1 2 1 1 1 1 1 1 1 1 1 . 1 1 1 1 1 1 1 1 1 + + + + + + − − ⋅ − − − − − = − − ⋅ − − − − = − − k m k m m k m m m k m k m k m m k m m P P P P P P P P P P P P(
)
(
)
(
)
(
)
(
(
)
1)
2 1 1 2 2 1 2 1 1 1 1 1 1 1 1 + + ⋅ − − − − − + − − = k m k m k m m m k m m m P P P P P P P P . Similarly,(
)
(
)
(
(
(
)
)
)
(
(
(
)
1)
)
2 1 1 1 2 1 1 2 2 1 2 2 1 1 1 1 1 1 1 1 + − − + ⋅ − − + − + − − = − − k m k m k m m m k m m m k m m P P P P P P P P P P .When Pm1> Pm2, the following formulas can hold fork >1:
(
)
(
)
k m m m k m m m P P P P P P 1− 1 1− 2 > 2 − 2 1− 1 , and(
)
(
)
k m m m k m m m P P P P P P 1 2 1− 2 > 1 2 1− 1 . Thus,(
)
(
)
(
)
(
)
(
(
)
)
(
(
(
)
)
)
(
(
(
)
1)
)
2 1 1 1 2 1 1 2 2 1 2 1 1 2 2 1 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 + + + + − − ⋅ − − − + − − > − − ⋅ − − − + − − k m k m k m m m k m m m k m k m k m m m k m m m P P P P P P P P P P P P P P P P . It implies(
)
(
)
(
(
)
1)
2 1 1 1 1 1 1− − + > − − k+ m k m P P P P .Thus according to the mathematical induction, when Pm1>Pm2, the following
inequality holds any n≧2.
(
)
(
)
(
(
)
n)
m m n m m P P P P 2 2 1 1 1 1 1 1− − > − − . ■Lemma 9: Let a gene set G consist of n sub gene sets, n≧2. When the gene set
is mutated by the gene-set mutation operator according to the sub-gene-set size, the
average number of mutated sub gene sets with mutation rate Pm1 is larger than that
with Pm2 if Pm1> Pm2.
Proof: When a gene set G is mutated with the mutation rate Pm1 by the gene-set
mutation operator, the conditional possibility for a sub gene set to mutate is Pm1/(1-(1-
Pm1)n). Its average number of mutated sub gene sets is
(
)
(
n)
m m P P n 1 1 1 1− − × . Similarly,the average number of mutated sub gene sets with the mutation rate Pm2 is
(
)
(
n)
m m P P n 2 2 1 1− − × . When Pm1>Pm2,(
)
(
)
(
(
)
n)
m m n m m P P P P 2 2 1 1 1 1 1 1− − > − − for any n≧2according to Lemma 8. Thus, when the gene set is mutated by the gene-set mutation
operator according to the sub-gene-set size, the average number of mutated sub gene
sets with mutation rate Pm1 is larger than that with Pm2 if Pm1> Pm2. ■
Theorem 3: Let a gene set G of length m consist of n sub gene sets, each with
a gene set to mutate is less than 0.5 and when the gene set G has been mutated, the
average number of changed bits for mutating G is larger than that for mutating sub
gene sets.
Proof: When the mutation rate Pm is set at 0.5, the probability of mutating a
certain group of k sub gene sets is 0.5x(1-0.5)n-x , which is equal to 0.5n. The
probability of mutating any arbitrary group of sub gene sets is thus the same for Pm =
0.5. When the gene set G has been mutated, the number of combinations for j sub
gene sets to be mutated is Cmj , j≧1. Its probability Pj is ( (2 1))
1 − =
∑
= n n j n k n k n j C C C ,where k starts from 1 since the gene set has been mutated and k=0 has been excluded.
When n>1, the average number of mutated sub gene sets is derived as follows:
(
)
(
)
(
(
)
(
)
(
)
)
(
)
(
)
(
)
⋅ = = − = = − = ⋅ − = ⋅ ⋅ − = ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ ⋅ − = ⎟ ⎟ ⎟ ⎠ ⎞ ⎜ ⎜ ⎜ ⎝ ⎛ ⎟ ⎠ ⎞ ⎜ ⎝ ⎛ ⋅ + ⋅ − = ⎟ ⎟ ⎟ ⎠ ⎞ ⎜ ⎜ ⎜ ⎝ ⎛ ⋅ − = − ⋅ − + − ⋅ = −∑
∑
∑
∑
∑
2 1 2 2 2 2 1 2 1 2 1 2 1 2 2 1 2 1 1 2 1 1 2 1 2 1 2 0 2 0 2 0 2 0 1 n n C n C n C n C n C j n C j C j n n n n n j n j n n j n j n n j n n j n j n n j n n j n n n j n j n n jAccording to Lemma 4, when a sub gene set has been mutated, the average
number of changed bits in the sub gene set is
2 1 2 2 z z z ⋅
− . Thus, the average number of changed bits in the mutated n sub gene sets is )
2 1 2 2 ( ) 2 1 2 2 ( n z z z n n ⋅ − ⋅ ⋅ − , which is 1 2 2 1 2 2 4 ⋅ − ⋅ z − z n n m . Since both 1 2 2 − n n and 1 2 2 − z z
are strictly decreasing for positive
the mutated n sub gene sets is thus less than , 1 2 2 1 2 2 4 1 1 2 2 − ⋅ − ⋅ m which is ) 3 2 ( 1 2 3 4 4 m m⋅ ⋅ =
. Besides, the average number of changed bits for mutating G is
2 1 2 2 m m m ⋅
− according to Lemma 4. Since 2 1 2 2 3 2m m m m ⋅ −
≤ for m≧2, the average
number of changed bits for mutating G is larger than that for mutating n sub gene sets
when the mutation rate is set at 0.5.
According to Lemma 9, if the mutation rate is less than 0.5, the average number
of mutated sub gene sets is less than
⋅ ⋅ − 21 2 2 n n n
. It implies that the average number of
changed bits is less than )
2 1 2 2 ( ) 2 1 2 2 ( n z z z n n ⋅ − ⋅ ⋅ − , which is equal to 1 2 2 1 2 2 4 ⋅ − ⋅ z − z n n m
. Thus, when the mutation rate Pm for a gene set is less than 0.5 and
when the gene set G has been mutated, the average number of changed bits for
mutating G is larger than that for mutating n sub gene sets. ■
From the above theorems, it can be seen that when a mutation or crossover
operator has been performed, using a longer gene set can cause more bits to be
changed than using a shorter one. The former can thus generate more different
Chapter 6
A Modified Gene-Set Genetic Algorithm
When a population converges, the solutions may be local optimal or global
optimal. Although the mutation operation can be used to help the genetic process
escape from local optimums, the escape speed is slow and depends on the mutation
rate. As mentioned above, the gene sets with a larger length will have larger variation.
This characteristic will be used here to speed up escape from local optima.
The escape operator based on this characteristic can easily be designed as fallows.
Let the current gene-set length is m. If the genetic process for the length has
converged, but has not gotten to the assigned number of generations, the gene-set
length will be changed to a value bigger than m. The above escape operator, when
used together with the other genetic operators, will thus cause a large variation to help
a population escape from local optima. The escape operator is performed when the
best solutions in the current population converge. The gene-set crossover and
mutation processes are then run for the enlarged gene-set length for only one
generation. After that, it is run for the original length until the best solutions converge.
predefined number of generations is achieved. The next genetic phase for a half
gene-set length then begins. Before the proposed algorithm is described in detail, the
concept of (δ, k)-converging is defined below.
Definition: (δ, k)-converging - Let δ>0 and best(r) be the best solution in the
population of the r-th generation. The r-th generation is called (δ, k)-converging if and
only if |best(r)– best(r–z+1)|≦δ for every z≦k.
When a generation is (δ, k)-converging, the best solution in that generation has
not been improved in recent k generations. Two possible situations may exist. One is
the global optimal solution has been found, and the other is the population has lost its
variety and has been trapped into local optima. The above escape operation will be
integrated in the proposed algorithm to avoid the occurrence of the latter situation.
The modified gene-set genetic algorithm with escape operation is described below.
The modified gene-set genetic algorithm with escape operation:
INPUT: A population size P, a chromosome size n, an initial gene-set length l1, a
mutation rate rm,, a crossover rate rc, an enlarged gene-set length l2 for escape
operation, a number N of generations in each phase, and two parameters δ
Step 1: Define a suitable chromosome representation with size n for the problem to be
solved.
Step 2: Define a suitable fitness function for evaluating the individuals.
Step 3: Randomly generate a population of P individuals.
Step 4: Set l = l1, where l is used to keep the gene-set length currently being
processed.
Step 5: Set g = 1, where g is used to count the number of generations.
Step 6: Execute the gene-set crossover and mutation operations on the population.
Step 7: Evaluate the fitness value of each individual.
Step 8: Select the superior P individuals according to their fitness values.
Step 9: Set g = g+1.
Step 10: If g < N and the current generation is not (δ, k)-converging, go to Step 6.
Step 11: If g < N and the current generation is (δ, k)-converging, do the escape
operation as follows:
Substep 11-1: Set temp = l, where temp is used to keep the current gene-set
length.
Substep 11-2: Set the gene-set length l = l2.
Substep 11-3: Execute the gene-set crossover and mutation operations for
Substep 11-4: Set g = g+1.
Substep 11-5: Set l = temp.
Substep 11-6: Go to Step 6.
Step 12: If g = N, set l = l/2.
Step 13: If l < 1, stop the algorithm and output the best chromosome in the current
generation as the result; otherwise, go to Step 5 for the next genetic phase.
After the process of the modified gene-set genetic algorithm, a nearly optimal
solution to the given problem can be found. Using the escape operation, the proposed
algorithm can help the escape of local optima through the change of gene-set sizes to
a large value. Even when the population has (δ, k)-converged, the proposed algorithm
can also help the search of global optima since the gene-set operations with the
original gene-set length will still be done as long as the predefined number of
termination generations is not achieved. The modified gene-set genetic algorithm can
Chapter 7
Theoretical Foundation for the Modified
Gene-set Genetic Algorithm with Dynamic
Mutation Rates
In this section, the property that using longer gene sets will cause the population
to produce less offspring is formally shown. The derivation is given below.
Lemma 1: Let a chromosome S of length m consist of m/n gene sets of length n,
m/n is an integer. When the mutation rate is Pm, the probability of a chromosome which is mutated is 1-(1- Pm)m/n.
Proof: When the mutation rate is Pm, the probability for a gene set not to mutate
is (1- Pm). Thus the probability for all the gene sets in the chromosome S not to mutate
is (1- Pm)m/n. The probability for the chromosome S to mutate is 1-(1-Pm)m/n. ■
Lemma 2: Let a population P consist of x chromosomes of length m, each of
is Pm, the expected value of the number of offspring produced from P by mutation is x
×(1-(1- Pm)m/n).
Proof: According to Lemma 1, the probability of a chromosome which is
mutated is 1-(1- Pm)m/n. Thus, the expected value of the number of offspring produced
from P by the mutation operator is x×(1-(1- Pm)m/n). ■
Theorem 1: Let there be two populations P1 and P2, each of which consists of x
chromosomes of length m. Assume each chromosome in P1 consists of m/n1 gene sets
of length n1, and each chromosome in P2 consists of m/n2 gene sets of length n2, where
m/n1 and m/n2 are integers. When the mutation rate is Pm and n1 > n2, P1 will in
average produce less offspring than P2 by the gene-set mutation operator.
Proof: According to Lemma 2, the expected value of the number of offspring
from P1 by the gene-set mutation operator is x×(1-(1- Pm)m/n1), and from P2 is x×(1-(1-
Pm)m/n2). When n1 > n2, it can be derived that m/n1 < m/n2 and x×(1-(1-Pm)m/n1) < x×
(1-(1-Pm)m/n2). P1 will thus in average produce less offspring than P2 by the gene-set
mutation operator. ■
From the above theorem, it can be concluded that using longer gene sets will
be allowed larger mutation rates, thus being able to mend the shortcoming of the
Chapter 8
A Modified Gene-Set Genetic Algorithm with
Dynamic Mutation Rates
When a population converges, the solutions may be local optimal or global
optimal. Although the mutation operation can be used to help the genetic process
escape from local optimums, the escape speed is slow and depends on the mutation
rate. As mentioned above, the gene sets with a larger length will have larger variation.
An escape operator based on this characteristic can easily be designed as fallows. Let
the current gene-set length is m. If the genetic process for the length has converged,
but has not gotten to the assigned number of generations, the gene-set length will be
changed to a value bigger than m. The above escape operator, when used together
with the other genetic operators, will thus cause a large variation to help a population
escape from local optima. The escape operator is performed when the best solutions in
the current population converge. The gene-set crossover and mutation processes are
then run for the enlarged gene-set length for only one generation. After that, it is run
enlarged again. The above process is repeated until the predefined number of
generations is achieved. The next genetic phase for a half gene-set length then begins.
In addition, from the above theorem, using longer gene sets will cause a
population to produce less offspring. Longer gene sets will thus be allowed larger
mutation rates, thus being able to amend the shortcoming of the generated offspring
due to the longer gene-set size and evolve better solutions. There are thus two
principles for adjusting the mutation rates to be followed. The first one is that the
mutation rate at a longer gene set is larger than that at a smaller one; the second one is
that all the mutation rates can not be set too large to guarantee the performance
becomes poorer than that by the simple GA. In this paper, the proposed algorithm will
simultaneously consider the escape operation and the dynamic mutation rate to
effectively help the escape of local optimums.
The modified gene-set genetic algorithm with the escape operation and dynamic
mutation rates is described below.
The modified gene-set genetic algorithm with dynamic mutation rates:
INPUT: A population size P, a chromosome size n, an initial gene-set length l1, an
generations in each phase, and two parameters δ and k.
OUTPUT: A nearly optimal solution.
Step 1: Define a suitable chromosome representation with size n for the problem to be
solved.
Step 2: Define a suitable fitness function for evaluating the individuals.
Step 3: Randomly generate a population of P individuals.
Step 4: Set l = l1, where l is used to keep the gene-set length currently being
processed.
Step 5: Set g = 1, where g is used to count the number of generations.
Step 6: Execute the gene-set crossover and mutation operations on the population
using the crossover rate rc and the mutation rate rmt for the gene-set length of
t.
Step 7: Evaluate the fitness value of each individual.
Step 8: Execute the selection mechanism to generate the next population.
Step 9: Set g = g+1.
Step 10: If g < N and the current generation is not (δ, k)-converging, go to Step 6.
Step 11: If g < N and the current generation is (δ, k)-converging, do the escape
Substep 11-1: Set temp = l, where temp is used to keep the current gene-set
length.
Substep 11-2: Set the gene-set length l = l2.
Substep 11-3: Execute the gene-set crossover and mutation operations on the
population using the crossover rate rc and the mutation rate
rml for the gene-set length l. Substep 11-4: Set g = g+1.
Substep 11-5: Set l = temp.
Substep 11-6: Go to Step 6.
Step 12: If g = N, set l = l/2.
Step 13: If l < 1, stop the algorithm and output the best chromosome in the current
generation as the result; otherwise, go to Step 5 for the next genetic phase.
After the process of the modified gene-set genetic algorithm with dynamic
mutation rates, a nearly optimal solution to the given problem can be found. Using the
escape operation, the proposed algorithm can help the escape of local optima through
the change of gene-set sizes to a large value. Using dynamic mutation rates, the
proposed algorithm can enlarge the searching region to amend the shortcoming of the
Chapter 9
Experimental Results
This section reports on experiments made to show the performance of the
proposed genetic algorithm. They were implemented in C Language on an AMD K7
2200+ with the Linux OS. Also presented are experiments made to compare the time
required by the proposed genetic algorithm and by the simple genetic algorithm. The
following three problems were used for test:
(A) fA
( ) (
x =−350−x)
2 +500, find the maximum;(B) fB
( ) (
x = x−2.6) (
× −19−x) (
× x+56.5) (
× x−3)
, find the maximum;(C) fC
( )
x = x4×sin(
2xπ)
for 0 < x < 100, find the maximum.The first function is a simple one-peak function with only one local (also global)
optimal solution. The second one has two peaks, and the third one has multiple peaks.
In all the experiments, the chromosome length was 32 and the initial gene-set length
was 4. The termination criterion was the generation number, with each phase being
Experiments were first made for Function A. The problem could actually be
solved by calculus. It was used here to compare the performance of the modified
approach and the others. The population size was set at 100 and the total generation
number was set at 2100 (each phase had 700 generations). When the mutation rates
were set at 0.06, 0.08, 0.1 and 0.32 respectively for the gene-set length of 1, 2, 4 and
16, the relation between fitness values and generations for Function A is shown in
Figure 8, where SGA represents simple GA, HGSGA represents the gene-set SGA
without the escape operation, HGSGA(m) represents the modified HGSGA with the
escape operation, and HGSGA(m2) represents the modified HGSGA with the escape
operation and dynamic mutation rates.
Population size: 100, Mutation rate: 0.06, Function A
-10000 -8000 -6000 -4000 -2000 0 2000 0 300 600 900 1200 1500 1800 2100 2400 generations fi tn es s va lu e
Figure 8: The fitness values along with different generations for Function A.
It can be observed from Figure 4 that the fitness values by all the four
approaches increased along with the increase of generations. It was quite consistent
with the characteristics of GAs. Besides, the gene-set genetic algorithms performed
better than SGA and the proposed gene-set genetic algorithm with dynamic mutation
rates performs best among the four. It can find the global optimal solution very fast.
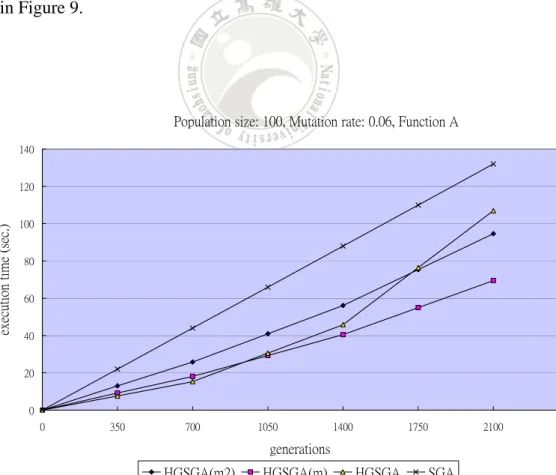
The execution time along with generations by the four algorithms for Function A is
shown in Figure 9.
Population size: 100, Mutation rate: 0.06, Function A
0 20 40 60 80 100 120 140 0 350 700 1050 1400 1750 2100 generations execut ion time ( sec.)
It can be seen from Figure 9 that the gene-set genetic algorithms needed less
computational time than SGA. HGSGA(m) could even spend less computational time
than HGSGA due to the escape operations. HGSGA(m2) spent a little more
computational time than HGSGA(m) since the former might generate more offspring
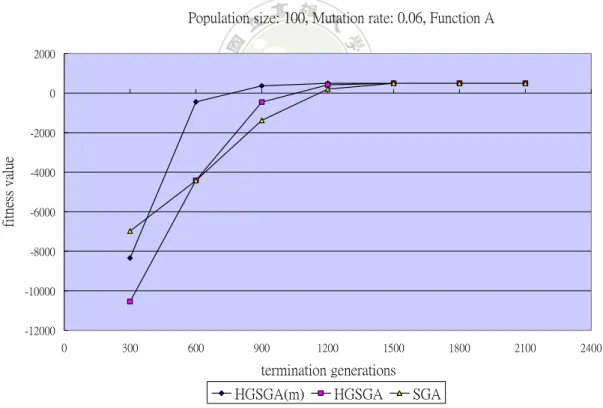
to process. Experiments were then made for showing the relations between fitness
values and termination generations. For mutation rate set at 0.06, the results for
Function A are shown in Figure 10.
Population size: 100, Mutation rate: 0.06, Function A
-12000 -10000 -8000 -6000 -4000 -2000 0 2000 0 300 600 900 1200 1500 1800 2100 2400 termination generations fitne ss va lue
HGSGA(m) HGSGA SGA
Figure 10: The fitness values along with different termination generations for
It can be observed from Figure 10 that the fitness values by all the three
algorithms increased along with the increase of termination generations. When the
number of termination generations was small, convergence could not be achieved.
The gene-set genetic algorithms had a worse effect than SGA since the former ran
only 1/3 of total generations for each phase. But along with the number of termination
generations increased, HGSGA and HGSGA(m) generated better fitness values than
SGA. Besides, HGSGA(m) is better than HGSGA. The execution time along with the
number of termination generations for Function A by the three algorithms is shown in
Population size: 100, Mutation rate: 0.06, Function A 0 20 40 60 80 100 120 140 0 300 600 900 1200 1500 1800 2100 2400 termination generations ex ec utio n tim e (s ec .)
HGSGA(m) HGSGA SGA
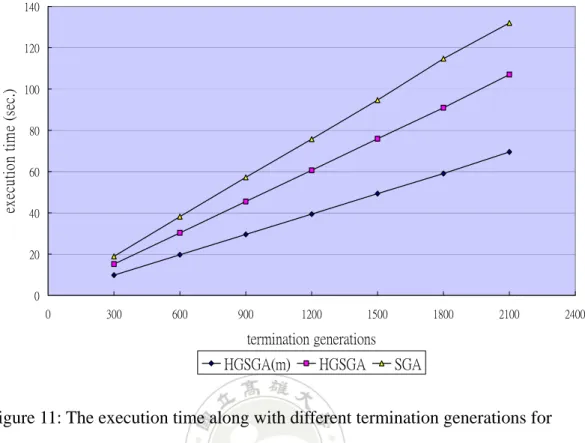
Figure 11: The execution time along with different termination generations for
Function A.
It can be seen from Figure 7 that HGSGA(m) needed the least computational
time among the three algorithms for different termination generations. Next,
experiments were made to show the effect of the mutation rate on the proposed
algorithm. The genetic process was terminated at 600 generations. The final best
fitness values along with different mutation rates for Function A by the three
Population: 100, Generation: 600, Function A -12000 -10000 -8000 -6000 -4000 -2000 0 2000 0 0.02 0.04 0.06 0.08 0.1 0.12 0.14 mutation rate fitn es s v alu e
HGSGA(m) HGSGA SGA
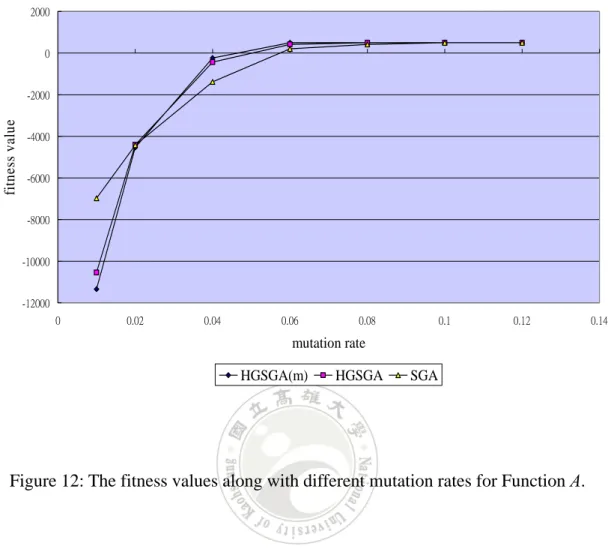
Figure 12: The fitness values along with different mutation rates for Function A.
It can be observed from Figure 12 that the fitness value increased along with the
increase of the mutation rates when the rates where lets then 0.12. Mutation rates were
thus very important to the final results of the algorithms. From the above experimental
results, the modified approach needed a higher number of generations and a higher
mutation rate to achieve a better performance. Experimental results for Function B are
Population size: 100, Mutation rate: 0.06, Function B 400000 450000 500000 550000 600000 650000 700000 0 300 600 900 1200 1500 1800 2100 2400 generations fi tn es s va lu e
HGSGA(m2) HGSGA(m) HGSGA SGA
Population size: 100, Mutation rate: 0.06, Function B 0 20 40 60 80 100 120 140 0 350 700 1050 1400 1750 2100 generations ex ec ut io n ti m e (s ec .)
HGSGA(m2) HGSGA(m) HGSGA SGA
Figure 14: The execution time along with different generations for Function B
Note that in Figure 13, the gene-set sizes in the HGSGA and HGSGA(m) were
long at the beginning generations, which were regarded as a rough optimization phase.
In this phase, the offspring generated by the two gene-set algorithms lacked delicate
local search but focused on big escape movement. The fitness of HGSGA and
HGSGA(m) was thus worse than that of SGA in this phase. Along with the gene-set
size becoming smaller, the local search in HGSGA and HGSGA(m) were performed
more and more delicately and the escape effect was still better than that in simple GA.
help search in the solution space, such that its fitness was better than the other
algorithms in each generation.
Population size: 100, Mutation rate: 0.06, Function B
400000 450000 500000 550000 600000 650000 700000 0 300 600 900 1200 1500 1800 2100 2400 termination generations fi tn es s va lu e
HGSGA(m) HGSGA SGA
Figure 15: The fitness values along with different termination generations for
Population size: 100, Mutation rate: 0.06, Function B 0 20 40 60 80 100 120 140 0 300 600 900 1200 1500 1800 2100 2400 termination generations ex ec ut io n ti m e (s ec. ) HGSGA(m) HGSGA GA
Figure 16: The execution time along with different termination generations for