Andrographolide down-regulates hypoxia-inducible factor-1
α in human non-small
cell lung cancer A549 cells
Hui-Hsuan Lin
a,b, Chia-Wen Tsai
c, Fen-Pi Chou
d, Chau-Jong Wang
b,d, Shu-Wen Hsuan
e,
Cheng-Kun Wang
f, Jing-Hsien Chen
e,⁎
a
School of Medical Laboratory and Biotechnology, Chung Shan Medical University, Taichung, Taiwan
b
Department of Medical Research, Chung Shan Medical University Hospital, Taichung, Taiwan
c
Department of Nutrition, China Medical University, Taichung, Taiwan
d
Institute of Biochemistry and Biotechnology, Chung Shan Medical University, Taichung, Taiwan
eDepartment of Medical Laboratory Science and Biotechnology, College of Medicine and Life Science, Chung Hwa University of Medical Technology, No.89, Wen Hwa 1st St.,
Rende Shiang, Tainan County 717, Taiwan
f
E-Chyun Dermatology Clinic, No.70, Sec. 3, Jhonghua E. Rd., East District, Tainan, Taiwan
a b s t r a c t
a r t i c l e i n f o
Article history: Received 13 September 2010 Revised 16 November 2010 Accepted 25 November 2010 Available online 4 December 2010 Keywords:Andrographolide
Non-small cell lung cancer A549 cell Hypoxia-inducible factor-1α Prolyl hydroxylase
Vascular endothelial growth factor
Andrographolide (Andro), a diterpenoid lactone isolated from a traditional herbal medicine Andrographis paniculata, is known to possess multiple pharmacological activities. In our previous study, Andro had been shown to inhibit non-small cell lung cancer (NSCLC) A549 cell migration and invasion via down-regulation of phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway. Here we demonstrated that Andro inhibited the expression of hypoxia-inducible factor-1α (HIF-1α) in A549 cells. HIF-1α plays an important role in tumor growth, angiogenesis and lymph node metastasis of NSCLC. The Andro-induced decrease of cellular protein level of HIF-1α was correlated with a rapid ubiquitin-dependent degradation of HIF-1α, and was accompanied by increased expressions of hydroxyl-HIF-1α and prolyl hydroxylase (PHD2), and a later decrease of vascular endothelial growth factor (VEGF) upon the treatment of Andro. The Andro-inhibited VEGF expression appeared to be a consequence of HIF-1α inactivation, because its DNA binding activity was suppressed by Andro. Molecular data showed that all these effects of Andro might be mediated via TGFβ1/ PHD2/HIF-1α pathway, as demonstrated by the transfection of TGFβ1 overexpression vector and PHD2 siRNA, and the usage of a pharmacological MG132 inhibitor. Furthermore, we elucidated the involvement of Andro in HIF-1α transduced VEGF expression in A549 cells and other NSCLC cell lines. In conclusion, these results highlighted the potential effects of Andro, which may be developed as a chemotherapeutic or an anti-angiogenesis agent for NSCLC in the future.
© 2010 Elsevier Inc. All rights reserved.
Introduction
Lung cancer is the leading cause of death among cancers worldwide. This high mortality is attributed to its early metastasis, especially for non-small-cell lung carcinoma (NSCLC) (Greenlee et al., 2001). Tumor metastasis occurs by a series of steps including angiogenesis, cell attachment, invasion, and cell proliferation, and is regulated by extremely complicated mechanisms (Fidler, 2005). At this stage, it is impossible to surgically remove all apparent lesions, and this accounts for the high rate of cancer recurrence after surgery;
hence, NSCLC is a prime target in chemotherapy research. Although the mechanism(s) involved in the metastatic progression of NSCLC are still unclear, increasing evidence in cancer therapy literature points to a role of pro-angiogenic factors in the development of lung tumorigenesis, indicating that the hypoxia-inducible factor-1 alpha (HIF-1α) plays an important role in NSCLC development (Choi et al., 2009).
HIF-1α is one of the most important regulatory molecules that respond to hypoxia for cell survival and angiogenesis (Harris, 2002). It is a heterodimer composed of HIF-1α and HIF-1β subunits (Wang and
Semenza, 1995). HIF-1α is metabolized by the ubiquitin-proteasome
pathway under normoxia, but it is stabilized in hypoxia (Salceda and
Caro, 1997). Under conditions of normoxia, post-translational
modifications including the hydroxylation of proline (P564 and P402) residues by prolyl hydroxylases (PHD) within the oxygen-dependent degradation domain (ODDD) promotes HIF-1α interaction with von Hipple–Lindau (pVHL) ubiquitin E3 ligase complex (Ellis et al., 2009; Fong and Takeda, 2008). There are three isoforms of PHD Abbreviations: Andro, andrographolide; NSCLC, non-small cell lung cancer; A549,
human lung cancer cells; HIF-1α, hypoxia-inducible factor-1α; PHD, prolyl hydro-xylases; VEGF, vascular endothelial growth factor; TGFβ1, transforming growth factor beta1; FIH-1, factor-inhibiting HIF-1; TBS, Tris-buffered saline; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis; ECL, enhanced chemilumines-cence; EMSA, electrophoretic mobility shift assay.
⁎ Corresponding author. Fax: +1 886 6 2902371. E-mail address:[email protected](J.-H. Chen).
0041-008X/$– see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.taap.2010.11.014
Contents lists available atScienceDirect
Toxicology and Applied Pharmacology
j o u r n a l h o m e p a g e : w w w. e l s e v i e r. c o m / l o c a t e / y t a a p(PHD1, PHD2 and PHD3) that exist in mammalian systems (Bruick
and McKnight, 2001). Although all three isozymes have potentials to
hydroxylate and thus regulate the HIF-1α level in cultured cells, PHD2 is considered to be the most important one at controlling the levels of HIF-1α during normoxia (Bruick and McKnight, 2001). The expres-sion of PHD2 gene is primarily regulated at the transcriptional through the activation of Smad proteins by transforming growth factor beta1 (TGFβ1) or few stimuli, including hypoxia, insulin, and insulin-like growth factor-1 (McMahon et al., 2006). Another enzyme, asparaginyl hydroxylase factor-inhibiting HIF-1, inhibits the binding of HIF-1α with various co-activators such as p300/CBP by hydroxyl-ating HIF-1α at residue N803, thus resulting in its inactivation (Tal et al., 2008). In solid tumors, HIF-1α is overexpressed by either hypoxic or normoxic stimuli that, in turn, up-regulates the expression of various genes that favor tumor growth and metastasis, and confer resistance to anti-cancer therapies (Yeo et al., 2004).
HIF-1α activates the transcription of many genes, including vascular endothelial growth factor (VEGF) (Semenza, 2000) by binding to the hypoxia response element (HRE) in the VEGF promoter
(Forsythe et al., 1996). Over 100 genes that regulate cellular
processes, such as glucose metabolism, angiogenesis, proliferation, and invasion/metastasis have been found to be targeted by HIF-1α (Rankin and Giaccia, 2008). HIF-1α is related with more aggressive phenotypes and therapeutic resistance of tumor cells. Indeed, several studies have shown that an elevated level of HIF-1α was correlated with radio-resistance and chemo-resistance that led to a poor prognosis (Hockel et al., 1999). Therefore, HIF-1α seems to be a potential target for enhancing the therapeutic efficacy against many tumor cells, including NSCLC.
Andrographis paniculata (Burm. F.) Nees (Acathaceae) is an important herbal medicine widely used in Southeastern Asian countries. The main components of A. paniculata are diterpene lactones of which andrographolide (Andro) is the major component and constitutes 70% of the plant extract (Zhao et al., 2002). Andro has been reported to have multiple pharmacological properties and has been widely used in clinic for the treatment of fever, cold, inflammation, diarrhea and other infectious diseases (Reddy et al.,
2005; Shen et al., 2002). Recent studies suggested that Andro is an
interesting pharmacophore with anti-cancer and immunomodulatory activities, and hence has the potential to be developed as a chemotherapeutic agent (Rajagopal et al., 2003; Srinivasa et al., 2006). The functional mechanisms of Andro could be attributed to its ability to induce cell-cycle arrest (Shi et al., 2008) and trigger apoptosis in human cancer cells (Zhou et al., 2006), and to enhance the secretion of interleukin-2 by cytotoxic T-lymphocytes that inhibits the tumor growth in BALB/c mice (Sheeja and Kuttan, 2007). In addition, Andro suppressed chemotic migration which was associated with inhibition of intracellular protein kinase cascades (Tsai et al., 2004). Our recent study demonstrated the effectiveness of Andro against tumor migration and invasion in human NSCLC A549 cells via down-regulation of PI3K/Akt signaling pathway (Lee et al., 2010).
Considering the importance of overcoming the therapeutic resistance of lung cancer and the possible role of HIF-1α in treatment failure, we investigated whether Andro could modulate the HIF-1α expression in the NSCLC cell line. In addition, the underlying mechanism of this HIF-1α modulation by Andro was examined. Material and methods
Chemicals. Andrographolide (purity 98%), Tris–HCl, EDTA, SDS, phenylmethylsulfonyl fluoride (PMSF), bovine serum albumin (BSA), leupeptin, nonidet p-40, deoxycholic acid, sodium orthovanadate, aprotinin and polyclonal antibody against β-actin were purchased from the Sigma-Aldrich Chemical Co. (St. Louis, MO). Protein assay kits were obtained from Bio-Rad Labs. (Hercules, CA). Dulbecco's Modified Eagle Medium (DMEM), fetal-bovine serum
(FBS), trypsin-EDTA, and penicillin–streptomycin mixed antibiotics were purchased from Gibco/BRL (Gaithersburg, MD). HIF-1α, HIF-1β (Arnt), Ubiquitin (Ub), PHD1, PHD2, PHD3, FIH-1, TGFβ1, Smad and VEGF polyclonal antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). HIF-2α antibody was purchased from Novus Biologicals (Littleton, CO), and Hydroxy-HIF-1α (P564) antibody was from Cell Signaling technology (Danvers, MA). Horse-radish peroxidase-conjugated anti-rabbit and anti-mouse secondary antibodies were purchased from NEN Life Science Products, Inc. (Boston, MA).
Cell culture. Human NSCLC A549, H1355 and H1299 cell lines were obtained from ATCC (Manassas, VA). CL1-0 and its sublines CL1-5 (human lung adenocarcinoma cell lines with different invasive and metastatic capabilities) were obtained from Dr. Min-Liang Kuo (Chang
et al., 2006). All NSCLC cell lines were cultured in growth media
prepared according to ATCC recommendations and supplemented with 10% FBS, 2 mM glutamine and 100 U/ml penicillin–streptomycin mixed antibiotics. All cell cultures were maintained at 37 °C in a humidified atmosphere of 5% CO2.
Preparation of stock solution and treatment. A stock solution of Andro in DMSO at a concentration of 100 mM was prepared and protected from light, and was stored at−20 °C. Before use, Andro solution was freshly prepared by diluting with medium to the desired con-centrations. Control experiments received the same volume of DMSO (final concentration of 0.2%). For experiments with the protease inhibitor, 10μM of MG132 (Sigma Chemical Co., St. Louis, MO, USA) was added 30 min before Andro treatment.
Western blot analysis. Western blotting was performed according to a method described previously (Lin et al., 2007). The preparation of cytosolic and nuclear fractions of the cells was performed as described previously (Lee et al., 2010). The cell lysates were denatured in a sample buffer containing SDS, and equal amounts of protein were separated on 8–15% SDS-poly-acrylamide gels, and, then, transferred to nitrocellulose membranes (Millipore, Bedford, MA, USA). After blocking with 5% nonfat dry milk, the membranes were incubated overnight at 4 °C with the indicated primary antibodies. On the next day, the membranes were incubated with the appropriate horseradish peroxidase-conjugated secondary antibodies, and detected with ECL reagent. The purities of cytosolic fraction were confirmed by Western blotting using anti-β-actin antibody as a loading control. Protein levels were quantitated by densitometry using Alphalmager Series 2200 software.
Real-time quantitative RT-PCR. Total RNA was isolated from cells with a guanidinium chloride procedure as described previously (Chen et al., 2008). The mRNA levels were analyzed by real-time quantitative RT-PCR using a Bio-Rad iCycler system (Bio-Rad, Hercules, CA, USA) (Lee et al., 2010). mRNAs were reverse-transcribed into cDNAs by an iScript cDNA synthesis kit (Bio-Rad). The specificity of primers was tested by running a regular PCR for 40 cycles at 42 °C for 60 min and 65 °C for 15 min followed by electrophoresis on an agarose gel. Real-time PCR was performed using a SYBR supermix kit (Bio-Rad) and run for 40 cycles at 95 °C for 10 s and 57 °C for 15 s, and extension at 72 °C for 20 s. Each 2-μl PCR mixture contained cDNA template, SYBR supermix kit, and 0.5μM of each gene-specific primer. Specific primers were designed using Beacon Designer 2.0 software. The sequences of specific primers for each gene are as follows: 5′-CTGACCCTGCACTCAATCAA-3′ (HIF-1α forward) and 5′-CTTTGCTTCTGTGTCTTCAGC-3′ (HIF-1α reverse), 245 bp; 5′-AAGCCCAGTTTGCTGACATT-3′ (PHD2 forward), and 5′-ATTGAGTTCAACCCTCACAC-3′ (PHD2 reverse), 197 bp; and 5′-AGATCGAGTACATCTTCAAGCCA-3′ (VEGF forward) and 5′-TCTTTCTTTGGTCTGCATTCACAT-3′ (VEGF reverse), 202 bp. The sequences for the house keeping gene, GAPDH, are as follows: 5
′-CGGAGTCAACGGA-TTTGGTCGTAT-3′ (GAPDH forward) and 5′-AGCCTTCTCCATGGTTGGTGAAGAC-3′ (GAPDH reverse), 306 bp. PCR efficiency was examined by serial dilution of the cDNA, and PCR
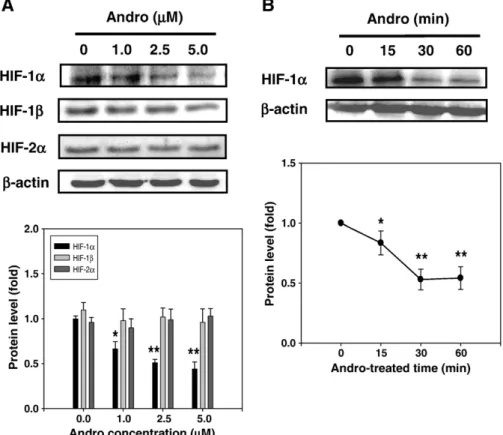
specificity was checked by melting curve data. Each cDNA sample was triplicated and the corresponding no-RT mRNA sample was included as a negative control. The primers of GAPDH were included in every plate Fig. 1. Effects of Andro on the protein level of HIF-1α in A549 cells. (A) A549 cells were treated with various concentrations (0, 1.0, 2.5 and 5.0 μM) of Andro for 60 min or (B) treated with 5.0μM of Andro for indicated times (0, 15, 30 and 60 min), after which cells were harvested and analyzed for the levels of HIF-1α family proteins. Equal amounts of cell lysates (50μg) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific HIF-1α, HIF-1β, HIF-2α and β-actin, served as an internal control, antibodies. The results were represented by using an ECL system, and represent as means ± SD of 3 repeats from three independent experiment. *pb0.05, **pb0.01 compared with the control.
Fig. 2. Effects of Andro on the mRNA level and the proteolytic degradation of HIF-1α in A549 cells. (A) A549 cells were treated with 5.0 μM of Andro for indicated times (0, 15, 30 and 60 min) and then the RNA sample was extracted and subjected to a real-time quantitative RT-PCR for HIF-1α and GAPDH, served as an internal control. (B) Protein extracts prepared from the same treatment condition were immunoprecipitated with HIF-1α. The precipitated complexes were examined for immunoblotting using Ub antibody. The quantitative data were presented as means ± SD of 3 repeats from three independent experiment. *pb0.05, **pb0.01 compared with the control.
to avoid sample variations. The mRNA level of each sample for each gene was normalized to that of GAPDH mRNA.
Immunoprecipitation. Immunoprecipitation assay was carried out as described previously (Chang et al., 2004). Cell lysates were prepared
using the above lysis buffer. Protein (500μg) from cell lysates was pre-cleared with protein A-Sepharose (Amersham Pharmacia Biotech), followed by immunoprecipitation using polyclonal anti-HIF-1αantibody. Immune complexes were harvested with protein A, and immunoprecipitated proteins were analyzed by SDS–PAGE, as above.
Fig. 3. Effect of Andro on the expressions of HIF-1α regulatory proteins in A549 cells. (A, C) A549 cells were treated with various concentrations (0, 1.0, 2.5 and 5.0 μM) of Andro for 60 min or (B, D) treated with 5.0μM of Andro for indicated times (0, 15, 30 and 60 min), after which cells were harvested and analyzed for the protein level of hydroxy-HIF-1α (P564), PHD family and FIH-1. Equal amounts of cell lysates (50μg) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific hydroxy-HIF-1α (A, B), PHD family, including PHD1, PHD2 and PHD3, FIH-1 (C, D) andβ-actin, served as an internal control, antibodies. The results were represented by using an ECL system, and represent as means ± SD of 3 repeats from three independent experiment. *pb0.05, **pb0.01 compared with the control.
Immunodetection was carried out using polyclonal anti-Ub antibody (Santa Cruz Biotech, CA, USA).
Electrophoretic mobility shift assay (EMSA). The binding of HIF-1α protein to a 26 bp DNA fragment (forward: 5 ′-CCACAGTGCATA-CGTGGGCTCCAACA-3′, reverse: 5′-TGTTGGAGCCCACGTATGCAC-TGTGG-3′) derived from the promoter of human VEGF gene with biotin-labeled was assessed by EMSA using the Lightshift kit from Pierce (Rockford, IL, USA) (Chen et al., 2008). The binding reactions containing 10μg of nuclear protein, 10 mM Tris, 50 mM KCl, 1 mM DTT, 5 mM MgCl2, 2μg poly (dI·dC) and 2 pmol of oligonucleotide
probe were incubated at room temperature for 20 min. Specific binding was confirmed by a 200-fold excess of unlabeled probe as specific competitor. Protein DNA complexes were separated on a 6% non-denaturing acrylamide gel, and then transferred to positively charged nylon membranes and linked in a Stratagene cross-linker. Band shifts were visualized with a streptavidin-horseradish peroxidase followed by chemiluminescent detection.
Transient transfection. Transient transfection assay was carried out as described previously (Lin et al., 2007). Liposome-mediated transfection was performed using Lipofectamine™ (Invitrogen) on A549 cells with a control pCMV6-XL4 empty vector or an expression construct for TGFβ1 cDNA in pCMV6-XL4 (Origene Technologies, Rockville, MD, USA), or PHD2 siRNA (Santa Cruz Biotech, CA, USA). Briefly, A549 cells were plated onto six well plates overnight, and transfected with the indicated plasmid at 70–80% confluence on the next day using Lipofectamine reagent according to the manufacturer's instructions. Briefly, lipofectamine (5 μl) and DNA (2 μg) were diluted in 100μl of DMEM followed by equilibration at room temperature for 5–10 min after mixing. The lipofectamine–DNA complex was added to A549 cells, and incubated for 12 h. Cells were then washed with PBS and replenished with DMEM containing 20% serum. At 12 h after transfection, the cells were incubated with 5.0μM of Andro for 60 min, and, then, expanded for further studies.
Statistical analysis. Results were reported as means ± SD, and statistical analysis was obtained using an unpaired t-test. A value of pb0.05 was considered statistically significant.
Result
Effects of Andro on the protein level of HIF-1α in A549 cells
To investigate whether Andro could affect the expression of HIF-1α in A549 cells, wefirst examined the cellular levels of HIF family proteins after the treatment of Andro by Western blotting. As shown inFig. 1A, incubation of A549 cells with Andro (60 min) led to a dose-dependent decreased level of HIF-1α, but not HIF-1β or HIF-2α. The protein level of HIF-1α was reduced to 0.5-fold of control level (p≤0.01) after 1.0 to 5.0μM of Andro addition. Time-course (0, 15, 30 and 60 min) study showed a significant reduction in HIF-1α level started at 15 min and persisted to 60 min after the exposure of 5.0μM of Andro in A549 cells (Fig. 1B).
Effects of Andro on the proteolytic degradation of HIF-1α in A549 cells To address the mechanism by which Andro decreased the protein level of HIF-1α, we further examined HIF-1α mRNA level. The data in
Fig. 2A demonstrated that no significant change in HIF-1α mRNA level was observed in A549 cells after 15 to 60 min of Andro incubation. These results indicated that the Andro-induced decrease in HIF-1α protein level involved a post-transcriptional mechanism.
To determine whether HIF-1α degradation was accelerated by Andro, we treated A549 cells with 5.0μM of Andro for the indicated time (0, 15, 30, and 60 min), and then measured the formation
of HIF-1α/Ub complex in the cells. As shown inFig. 2B, the level of HIF-1α/Ub complex increased faster in the cells treated with Andro than that in the untreated cells (data not shown). This result suggested that a post-translational mechanism contributed to the Andro-induced HIF-1α depletion in A549 cells.
Effects of Andro on the expressions of HIF-1α regulatory proteins in A549 cells
It is well known that by controlling the ubiquitin-proteasome pathway of HIF-1α, prolyl hydroxylation of HIF-1α is critical in the regulation of the steady state levels of HIF-1α (Ivan et al., 2001; Jaakkola et al., 2001). Immunoblot analysis showed that the treatment of A549 cells with 5.0μM of Andro (60 min) resulted in an about 4-fold increase
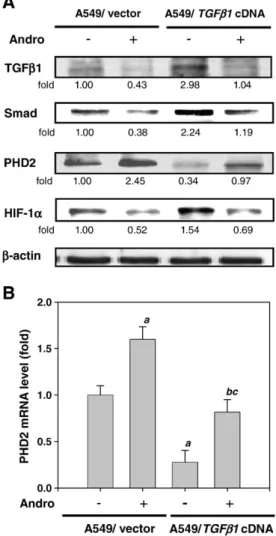
Fig. 4. Effects of mutant TGFβ1 expression vector on Andro-mediated PHD2/HIF-1α signaling pathway. A549 cells were transfected with empty vector or TGFβ1 cDNA (activated) and treated with or without 5.0μM of Andro for 60 min. (A) The protein levels of TGFβ1, Smad, PHD2 and HIF-1α were analyzed by Western blot. β-Actin was served as an internal control. Determined expression of these proteins was subsequently quantified by densitometric analysis with that of control being 1.00 fold as shown just below the gel data. Results from three repeated and separated experiments were similar. (B) The mRNA level of PHD2 was analyzed by real-time quantitative RT-PCR. The quantitative data were presented as means ± SD of 3 repeats from three independent experiment.apb0.01 compared with the A549/vector treated
without Andro group.bpb0.01 compared with the A549/ TGFβ1 cDNA treated with
Andro group compared with A549/TGFβ1 cDNA treated without Andro group.c
pb0.01 the A549/TGFβ1 cDNA treated with Andro group compared with A549/vector treated with Andro group.
in the amount of hydroxy-HIF-1α (P564) (Fig. 3A and B). Next, tofind out the specific HIF-1α regulatory proteins responsible for the proteolytic degradation of HIF-1α following Andro treatment, Western blot analysis using antibodies specific to PHD1, PHD2, PHD3 and FIH-1 was performed (Fig. 3C and D). A dramatic increase in the PHD2 protein level was observed at 15 min after Andro treatment that reached the maximum level at 60 min (Fig. 3D), whereas PHD1, PHD3 and FIH-1 remained unchanged (Fig. 3C). Therefore, Andro stimulated a concen-tration- and time-dependent increase in the expressions of hydroxy-HIF-1α (P564) and PHD2 in A549 cells.
Effects of Andro on TGFβ1/PHD2/HIF-1α signaling pathway in A549 cells Very little information is available on the molecular control of PHD2 expression. Previous study has revealed that TGFβ1 decreased PHD2 gene transcription levels through the activation of Smad proteins (McMahon et al., 2006). To confirm whether the TGFβ1 protein involved in Andro-regulated PHD2/HIF-1α signaling, we used a genetic approach to over-express TGFβ1 in A549 cells. The Western blotting results showed that the cells expressed a control vector indeed had diminished levels of TGFβ1, Smad and HIF-1α when cells
Fig. 5. Effect of Andro on the expression of VEGF and the VEGF DNA binding activities of HIF-1α in A549 cells. (A) A549 cells were treated with various concentrations (0, 1.0, 2.5 and 5.0μM) of Andro for 60 min or (B) treated with 5.0 μM of Andro for indicated times (0, 15, 30 and 60 min), after which cells were harvested and analyzed for the protein level of VEGF. Equal amounts of cell lysates (50μg) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific VEGF and β-actin, served as an internal control, antibodies. (C) Real-time quantitative RT-PCR analysis of the mRNA level of VEGF in cells treated with 5.0μM of Andro and harvested at the indicated times. The quantitative data were presented as means ± SD of 3 repeats from three independent experiment. *pb0.05, **pb0.01 compared with the control. (D) Cells were treated with 5.0 μM of Andro for indicated times and then nuclear extracts were analyzed for the DNA binding activity of HIF-1α using biotin labeled HIF-1α specific oligonucleotide by EMSA. Lane 1 represented nuclear extracts incubated with unlabeled oligonucleotide (free probe) to confirm the specificity of binding. Results from three repeated and separated experiments were similar.
were treated with Andro (lines 1, 2 and 4,Fig. 4A). This suppressive effect of Andro was improved by an atypical overexpression of TGFβ1 (lanes 2 and 4,Fig. 4A). The expression of constitutively active TGFβ1 also abrogated the increase levels of PHD2 protein and mRNA that was originally induced by Andro as analyzed by Western blotting (line 3,
Fig. 4A) and real-time quantitative RT-PCR assay (Fig. 4B).
Effects of Andro on the expression of VEGF and the VEGF DNA binding activities of HIF-1α in A549 cells
Because HIF-1α activates the expression of VEGF gene by binding to the hypoxia response element (HRE) in the VEGF promoter region
(Forsythe et al., 1996), Western blot analysis was performed to
investigate the effect of Andro on the VEGF expression. We found that A549 cells exposed to 2.5 and 5.0μM of Andro for 60 min demonstrated a ~40% and 50% decrease in the protein level of VEGF, respectively
(Fig. 5A). Time course experiments with 5.0μM of Andro revealed a
significant and prolonged inhibition of VEGF protein expression
(Fig. 5B). The Andro-induced decrease in the protein level of VEGF
coincided well with its mRNA level as evidenced by the result of real-time quantitative RT-PCR (Fig. 5C), indicating that Andro might regulate VEGF expressions at the transcriptional level.
Next, a DNA probe derived from the promoter of VEGF gene containing a HIF-1α binding site was used for EMSA analysis to assess the binding activity of HIF-1α in the nuclear extracts prepared from the cells treated with or without Andro. The result showed that Andro significantly decreased the HIF-1α binding activity to the DNA probe in a time-dependent manner (Fig. 5D).
Effects of a 26S proteasome inhibitor and PHD2 siRNA on Andro-regulated HIF-1α signaling
To determine whether Andro-reduced the protein level of HIF-1α is due to an effect of degradation, we studied the regulation of a 26S proteasome inhibitor, MG132, in the presence of Andro. As shown in
Fig. 6, A549 cells with either MG132 alone (lane 3) or control cells without treatment of Andro (lane 1) showed a reduction in the HIF-1α/
Ub complex concomitantly with a marked induction on HIF-1α protein level, whereas treatment with MG132 completely inhibited Andro-induced the proteolytic degradation of HIF-1α (lane 4). These results suggested that ubiquitin proteasome system was involved in the inactivation of the HIF-1α that transduced VEGF expression in A549 cells by Andro.
To further determine whether the observed increase of PHD2 protein levels is responsible for the proteolytic degradation of HIF-1α by Andro, we used a genetic approach to deplete endogenous PHD2 using PHD2-specific siRNA. As presented inFig. 6(lanes 5 and 6), an increase in the HIF-1α/Ub complex and the decrease of HIF-1α and VEGF protein levels in response to Andro were specifically reversed by PHD2 siRNA. These results, which are in line with previous data demonstrating that PHD2 is critical for controlling the low steady state levels of HIF-1α in normoxia (Berra et al., 2003), designate the inhibition of PHD2 expression levels as the mechanism underlying Andro-reduced HIF-1α stabilization.
Effect of Andro on PHD2/HIF-1α signaling pathway in a panel of NSCLC cell lines
To investigate the implications of PHD2/HIF-1α signaling on Andro-response, we characterized the panel of NSCLC tumor cell lines. Treatment of NSCLC cell lines, H1355, H1299, CL1-0 and CL1-5, with Andro (5.0μM) effectively inhibited the protein levels of HIF-1α and VEGF, and increased that of PHD2 (Fig. 7A). Hence, we confirmed that HIF-1α and VEGF were highly expressed in all NSCLC cell lines and the expression of these proteins was found to be correlated with tumor malignant. These similarities suggested that Andro is indeed capable of decreasing HIF-1α protein half-life through induction of PHD2 expression in NSCLC cells.
Discussion
Andrographolide (Andro) is diterpene lactone that has been shown to possess anti-tumor properties, and therefore acquires special interest for the development of a novel chemotherapeutic agent for cancer. In in vitro studies, Andro inhibited cancer cell proliferation (Satyanarayana et al., 2004), induced cell-cycle arrest (Shi et al., 2008) and promoted apoptosis (Zhou et al., 2006) in human cancer cells. The pro-apoptotic effect of Andro was shown to be mediated by inducing mitochondrial cytochrome c release that was accompanied by an increased expression level of Bax and a decrease in Bcl-2 protein in human leukemia HL-60 cells (Cheung et al., 2005), and by causing apoptotic morphological change and activations of caspases 3 and 8 in human prostate carcinoma PC-3 cells (Kim and Milner, 2005). Our previous study also suggested that Andro arrested cell-cycle at G1-S phase via CKI-cyclin-Cdk network in human colorectal carcinoma Lovo cells (Shi et al., 2008). In addition, Andro inhibited chemotic migration which was associated with inhibition of ERK1/2 and Akt protein kinase cascades (Tsai et al., 2004). Because of its suppressive effect on PI3K/Akt/AP-1, Andro might, in turn, have a significant impact on the mechanism and lead to an inhibition of the MMP-mediated cellular events in non-small-cell lung cancer (NSCLC) A549 cells (Lee et al., 2010). However, the molecular mechanism underlying the Andro-mediated inhibition of NSCLC tumorigenesis has not been elucidated until present study showing that the down-regulation of HIF-1α signaling pathway might be responsible for the inhibitory effect of Andro.
NSCLC is the most prevalent type of lung cancer. Despite treatment advances in surgery, chemotherapy, and radiotherapy, the overall survival rate is poor. Thus, the search for an effective chemothera-peutic approach is important to NSCLC treatment (Liu et al., 2005). A growing body of evidence implies activation of HIF-1α pathway as a critical step in carcinogenesis due to its intermediate link in the evolution of the tumor, and regulation of a variety of gene expression Fig. 6. PHD2 and the ubiquitin proteasome system are essential for the HIF-1α
signaling regulated by Andro. A549 cells were pretreated with MG132 for 30 min or transfected with PHD2 siRNA, and then treated with 5.0μM of Andro for 60 min. Protein extracts were immunoprecipitated with HIF-1α, and the precipitated complexes were examined for immunoblotting using Ub antibody (line 1). The protein levels of HIF-1α, PHD2 and VEGF were analyzed by Western blot (lines 2–4). β-Actin was served as an internal control. Determined expression of these proteins was subsequently quantified by densitometric analysis with that of control being 1.00 fold as shown just below the gel data. Results from three repeated and separated experiments were similar.
(Chen et al., 2009; Teicher, 1994). The expression of HIF-1α is related to differentiation, metastasis and clinical stage of lung cancer (Bos et al., 2003). Elevated HIF-1α expression also has been reported in NSCLC tissues with poor disease prognosis (Zhang et al., 2007). In this study, we confirmed that HIF-1α is highly expressed in this NSCLC cell line. To our knowledge, we showed for thefirst time that treatment with Andro decreased HIF-1α protein accumulation (Fig. 1), which blocked, at least in part, the expression of angiogenic factors, such as VEGF (Fig. 5), in human NSCLC cell lines (Fig. 7A). Therefore, these findings have provided novel mechanisms of biological functions of Andro in the inhibition of NSCLC tumorigenesis.
The results of the present study indicated that the ubiquitin-proteasome pathway was involved in regulating the turnover of
HIF-1α in A549 cells, and that Andro decreased HIF-1α stability by post-translationally enhancing the proteasome-dependent degrada-tion of the protein (Fig. 2), as demonstrated by the usage of a 26S proteasome inhibitor, MG132 (Fig. 6). The four members of hydro-xylases (asparaginyl hydroxylase FIH-1 and three prolyl hydrohydro-xylases PHD1–3) have been demonstrated to hydroxylate HIF-1α sequences in vitro and to down-regulate the hypoxia-responsive reporter gene activity in forced expression studies (Schofield and Ratcliffe, 2004). Thus, we reasoned that Andro might affect FIH-1 and PHD family that, in turn, has been shown to control the degradation of the HIF-1α. We demonstrated here that Andro enhanced the protein level of PHD2 in A549 cells, but not other PHD isoforms and FIH-1 (Fig. 3). Thefinding raised an important question as to the contribution of individual Fig. 7. (A) Effect of Andro on PHD2/HIF-1α signaling pathway in a panel of NSCLC cell lines. Five kinds of cell lines were treated with or without 5.0 μM of Andro for 60 min, after which cells were harvested and analyzed for the protein levels of HIF-1α, PHD2 and VEGF. Equal amounts of cell lysates (50 μg) were resolved by SDS-PAGE, transferred to nitrocellulose, and probed with specific HIF-1α, PHD2, VEGF and β-actin, served as internal control, antibodies. Results are representative of at least three independent experiments. *pb0.05, **pb0.01 compared with the untreated Andro sample of 5 kinds of cell lines, respectively. (B) A proposed model for the inhibitory effect of Andro on HIF-1α signaling in human NSCLC cells. See the text for discussion.
enzyme in the physiological regulation of HIF-1α. Anyhow, in the present study, a clear decrease in HIF-1α expression was found in the Andro-treated A549 cells, in concert with a reciprocal increase in hydroxy-HIF-1α and PHD2 expression.
Little is known about the cellular aspects controlling PHD2 expression. Only few stimuli, including hypoxia, insulin, and insulin-like growth factor-1, were shown (in contrast to TGFβ1) to increase PHD2 expression through transcriptional regulation (Aprelikova et al.,
2004; McMahon et al., 2006; Metzen et al., 2005). Previous studies
have revealed that TGFβ1 decreased PHD2 gene transcription levels through the activation of Smad proteins. This signaling pathway seems to be central to the TGFβ1 effect, because its inhibition restored PHD2 expression and impaired HIF-1α protein accumulation in response to the growth factor (McMahon et al., 2006). Consistent with this possibility, A549 cells transfected with TGFβ1 cDNA (active form) showed an increase in TGFβ1 protein level and was accompa-nied by increased expressions of Smad and HIF-1α. This TGFβ1 activation resulted in the decreased protein and mRNA levels of PHD2
(Fig. 4). Our preliminary evidences demonstrated that efforts are
presently deployed to address the possible role of Andro-induced TGFβ1/Smad inhibition in PHD2 regulation. However, the mecha-nisms involved are likely pathway and have therefore not been fully characterized. Our future work will focus on defining the mechanism by which Andro affects TGFβ1 levels. Furthermore, because TGFβ1 is abundantly and widely expressed in NSCLC cells, and in view of recent advances indicating that this growth factor is a key in the promotion of both tumor growth and inflammation (Gu et al., 2007; McMahon et al., 2006), it appears that agents, such as Andro, that counteract or overcome PHD2 inhibition may provide a therapeutic advantage.
The process of HIF-1α stabilization and activation can be affected by the major signal transduction pathways, including the ERK and PI3K/Akt cascades (Minet et al., 2000; Zhong et al., 2000). Therefore, one possibility that would account for the decreased levels of HIF-1α protein is the decreased PI3K/Akt signaling in Andro-treated A549 cells. Studies of the role of PI3K signaling in HIF-1α expression were contradictory based on the cell lines used. PI3K and Akt activities were observed to be required for HIF-1α expression in prostate cancer cells (Yin et al., 2001; Zhong et al., 2000), while its inhibition in 1c1c7 mouse hepatocyte cells did not affect HIF-1α expression (Arsham et al., 2002). In ovarian carcinoma, the catalytic unit of PI3K is directly implicated in the control of HIF-1α protein and VEGF expression (Zhang et al., 2003). Our unpublished data revealed that blockage of PI3K/Akt signaling by 5.0μM of Andro (24 h) significantly suppressed HIF-1α protein and mRNA levels, VEGF transcriptional activation, and VEGF protein levels in A549 cells (data not shown). All these suggest that PI3K/Akt signaling may account for the expression of HIF-1α in lung cancer cells. However, the data of present study showed that treatment with 5.0μM of Andro for 60 min enhanced the proteolytic degradation of HIF-1α without affecting its mRNA levels in the short term. We therefore suggested that the influence of Andro on HIF-1α expression might be differential depending on the length of exposure, and that the PI3K/Akt signaling might be responsible for long-term regulation of HIF-1α. Further studies need to be conducted to clarify the detail pathway of Andro's action.
There are a number of reported HIF-1α-binding sites in the promoter region of VEGF gene, and indeed VEGF expression was up-regulated by HIF-1α binding to the promoter in cancer cells (Forsythe et al., 1996; Shima et al., 1996). Consistent with thesefindings, we observed that the decreased protein and mRNA levels of VEGF were coincided with an inhibitory effect on the binding activity of HIF-1α to VEGF promoter upon the treatment of Andro (Fig. 5). Overexpression of VEGF mediated by the stabilization of HIF-1α has been identified in multiple malignancies and for this reason targeting the tumor vasculature via the inhibition of VEGF either directly or indirectly has become an attractive target in novel anti-cancer drug develop-ment (Ellis et al., 2009). In this study, the involvement of PHD2 signal
pathway in the effect of Andro on the HIF-1α degradation was further confirmed in the experiments using PHD2 siRNA (Fig. 6), implying that inhibition of PHD2 expression levels could increase HIF-1α protein half-life concomitantly with a marked induction on VEGF protein level. Our genetic evidences demonstrated that PHD2/HIF-1α signaling played a crucial role in the suppressive effect of Andro on VEGF expression in A549 cells.
Based on these results, we proposed a schematic presentation of possible mechanisms for the effects of Andro on HIF-1α signaling in A549 cells, and other NSCLC cell lines (Fig. 7A). These results clearly demonstrated that Andro regulated the tumor angiogenetic TGFβ1/ PHD2/HIF-1α/VEGF pathway (Fig. 7B). Through our experiments, a link emerged between PHD2/HIF-1α signaling and TGFβ1, which can be regulated by Andro. We showed that Andro enhanced the proteolytic degradation of HIF-1α through PHD2 signaling pathways that subsequently inhibited the expression of VEGF at the transcrip-tional level. Thesefindings revealed a novel role of Andro in inhibiting HIF-1α and VEGF expression that may be important for lung tumor angiogenesis and metastasis, and identified detail signaling molecules that mediate this regulation. Molecular targeting of the HIF-1α or VEGF by Andro may be a useful and novel strategy for chemopreven-tion and/or treatment of NSCLC.
Conflict of interest statement
The authors declare that there are no conflicts of interest. Acknowledgment
This work was supported by the grant from the National Science Council, Taiwan (NSC 97-2313-B-273-002).
References
Aprelikova, O., Chandramouli, G.V., Wood, M., Vasselli, J.R., Riss, J., Maranchie, J.K., Linehan, W.M., Barrett, J.C., 2004. Regulation of HIF prolyl hydroxylases by hypoxia-inducible factors. J. Cell. Biochem. 92, 491–501.
Arsham, A.M., Plas, D.R., Thompson, C.B., Simon, M.C., 2002. Phosphatidylinositol 3-kinase/Akt signaling is neither required for hypoxic stabilization of HIF-1 alpha nor sufficient for HIF-1-dependent target gene transcription. J. Biol. Chem. 277, 15162–15170.
Berra, E., Benizri, E., Ginouvès, A., Volmat, V., Roux, D., Pouysségur, J., 2003. HIF prolyl-hydroxylase 2 is the key oxygen sensor setting low steady-state levels of HIF-1alpha in normoxia. EMBO J. 22, 4082–4090.
Bos, R., van der Groep, P., Greijer, A.E., Shvarts, A., Meijer, S., Pinedo, H.M., Semenza, G.L., van Diest, P.J., van der Wall, E., 2003. Levels of hypoxia-inducible factor-1alpha independently predict prognosis in patients with lymph node negative breast carcinoma. Cancer 97, 1573–1581.
Bruick, R.K., McKnight, S.L., 2001. A conserved family of prolyl-4-hydroxylases that modify HIF. Science 294, 1337–1340.
Chang, Y.C., Chou, F.P., Huang, H.P., Hsu, J.D., Wang, C.J., 2004. Inhibition of cell cycle progression by penta-acetyl geniposide in rat C6 glioma cells. Toxicol. Appl. Pharmacol. 198, 11–20.
Chang, C.C., Lin, M.T., Lin, B.R., Jeng, Y.M., Chen, S.T., Chu, C.Y., Chen, R.J., Chang, K.J., Yang, P.C., Kuo, M.L., 2006. Effect of connective tissue growth factor on hypoxia-inducible factor 1alpha degradation and tumor angiogenesis. J. Natl Cancer Inst. 98, 984–995.
Chen, J.H., Lin, H.H., Chiang, T.A., Hsu, J.D., Ho, H.H., Lee, Y.C., Wang, C.J., 2008. Gaseous nitrogen oxide promotes human lung cancer cell line A549 migration, invasion, and metastasis via iNOS-mediated MMP-2 production. Toxicol. Sci. 106, 364–375. Chen, Y.Q., Zhao, C.L., Li, W., 2009. Effect of hypoxia-inducible factor-1alpha on
transcription of survivin in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 28, 29.
Cheung, H.Y., Cheung, S.H., Li, J., Cheung, C.S., Lai, W.P., Fong, W.F., Leung, F.M., 2005. Andrographolide isolated from Andrographis paniculata induces cell cycle arrest and mitochondrial-mediated apoptosis in human leukemic HL-60 cells. Planta Med. 71, 1106–1111.
Choi, Y.J., Rho, J.K., Lee, S.J., Jang, W.S., Lee, S.S., Kim, C.H., Lee, J.C., 2009. HIF-1alpha modulation by topoisomerase inhibitors in non-small cell lung cancer cell lines. J. Cancer Res. Clin. Oncol. 135, 1047–1053.
Ellis, L., Hammers, H., Pili, R., 2009. Targeting tumor angiogenesis with histone deacetylase inhibitors. Cancer Lett. 280, 145–153.
Fidler, I.J., 2005. The organ microenvironment and cancer metastasis. Differentiation 70, 498–505.
Fong, G.H., Takeda, K., 2008. Role and regulation of prolyl hydroxylase domain proteins. Cell Death Differ. 15, 635–641.
Forsythe, J.A., Jiang, B.H., Iyer, N.V., Agani, F., Leung, S.W., Koos, R.D., Semenza, G.L., 1996. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol. Cell. Biol. 16, 4604–4613.
Greenlee, R.T., Hill-Harmon, M.B., Murray, T., Thun, M., 2001. Cancer statistics, 2001. CA Cancer J. Clin. 51, 15–36.
Gu, L., Zhu, Y.J., Yang, X., Guo, Z.J., Xu, W.B., Tian, X.L., 2007. Effect of TGF-beta/Smad signaling pathway on lung myofibroblast differentiation. Acta Pharmacol. Sin. 28, 382–391.
Harris, A.L., 2002. Hypoxia—a key regulatory factor in tumour growth. Nat. Rev. Cancer 2, 38–47.
Hockel, M., Schlenger, K., Hockel, S., Vaupel, P., 1999. Hypoxic cervical cancers with low apoptotic index are highly aggressive. Cancer Res. 59, 4525–4528.
Ivan, M., Kondo, K., Yang, H., Kim, W., Valiando, J., Ohh, M., Salic, A., Asara, J.M., Lane, W. S., Kaelin Jr., W.G., 2001. HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science 292, 464–468. Jaakkola, P., Mole, D.R., Tian, Y.M., Wilson, M.I., Gielbert, J., Gaskell, S.J., Kriegsheim, A.,
Hebestreit, H.F., Mukherji, M., Schofield, C.J., Maxwell, P.H., Pugh, C.W., Ratcliffe, P.J., 2001. Targeting of HIF-alpha to the von Hippel–Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 292, 468–472.
Kim, Y.S., Milner, J.A., 2005. Targets for indole-3-carbinol in cancer prevention. J. Nutr. Biochem. 16, 65–73.
Lee, Y.C., Lin, H.H., Hsu, C.H., Wang, C.J., Chiang, T.A., Chen, J.H., 2010. Inhibitory effects of andrographolide on migration and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. Eur. J. Pharmacol. 632, 23–32.
Lin, H.H., Chen, J.H., Huang, C.C., Wang, C.J., 2007. Apoptotic effect of 3, 4-dihydroxybenzoic acid on human gastric carcinoma cells involving JNK/p38 MAPK signaling activation. Int. J. Cancer 120, 2306–2316.
Liu, L.Z., Fang, J., Zhou, Q., Hu, X., Shi, X., Jiang, B.H., 2005. Apigenin inhibits expression of vascular endothelial growth factor and angiogenesis in human lung cancer cells: implication of chemoprevention of lung cancer. Mol. Pharmacol. 68, 635–643. McMahon, S., Charbonneau, M., Grandmont, S., Richard, D.E., Dubois, C.M., 2006.
Transforming growth factor beta1 induces hypoxia-inducible factor-1 stabilization through selective inhibition of PHD2 expression. J. Biol. Chem. 281, 24171–24181. Metzen, E., Stiehl, D.P., Doege, K., Marxsen, J.H., Hellwig-Bürgel, T., Jelkmann, W., 2005. Regulation of the prolyl hydroxylase domain protein 2 (phd2/egln-1) gene: identification of a functional hypoxia-responsive element. Biochem. J. 387, 711–717. Minet, E., Arnould, T., Michel, G., Roland, I., Mottet, D., Raes, M., Remacle, J., Michiels, C., 2000. ERK activation upon hypoxia: involvement in HIF-1 activation. FEBS Lett. 468, 53–58. Rajagopal, S., Ajaya, K.R., Devi, D.S., Satyanarayana, C., Rajagopalan, R., 2003. Andrographolide, a potential cancer therapeutic agent isolated from Andrographis paniculata. J. Exp. Ther. Oncol. 3, 147–158.
Rankin, E.B., Giaccia, A.J., 2008. The role of hypoxia-inducible factors in tumorigenesis. Cell Death Differ. 15, 678–685.
Reddy, V.L., Reddy, S.M., Ravikanth, V., Krishnaiah, P., Goud, T.V., Rao, T.P., Ram, T.S., Gonnade, R.G., Bhadbhade, M., Venkateswarlu, Y., 2005. A new bis-andrographolide ether from Andrographis paniculata nees and evaluation of anti-HIV activity. Nat. Prod. Res. 19, 223–230.
Salceda, S., Caro, J., 1997. Hypoxia-inducible factor 1alpha (HIF-1alpha) protein is rapidly degraded by the ubiquitin-proteasome system under normoxic conditions. Its stabilization by hypoxia depends on redox-induced changes. J. Biol. Chem. 272, 22642–22647.
Satyanarayana, C., Deevi, D.S., Rajagopalan, R., Srinivas, N., Rajagopal, S., 2004. DRF 3188 a novel semi-synthetic analog of andrographolide: cellular response to MCF 7 breast cancer cells. BMC Cancer 4, 26–33.
Schofield, C.J., Ratcliffe, P.J., 2004. Oxygen sensing by HIF hydroxylases. Nat. Rev. Mol. Cell Biol. 5, 343–354.
Semenza, G.L., 2000. Hypoxia, clonal selection, and the role of HIF-1 in tumor progression. Crit. Rev. Biochem. Mol. Biol. 35, 71–103.
Sheeja, K., Kuttan, G., 2007. Activation of cytotoxic T lymphocyte responses and attenuation of tumor growth in vivo by Andrographis paniculata extract and andrographolide. Immunopharmacol. Immunotoxicol. 29, 81–93.
Shen, Y.C., Chen, C.F., Chiou, W.F., 2002. Andrographolide prevents oxygen radical production by human neutrophils: possible mechanism(s) involved in its anti-inflammatory effect. Br. J. Pharmacol. 135, 399–406.
Shi, M.D., Lin, H.H., Lee, Y.C., Chao, J.K., Lin, R.A., Chen, J.H., 2008. Inhibition of cell-cycle progression in human colorectal carcinoma Lovo cells by andrographolide. Chem. Biol. Interact. 174, 201–210.
Shima, D.T., Kuroki, M., Deutsch, U., Ng, Y.S., Adamis, A.P., D'Amore, P.A., 1996. The mouse gene for vascular endothelial growth factor. Genomic structure, definition of the transcriptional unit, and characterization of transcriptional and post-transcrip-tional regulatory sequences. J. Biol. Chem. 271, 3877–3883.
Srinivasa, R.J., Hamzah, A.S., Saad, M.S., Lajis, N.H., Stevens, M.F.G., Stanslas, J., 2006. Semisynthesis and cytotoxic activities of andrographolide analogues. J. Enzyme Inhib. Med. Chem. 21, 145–155.
Tal, R., Shaish, A., Bangio, L., Peled, M., Breitbart, E., Harats, D., 2008. Activation of C-transactivation domain is essential for optimal HIF-1 alpha-mediated transcrip-tional and angiogenic effects. Microvasc. Res. 76, 1–6.
Teicher, B.A., 1994. Hypoxia and drug resistance. Cancer Metastasis Rev. 13, 139–168. Tsai, H.R., Yang, L.M., Tsai, W.J., Chiou, W.F., 2004. Andrographolide acts through inhibition of ERK1/2 and Akt phosphorylation to suppress chemotactic migration. Eur. J. Pharmacol. 498, 45–52.
Wang, G.L., Semenza, G.L., 1995. Purification and characterization of hypoxia-inducible factor 1. J. Biol. Chem. 270, 1230–1237.
Yeo, E.J., Chun, Y.S., Park, J.W., 2004. New anticancer strategies targeting HIF-1. Biochem. Pharmacol. 68, 1061–1069.
Yin, F., Giuliano, A.E., Law, R.E., Van Herle, A.J., 2001. Apigenin inhibits growth and induces G2/M arrest by modulating cyclin-CDK regulators and ERK MAP kinase activation in breast carcinoma cells. Anticancer Res. 21, 413–420.
Zhang, L., Yang, N., Katsaros, D., Huang, W., Park, J.W., Fracchioli, S., Vezzani, C., Rigault de la Longrais, I.A., Yao, W., Rubin, S.C., Coukos, G., 2003. The oncogene phosphatidylinositol 3′-kinase catalytic subunit alpha promotes angiogenesis via vascular endothelial growth factor in ovarian carcinoma. Cancer Res. 63, 4225–4231.
Zhang, Q., Tang, X., Zhang, Z.F., Velikina, R., Shi, S., Le, A.D., 2007. Nicotine induces hypoxia-inducible factor-1alpha expression in human lung cancer cells via nicotinic acetylcholine receptor-mediated signaling pathways. Clin. Cancer Res. 13, 4686–4694.
Zhao, J., Yang, G., Liu, H., Wang, D., Song, X., Chen, Y., 2002. Determination of andrographolide, deoxyandrographolide and neoandrographolide in the Chinese herb Andrographis paniculata by micellar electrokinetic capillary chromatography. Phytochem. Anal. 13, 222–227.
Zhong, H., Chiles, K., Feldser, D., Laughner, E., Hanrahan, C., Georgescu, M.M., Simons, J.W., Semenza, G.L., 2000. Modulation of hypoxiainducible factor 1alpha expression by the epidermal growth factor/ phosphatidylinositol 3-kinase/PTEN/AKT/FRAP pathway in human prostate cancer cells: implications for tumor angiogenesis and therapeutics. Cancer Res. 60, 1541–1545.
Zhou, J., Zhang, S., Ong, C.N., Shen, H.M., 2006. Critical role of pro-apoptotic Bcl-2 family members in andrographolide-induced apoptosis in human cancer cells. Biochem. Pharmacol. 72, 132–144.