Full-length article
Tetramethylpyrazine suppresses HIF-1
α, TNF-α, and activated caspase-3
expression in middle cerebral artery occlusion-induced brain ischemia
in rats
1Yi CHANG2,3, George HSIAO4, Seu-hwa CHEN4, Yi-cheng CHEN4, Jiing-han LIN4, Kuang-hung LIN4, Duen-suey CHOU4,
Joen-rong SHEU4,5
2Department of Anesthesiology, Shin Kong Wu Ho-Su Memorial Hospital, Taipei 110, Taiwan, China; 3School of Medicine, Fu-Jen Catholic
University, Taipei, 110, Taiwan, China; 4Graduate Institute of Medical Sciences, Taipei Medical University, Taipei 110, Taiwan, China
Abstract
Aim: To examine the detailed mechanisms underlying the inhibitory effect of tetramethylpyrazine (TMPZ) in inflammatory and apoptotic responses induced by middle cerebral artery occlusion (MCAO) in rats. Methods: MCAO-induced focal cerebral ischemia in rats was used in this study. The hypoxia-inducible factor-1α (HIF-1α), activation of caspase-3, and TNF-α mRNA transcription in ischemic regions were detected by immunoblotting and RT-PCR, respectively. Anti-oxidative activity was investigated using a thiobarbituric acid-reactive sub-stance (TBARS) test in rat brain homogenate preparations. Results: We showed the statistical results of the infarct areas of solvent- and TMPZ (20 mg/kg)-treated groups at various distances from the frontal pole in MCAO-induced focal cerebral ischemia in rats. Treatment with TMPZ (20 mg/kg) markedly reduced the infarct area in all regions, especially in the third to fifth sections. MCAO-induced focal cerebral ischemia was associated with increases in HIF-1α and the activation of caspase-3, as well as TNF-α transcription in ischemic regions. These expressions were markedly inhibited by treatment with TMPZ (20 mg/kg). However, TMPZ (0.5–5 mmol/L) did not significantly inhibit TBARS reaction in rat brain homogenates. Conclusion: The neuroprotective effect of TMPZ may be mediated at least by a portion of the inhibition of HIF-1α and TNF-α activations, followed by the inhibi-tion of apoptosis formainhibi-tion (active caspase-3), resulting in a reducinhibi-tion in the infarct volume in ischemia-reperfusion brain injury. Thus, TMPZ treatment may represent an ideal approach to lowering the risk of or improving function in is-chemia-reperfusion brain injury-related disorders.
Key words
tetramethylpyrazine; middle cerebral artery occlusion; HIF-1α; caspase-3; TNF-α; thio-barbituric acid-reactive substance
1 Pr oject suppor ted by grants from t he Topnotch Stroke Research Center, Taipei Medical University; the Shin Kong Wu Ho-Su Memorial Hospital (SKH-TMU-94-07); and the Committee on Chinese Medicine and Pharmacy (CCMP 94-RD-031).
5 Correspondence to Prof Joen-rong SHEU. Phn/Fax 886-2-2739-0450. E-mail sheujr@tmu.edu.tw Received 2006-07-15 Accepted 2006-10-15 doi: 10.1111/j.1745-7254.2007.00514.x Introduction
Tetramethylpyrazine (TMPZ) is an active alkaloid puri-fied from a commonly used Chinese herb named “Chung Chong” (Ligusticum wallichii Franchat), which has been used for at least 2000 years by traditional Chinese physi-cians to stimulate blood circulation, relieve pain (Figure 1)[1],
and treat a variety of vascular diseases, notably ischemic stroke and pulmonary hypertension secondary to chronic
obstructive pulmonary diseases[1,2]. It has been reported
that TMPZ improves changes in the microcirculation of
patients with acute cerebral thrombosis[3]. Ho et al[4] showed
that TMPZ increased the survival rate of Mongolian gerbils with experimentally-induced stroke. Furthermore, TMPZ has exhibited effective antiplatelet activity in both in vitro and
in vivo studies[5,6].
Ischemic brain injury often causes irreversible brain damage. The cascade of events leading to neuronal injury and death in ischemia includes excitotoxicity, inflammation,
edema formation, apoptosis, and necrosis[7]. As described
previously, we demonstrated that TMPZ possesses marked
on cerebral ischemic damage in middle cerebral artery occlu-sion (MCAO)-reperfuocclu-sion rats is assumed to be mediated, at least partially, by the inhibition of platelet aggregation, neu-trophil activation, and inflammatory responses such as in-ducible nitric oxide synthase (iNOS) expression, and reac-tive oxygen species formation in cerebral ischemic areas. Furthermore, several reports have also demonstrated that TMPZ possesses neuroprotection against ischemic brain injury through the reduction of neuronal apoptosis (eg, Bcl-xL)[9,10].
Recently, it was demonstrated that hypoxia-inducible factor-1α (HIF-1α) expression increases in the rat brain dur-ing cerebral ischemia induced by different models of arterial
occlusion or cardiac arrest[11]. The increased HIF-1α
pro-tein level observed after ischemia-reperfusion was
presum-ably induced by the loss of the oxygen supply[12], resulting
in a greater extent of binding activity to the iNOS gene which reaches a consequent peak of iNOS protein expression. HIF-1α may bind to the iNOS promoter gene under hypoxic condi-tions. Such binding is associated with an increase in iNOS expression[13].
In our previous study[8], we partially resolved the
mecha-nisms of TMPZ in MCAO-induced transient focal cerebral ischemia in rats; however, the detailed mechanisms underly-ing the inhibitory effect of TMPZ in inflammatory and apoptotic responses induced by MCAO have still not yet been completely resolved. We therefore further examined the effect of TMPZ on MCAO-induced cerebral ischemia, and utilized the findings to further characterize the neuroprotective effect of TMPZ.
Materials and methods
Animals Male Wistar rats (250–300 g) were used in this study. All animal experiments and care were performed ac-cording to the Guide for the Care and Use of Laboratory Animals (National Academy Press, Washington DC, 1996). Before undergoing the experimental procedures, all animals were clinically normal and free of apparent infection or
inflammation, and showed no neurological deficits. The
ani-mals were anesthetized with a mixture of 75% air and 25% O2
gases containing 3% isoflurane. The rectal temperature was maintained at 37±0.5 °C.
MCA occlusion assay The right MCA was occluded as described previously[8,14]. Briefly, the right common carotid
artery was exposed and a 4-0 monofilament nylon thread (25 mm) coated with silicon was then inserted from the external into the internal carotid artery until the tip occluded the ori-gin of the MCA. After closure of the operative sites, the animals were allowed to awake from the anesthesia. During another brief period of anesthesia, the filament was gently removed after 1 h of MCAO.
Forelimb akinesia test An observer blinded to the iden-tity of the groups assessed the neurological deficits at 1 and 24 h after reperfusion (before sacrifice) using the forelimb akinesia (also called the postural tail-hang) test, while the spontaneous rotational test was used as a criterion for
evalu-ating the ischemic insult[15]. Animals not showing
behav-ioral deficits at the above time points after reperfusion were excluded from the study. However, reperfusion was also ensured by an improvement in the ipsilateral local blood flow of at least 60% of the baseline following an initial sharp de-crease to about 30% of the baseline caused by MCAO as determined using a continuous laser Doppler flowmeter (Oxford Optronix, Oxford, UK) with a standard needle probe (pp-051). The rats were sacrificed by decapitation after 24 h of reperfusion. Their brains were cut into 2 mm coronal slices. Each stained brain (2% 2,3,5-triphenyltetrazolium, TTC) slice was drawn using a computerized image analyzer (Image-Pro Plus, Media Cybernetics, Silver Spring, MD, USA). The cal-culated infarct areas were then compiled to obtain the infarct
volumes (mm3) for each brain. Infarct volumes were expressed
as a percentage of the contralateral hemisphere volume us-ing the formula [the area of the intact contralateral (left) hemi-sphere – the area of the intact region of the ipsilateral (right) hemisphere] to compensate for edema formation in the ipsi-lateral hemisphere[14]. All of the animals were divided into 3
groups: (i) a sham-operated group; (ii) a solvent solution-treated group (cremophor: ethanol: normal saline, 1:1:4); and (iii) a single dose (20 mg/kg, ip) of TMPZ (Aldrich, Milwaukee, MI, USA)-treated group. In the group treated with the sol-vent or TMPZ, the rats were given isovolumetric solsol-vent or TMPZ (20 mg/kg) 20 min before MCAO.
Preparation of brain tissue sample The MCAO-insulted and sham-operated rats were anesthetized with chloral hy-drate (400 mg/kg, ip), then the apex of the heart was pen-etrated with a perfusion cannula inserted through the left ventricle into the ascending aorta. Perfusion with ice-cold Figure 1. Chemical structure of tetramethylpyrazine.
phosphate-buffered saline was performed, and an incision was made in the right atrium for venous drainage. Fresh brains were removed and sectioned coronally into 4 sequen-tial parts from the frontal lobe to the occipital lobe. The third part (3–9 mm from the frontal lobe) of each hemisphere was separately collected, snap-frozen in liquid nitrogen, and stored at -70 °C.
Western blot assay Expression of HIF-1α and activated
caspase-3 in the brain were analyzed by Western blotting as
described by Rodrigo et al [16] with some modifications. The
frozen tissues were weighed and placed in ice-hold homoge-nate buffer at a ratio of 1 g tissue to 1 mL buffer. Each brain tissue sample was homogenized using a polytron homo-genizer, then sonicated for 10 s 3 times at 4 °C. The soni-cates were subjected to centrifugation (at 10 000×g).The su-pernatant (50 µg protein) was subjected to SDS-PAGE and electrophoretically transferred to PVDF membranes (0.45 µm; Hybond-P, Amersham, Buckinghamshire, HP, UK). After incubation in blocking buffer (10 mmol/L Tris-base, 100 mmol/L NaCl, 0.1% Tween 20, and 5% dry-skim milk, pH 7.5) and after washing 3 times with TBST buffer (10 mmol/L Tris-base, 100 mmol/L NaCl, and 0.1% Tween 20, pH 7.5), the blots were hybridized with an HIF-1α polyclonal body (1:1000, R&D, Minneapolis, CA, USA), and an anti-active caspase-3 pAb (1:250, BioVision, Mountain View, CA, USA) or an anti-actin mAb (1:7000, Sigma, St Louis, MO, USA) in TBST buffer overnight. Blots were subsequently washed 4 times with TBST and incubated with secondary horseradish peroxidase-conjugated goat anti-mouse mAb (Amersham) for 1 h. The blots were then washed and the immunoreactive protein was detected using film exposure with enhanced chemiluminescence detection reagents (Amersham).
Isolation of total RNA and RT-PCR Total RNA was iso-lated from the ipsilateral cortex by a commercially available kit (TRIzol, Gibco, Grand Island, NY, USA) according to the manufacturer’s instructions. For each RT-PCR reaction, 0.5 mg of the RNA sample and 0.2 µmol/L of primers were reverse-transcribed and amplified in a 50 µL of reaction mix-ture of commercially available reagents (SUPERSCRIPT One-Step RT-PCR with PLATINUM Taq Kit, Invitrogen, Carlsbad, CA, USA) containing a 1× reaction mixture and 0.2 µmol/L of an RT/Taq mixture in 1 cycle of 30 min at 50 °C for reverse transcription and 1 cycle at 95 °C for 3 min; followed by 40 cycles at 95, 62, and 72 °C for 30, 40, and 40 s, respectively; with a single extension step at 72 °C for 5 min, followed by 4 °C for amplification in a thermal cycler (GeneAmp PCR system 2400, Perkin-Elmer, Wellesley, MA, USA). For visu-alization and quantification by densitometry of each RT-PCR
reaction, a 10 µL aliquot was subjected to electrophoresis on a 1.5% agarose gel using a mini horizontal submarine unit (HE 33) containing 0.5 mg/mL ethidium bromide to allow UV-induced fluorescence (TCP-20.M, Vilber Lourmat, Marne-la-Vallee Cedex, France).
Anti-oxidative activity in rat brain homogenate prepara-tions Rat brain homogenates were prepared from the brains of freshly killed Wistar rats, and the peroxidation in the pres-ence of iron ions was measured by the thiobarbituric acid
method, as described by Braughler et al[17], with some
modi-fications. In brief, whole brain tissue, excluding the cere-bellum, was washed and homogenized in 10 volumes of ice-cold Krebs buffer using a homogenizer (Glas-col, Millville, NJ, USA). The homogenate was centrifuged at low speed (1000×g) for 10 min, and the resulting supernatant (adjusted to 2 g/L) was used immediately in the lipid peroxidation assay. The reaction mixture with TMPZ or vehicle solution (0.5% DMSO) was incubated for 10 min, then stimulated by the addition of a ferrous ion (200 mmol/L, freshly prepared), and maintained at 37 °C for 30 min. The reaction was terminated by adding 10 µL of ice-cold trichloroacetic acid solution [4% (w/v) in 0.3 mol/L HCl] and 200 µL of thiobarbituric acid-reactive substance reagent [TBARS, 0.5% (w/v) thiobar-bituric acid in 50% (v/v) acetic acid]. After boiling for 15 min, the samples were cooled and extracted with n-1-butanol. The extent of lipid peroxidation was estimated by TBARS and was read at 532 nm in a spectrophotometer (Model U3200, Hitachi, Tokyo, Japan). Tetramethoxypropane was used as a standard, and the results were expressed as nanomoles of malondialdehyde equivalents per milligram protein of the supernatant of rat brain homogenates. The protein contents of the brain homogenates and other preparations were de-termined with the Bio-Rad method.
Statistical analysis The experimental results are ex-pressed as the mean±SEM and are accompanied by the num-ber of observations. Student’s unpaired t-test was used to determine significant differences in the study of MCAO-induced cerebral ischemia. The other experiments were as-sessed by ANOVA. If this analysis indicated significant differences compared with the group means, then each group was compared using the Newman-Keuls method. A P value of less than 0.05 was considered statistically significant.
Results
Treatment with TMPZ can reduce the infarct area in rat brain The animals of all the groups in this study showed similar physiological values for rectal temperature, mean ar-terial blood pressure, plasma glucose, and hematocrit (%)
before, during, and after MCAO (data not shown). Neither abnormal behavior, depression of respiratory function, nor hypothermia was observed in the solvent- or TMPZ-treated groups. Cerebral infarction was examined in 2 mm-thick slices of the cerebrum of MCAO-reperfused rats through TTC
staining. In our previous report[8], administration of TMPZ
at 10 and 20 mg/kg produced concentration-dependent re-ductions in infarct volumes compared with the solvent group. In this study, we further showed the statistical results of the infarct areas of the solvent- and TMPZ (20 mg/kg)-treated groups at various distances from the frontal pole (Figure 2). Treatment with TMPZ (20 mg/kg) reduced the infarct area in all regions, especially in the third to fifth sections (Figure 2). In the solvent-treated rats, approximately 58.6%±4.1% of the entire area was infarcted in the third section, while TMPZ (20 mg/kg) ingestion reduced the area to 37.6%±4.7% in the third section (Figure 2).
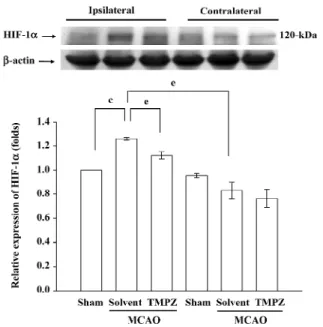
TMPZ treatment can reduce expression of HIF-1α and
activated caspase-3 in the ischemic rat brain We further examined the expression of HIF-1α when treated with TMPZ in the ischemic rat brain. As shown in Figures 3 and 4, the expression of HIF-1α and activated caspase-3 were upregu-lated after MCAO-reperfusion injury. TMPZ (20 mg/kg)
treat-ment significantly (P<0.05) suppressed the level of HIF-1α
and activated caspase-3 in the ipsilateral hemisphere (Figures 3 and 4).
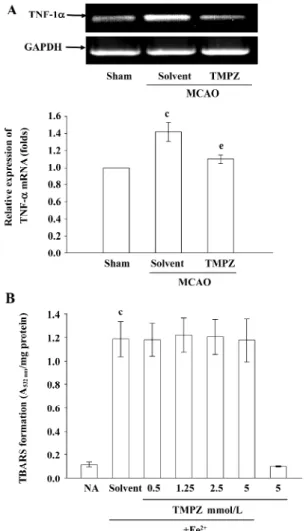
TMPZ treatment can reduce the transcription of TNF-α
gene in the ischemic rat brain Transient MCAO also resulted in a significant and more sustained increase in the
expression of TNF-α mRNA in the injured hemisphere com-pared with the levels obtained in the corresponding areas of the sham-operated group (Figure 5A). TMPZ (20 mg/kg) treatment significantly reduced this reaction (Figure 5A).
In addition, TMPZ was further tested for its ability to inhibit non-enzymatic lipid peroxidation in rat brain homogenates stimulated by a ferrous ion. At 0.5–5 mmol/L, TMPZ did not significantly inhibit the ferrous-induced lipid peroxidation in normal (Figure 5B) or MCAO-insulted rat brain homogenates (n=3, data not shown). In addition, TMPZ (0.5–5 mmol/L) also did not inhibit the ferrous-induced lipid peroxidation in rat brain homogenates. TMPZ did not inter-fere with the thiobarbituric acid test, since the color forma-tion was unchanged if it was added after the incubaforma-tion with
thiobarbituric acid reagents. However, α-tocopherol (200
µmol/L) inhibited ion-dependent lipid peroxidation in this reaction (data not shown).
Figure 2. Protective effect of TMPZ in MCAO-induced rats with
cerebral ischemia. Cerebral infarction in MCAO-reperfusion rats is from repr esenta t ive animal s tha t received solvent (sol vent ; cremophor: ethanol: normal saline 1:1:4, n=14) or TMPZ (20 mg/ kg, n=8) intraperitoneally. Infarct volumes are calculated as de-scribed in Materials and methods. Each point (○, solvent-treated group, n=14; △, TMPZ 20 mg/kg-treated group, n=8) and vertical bar represent mean±SEM. bP<0.05, cP<0.01 as compared with the solvent-treated group.
Figure 3. Effect of TMPZ on the expression of HIF-1α in MCAO-reperfusion injury in rats. Fresh brains from sham-operated (sham) or solvent-treated (solvent), and TMPZ (20 mg/kg)-treated rats were removed and sectioned coronally into 4 sequential parts from the frontal lobe to the occipital lobe. The third parts of the 4 sequential parts of the ipsilateral (right) and contralateral (left) hemispheres were separately collected, homogenized, and centrifuged. The super-natant (50 µg protein) was then subjected to SDS-PAGE, and trans-ferred onto membranes for analysis of HIF-1α expression. The re-sults are representative examples of 4 similar experiments. Data are presented as the mean±SEM. cP<0.01 compared with the sham-oper-ated group; eP<0.05 as compared with the solvent-treated groups. Equal loading in each lane is demonstrated by similar intensities of β-actin.
Discussion
TMPZ can permeate the blood–brain barrier and can be enriched in the brain, especially the brainstem. The present study demonstrates that MCAO-reperfusion injury induces increases in HIF-1α and active caspase-3 protein expressions, and TNF-α mRNA expression, which may represent the re-sponse of neurons suffering from the ischemic insult. In fact, the increase in HIF-1α expression after an ischemic in-sult may exert opposite effects on neuronal fates[11, 12].
HIF-1α activation of target genes related to vascularization,
glu-cose transport, and glycolytic metabolism may be regarded as an adaptive response to ischemic conditions that may promote neuronal survival within ischemic areas. However, in other instances, the increased activation of HIF-1α may act as a noxious signal for neuronal survival. HIF-1α, which combines with p53, may promote apoptotic cell death in is-chemic areas[11].
In this study, we showed that the elevation of active caspase-3 expression occurred in the same time frame as HIF-1α expression after ischemic injury, and these expres-sions could be significantly suppressed by pretreatment with TMPZ (20 mg/kg).
It has been reported that several apoptosis-related genes, including caspase-9 and -3, are all strongly expressed after
ischemic injury[18]. In addition, hypoxia may cause HIF-1α
to bind to p53 in order to stabilize it, and also activates the expression of various genes, including bax (a pro-apoptotic
member of Bcl-2 family proteins)[18]. Bax is translocated to
the mitochondria where it releases cytochrome c into the cytosol to interact with Apaf-1 to activate caspase-9, which
in turn activates downstream caspases, such as caspase-3[19].
TNF-α is one of the key immunomodulatory and pro-Figure 4. Effect of TMPZ on the expression of active caspase-3 in
cerebral homogenates after MCAO-reperfusion injury in rats. The results are representative examples of 4 similar experiments. Data are presented as mean±SEM. cP<0.01 compared with the sham-oper-ated group; eP<0.05 and fP<0.01 compared with the solvent-treated groups. Equal loading in each lane is demonstrated by similar inten-sities of β-actin.
Figure 5. Effects of TMPZ on the (A) expression of TNF-α mRNA in MCAO-reperfusion in rats, and (B) the anti-oxidation in TBARS in rat brain homogenate. (A) Results are representative of 4 similar experiments. GADPH levels were used to normalize the amount of the cDNA template used in each PCR reaction. Data are presented as mean±SEM. (n=4). cP<0.01 compared with the sham-operated group; eP<0.05 compared with the solvent-treated groups. (B) Brain homo-genates were pre-incubated with solvent (0.5% DMSO) or various concentrations of TMPZ (0.5, 1.25, 2.5, and 5 mmol/L) for 10 min followed by the addition of 200 µmol/L Fe2+. Results are presented as the absorbance at 532 nm/mg protein in brain homogenates. Data are presented as mean±SEM. (n=3). cP<0.01 compared with the normal saline group (NA, resting group).
inflammatory cytokines upregulated during brain ischemia[20].
Administration of TNF-α during an ischemic brain insult has been shown to augment the injury, as evidenced by increased
tissue damage and neurological deficits[20]. In addition to
inflammation, TNF-α has also been shown to be involved in
apoptosis[21]. In our study, we also found TMPZ can
downregulate the transcription of TNF-α during brain ischemia. Therefore, TMPZ inhibition of active caspase-3 expression may occur, at least partially, through the inhibi-tion of TNF-α expression in ischemic brain injury.
The phospholipid bilayers of cellular and subcellular membranes are undoubtedly major targets for free radicals. The compound that inhibits membrane phospholipid peroxidation seems to exert a pharmacological effect in the prevention of radical-induced oxidative pathological events. Among cell-free systems, brain homogenates are usually chosen to evaluate an anti-oxidant’s effects on lipid
peroxidation[17]. Rat brain homogenates exposed to a
fer-rous ion exhibit lipid peroxidation in air by a mechanism whose induction step may primarily involve site-bound, iron-medi-ated decomposition of lipid hydroperoxides to yield alkoxy or peroxyl radicals, leading to the chain reaction of lipid
peroxidation[22]. In this system, TMPZ did not effectively
inhibit lipid peroxidation, indicating that the inhibition of lipid peroxidation might not be the neuroprotective mecha-nism for TMPZ in ischemic brain injury.
In conclusion, we found that the neuroprotective effect of TMPZ on cerebral ischemic damage in MCAO-reperfusion rats is probably mediated by the inhibition of HIF-1α and TNF-α activation, followed by the inhibition of apoptosis (active caspase-3). The rationale for the use of TMPZ is based on the fact that the multiple deleterious process in different cell types of organelles are initiated during ischemia-reperfusion injury which ultimately synergistically moves toward irreversible injury. Therefore, treatment using TMPZ is not limited to 1 factor, but involves many mechanisms, most of which may be interrelated. We speculate that the correction of these molecules and morphological changes may lead to neurobehavioral improvement in patients; thus, treatment using TMPZ may represent an ideal approach for improving function after ischemia-reperfusion brain injury.
References
1 Beijing Institute of Pharmaceutical Industry. Studies of active components of Ligusticum wallichii Franch. III. A report of the treatment of tetramethylpyrazine in 218 patients with ischemia cerebrovascular diseases. Chin Med J 1977; 8: 467–9. 2 Chen KJ, Chen K. Ischemic stroke treated with Ligusticum
chuanxiong. Chin Med J 1992; 105: 870–3.
3 Wang WF. Changes of nail fold microcirculation in 28 cases of acute cerebral thrombosis treated with Ligustrazine. Chin J Neurol Psychiatry 1984; 17: 121–4. Chinese.
4 Ho WKK, Web HL, Lee CM. Tetramethylpyrazine for treat-ment of experitreat-mentally induced stroke in Mongolian gerbils. Stroke 1989; 20: 96–9.
5 Sheu JR, Kan YC, Hung WC, Ko WC, Yen MH. Mechanisms involved in the antiplatelet activity of tetramethylpyrazine in human platelets. Thromb Res 1997; 88: 259–70.
6 Sheu JR, Hsiao G, Lee YM, Yen MH. Antithrombotic effects of tetramethylpyrazine in in vivo experiments. Int J Hematol 2001; 73: 393–8.
7 Kuroda S, Siesjo BK. Reperfusion damage following focal ischemia: pathophysiology and therapeutic windows. Clin Neurosci 1997; 4: 199–212.
8 Hsiao G, Chen YC, Lin JH, Lin KH, Chou DS, Lin CH, et al. Inhibitory mechanisms of tetramethylpyrazine in middle cere-bral artery occlusion (MCAO) -induced focal cerecere-bral ischemia in rats. Planta Med 2006; 72: 411–7.
9 Liao SL, Kao TK, Chen WY, Lin YS, Chen SY, Raung SL, et al. Tetramethylpyrazine reduces ischemic brain injury in rats. Neurosci Lett 2004; 372: 40–5.
1 0 Kao TK, Ou YC, Kuo JS, Chen WY, Liao SL, Wu CW, et al. Neuroprotection by tetramethylpyrazine against ischemic brain injury in rats. Neurochem Int 2006; 48: 166–76.
1 1 Matrone C, Pignataro G, Molinaro P, Irace C, Scorziello A, Di Renzo GF. HIF-1α reveals a binding activity to the promoter of iNOS gene after permanent middle cerebral artery occlusion. J Neurochem 2004; 90: 368–78.
1 2 Wiener CM, Booth G, Semenza GL. In vivo expression of mRNAs encoding hypoxia-inducible factor 1. Biochem Biophys Res Commun 1996; 225: 485–8.
1 3 Melillo G, Musso T, Sica A, Taylor LS, Cox GW, Varesio L. A hypoxia-responsive element mediates a novel pathway of acti-vation of the inducible nitric oxide synthase promoter. J Exp Med 1995; 182: 1683–93.
1 4 Hsiao G, Lin KH, Chang Y, Chen TL, Tzu NH, Chou DS, et al. Protective mechanisms of inosine in platelet activation and ce-rebral ischemic damage. Arterioscler Thromb Vasc Biol 2005; 25: 1998–2004.
1 5 Nishino H, Czurko A, Onizuka K, Fukuda A, Hida H, Ungsuparkorn C, et al. Neuronal damage following transient cerebral ischemia and its restoration by neural transplant. Neurobiology 1994; 2: 223–34.
1 6 Rodrigo J, Alonso D, Fernandez AP, Serrano J, Richart A, Lopez JC, et al. Neuronal and inducible nitric oxide synthase expression and protein nitration in rat cerebellum after oxygen and glucose deprivation. Brain Res 2001; 909: 20–45.
1 7 Braughler JM, Burton PS, Chase RL, Pregenzer JF, Jacobsen EJ, Vandoornik FJ, et al. Novel membrane localized iron chelators as inhibitors of iron-dependent lipid peroxidation. Biochem Pharmacol 1988; 37: 3853–60.
1 8 Gibson ME, Han BH, Choi J, Knudson CM, Korsmeyer SJ, Parsadanian M, et al. BAX contributes to apoptotic-like death following neonatal hypoxia-ischemia: evidence for distinct apoptosis pathways. Mol Med 2001; 7: 644–55.
1 9 Suzuki H, Tomida A, Tsuruo T. Dephosphorylated hypoxia-inducible factor 1 alpha as a mediator of p53-dependent apoptosis
during hypoxia. Oncogene 2001; 20: 5779–88.
2 0 Barone FC, Arvin B, White RF, Miller A, Webb CL, Willette RN, et al. Tumour necrosis factor α. A mediator of focal ischemic brain injury. Stroke 1997; 28: 1233–44.
2 1 Gupta S. Molecular steps of tumor necrosis factor
receptor-mediated apoptosis. Curr Mol Med 2001; 1: 317–24.
2 2 Braughler JM, Chase RL, Pregenzer JF. Oxidation of ferrous iron during peroxidation of lipid substrates. Biochim Biophys Acta 1987; 921: 457–64.
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
The 7
thChinese Symposium on Calcium Signalling
(The 7
thCSCS Meeting)
October/November 2008, Wuhan, China
Info: Prof Zong-jie CUI, PhD Institute of Cell Biology Beijing Normal University Beijing 100875, China Phn/Fax 86-10-5880-9162 E-mail zjcui@bnu.edu.cn