Regeneration patterns of yellow cypress on down logs in mixed
coniferous-broadleaf forest of Yuanyang Lake Nature Preserve,
Taiwan
Chi-Cheng
Liao
1, Chang-Hung Chou
2, and Jiunn-Tzong Wu
1,3,*
1Department of Botany, National Taiwan University, Taipei 106, Taiwan
2Graduate Institute of Tropical Agriculture, National Pingtung University of Science and Technology, Pintung 912,
Taiwan
3Institute of Botany, Academia Sinica, Taipei 115, Taiwan
(Received August 26, 2002; Accepted May 6, 2003)
Abstract. The importance of down logs in forest gaps for forest development, tree regeneration, and the co-existence
of yellow cypress (Chamaecyparis obtusa Sieb. & Zucc. var. formosana (Hayata) Rehder) and broadleaf trees was studied in the temperate forest of Yuanyang Lake Nature Reserve (YYL), Taiwan. On nine down logs in a plot of 0.25-ha, seedlings with diameter at breast height larger than 1 cm were recorded. It was found that the species occur-ring on the sampled down logs were the same as those in the studied area. Few broadleaf trees could reach the height of 10 m necessary to change the current canopy layer, which mainly consisted of yellow cypress. Regeneration of trees on down logs was sufficient to maintain the forest community, including its floristic composition, canopy structure, and species diversity. Tree seedlings of different species also varied in their distribution patterns on down logs. Yellow cypress seeds germinated on new down logs, but not on root swellings. The seedlings grew to fill gaps, while density was reduced from young to old down logs, possibly due to intra-species competition. Epiphytic bryo-phytes on new down logs seemed to provide a suitable habitat for the germination of yellow cypress seeds. In contrast, broadleaf trees grew mainly on the base and root swellings of new down logs and gradually recruited their seedlings onto other parts of older down logs. Availability of nutrient resources seemed to be the important determi-nant for the competition between seedlings of cypress and broadleaf plants. The niche partition hypothesis was employed to explain the regeneration of cypress plants in this nature preserve.
Keywords: Chamaecyparis obtusa var. formosana; Down log; Niche partitioning; Regeneration; Yellow cypress;
Yuanyang Lake Nature Preserve.
Introduction
Gap dynamics, including tree death, seedling regeneration, and adult replacement, have been studied in temperate (Runkle, 1981; Runkle, 1982; Veblen, 1986; Yamamoto, 1988; Nakashizuka, 1989; Lertzman, 1992; Gray and Spies, 1996; Runkle, 2000) and tropical forests (Brokaw, 1985; Arriaga, 1988; Lawton and Putz, 1988; Uhl et al., 1988; Dalling et al., 1998), but have received little attention in the temperate forests of Taiwan. Previous studies have considered succession trends of the forests (Chang, 1961; Liu, 1971), but not gap dynamics or regen-eration of trees.
Chamaecyparis obtusa Sieb. & Zucc. var. formosana
(Hayata) Rehder (yellow cypress) is one of the dominant tree species in the temperate forests in Taiwan (Su, 1984; Jen, 1995). Yellow cypress was reported to establish seed-lings on down logs and mineral soils in some Taiwan for-*Corresponding author. Tel: 886-2-2789-9590 ext 420; Fax:
886-2-2782-7954; E-mail: jtwu@gate.sinica.edu.tw
ests (Liu et al., 1961), but no further studies were done on this phenomenon. Most studies concerning recruitment of yellow cypress has been done in plantations, and they concluded that thinning broadleaf trees could promote es-tablishment of yellow cypress seedlings by reducing in-ter-specific competition (Hung, 1984; Chiu et al., 1995; Lo-Cho et al., 1999). However, this conclusion was based on the premise that the resource requirements of broad-leaf and yellow cypress trees overlap and that yellow cy-press is a weak competitor. There is some uncertainty regarding these viewpoints.
Yellow cypress and broadleaf trees are known to co-exist in a forest at Yuanyang Lake Natural Preserve (YYL), northern Taiwan (Chou et al., 2000). This virgin mixed co-niferous-broadleaf forest is free from human disturbance, and it is of interest to consider the mechanisms that might allow the trees to coexist in this natural system. Based on past research, both broadleaf trees and yellow cypress seedlings occur in abundance on down logs (Liu et al., 1961). The role of these logs may therefore be important to the regeneration and co-existence of trees in YYL.
Gap dynamics are thought to be important in promot-ing species diversity and forest development (Lertzman, 1992; Poulson and Platt, 1996; Gray and Spies, 1996; Dalling et al., 1998). Tree species diversity in forests could be main-tained by providing a different regeneration niche for the species (Grubb, 1977). Forest development is determined by the composition of seedlings and the overlying forest community. If seedling composition is the same as the forest composition, the current community might be main-tained (Lertzman, 1992; Poulson and Platt, 1996). Otherwise, floristic composition might change over time (Lertzman, 1992; Poulson and Platt, 1996). If down logs are important for species co-existence and dynamics, they might be important for determining floristic composition and forest development.
The niche-partitioning hypothesis has been proposed to explain the correlations among the resource gradient of gaps, seedling establishment, and species co-existence (Ricklefs, 1977; Denslow, 1980). The resource gradient was presumed to correspond with gap size classes and within-gap positions, and tree seedlings were thought to respond to the gradient by establishing at different locations (Runkle, 1981; Runkle, 1982; Brokaw, 1985; Gray and Spies, 1996; Dalling et al., 1998). Instead of a resource gradient based on gaps, substrate variation was reported to affect seedling establishment in some coniferous forests (Putz, 1983; Harmon and Franklin, 1989). Regeneration of some conifer seedlings was favored by down logs (Spies et al., 1988; Nakashizuka, 1989; Gray and Spies, 1997). In YYL, co-existence of trees has been observed on down logs, but there are no reports on tree species establishment or dynamic patterns of the species on these substrata.
In this article, the objective is to quantify the relation-ship between tree regeneration on down logs and forest development in YYL. In particular, this study aims to elu-cidate the role of down logs in tree regeneration, to deter-mine whether broadleaf trees and yellow cypress had regular recruitment patterns, and whether niche partition-ing theory could explain the coexistence of these two groups of plants.
Materials and Methods
Site DescriptionThe study was done in the YYL, located on Chi-lan Mountain (24°35' N, 121°24' E), part of the Sheashan Moun-tain Range in the northeastern part of Taiwan, at an eleva-tion of 1,650-2,432 m above sea level (a.s.l.), from 1998 to 2001. The YYL was declared a protected site to preserve the integrity of a mountain lake, virgin forest, and some aquatic species (Hwang et al., 1996; Chou et al., 2000). The lake has an area of 3.6 ha and is located at the east side of the reserve. A mixed coniferous-broadleaf forest extends from the lakeside (1,760 m a.s.l.) to the mountain ridge (2,432 m a.s.l.). The YYL has been virtually undisturbed by human activities.
Within the forest, yellow cypress is the dominant species, and it is the major species occupying the canopy
layer (Liu and Hsu, 1973; Chou et al., 2000). Tsuga
chinensis is another rare conifer, found on the summit of
YYL. Most of the broadleaf woody plants are shrubs, and only a few reach the canopy layer. The open canopy layer is formed mostly by yellow cypress, and small gaps in it are usually found. The canopy and shrub layers of the for-ests at lakeside and summit have similar components, but the dominant components of the undergrowth are different. Plagiogyria glauca var. philippinensis domi-nates the lakeside forest (Chou et al., 2000). In contrast, a dense bamboo, Yushania niitakayamensis, occupying the undergrowth at the summit, dominates most herb species on the forest floor.
The climate of YYL is characterized by abundant precipitation. Heavy cloud cover was previously reported to be associated with the presence of yellow cypress (Su, 1984). Annual precipitation, mostly contributed by the monsoon, is about 3,000 mm and occasionally is higher than 4,000 mm (Hwang et al., 1996; Chang et al., 2001). Heavy cloud and fog are so frequent that solar radiation is remarkably reduced. Heavy moisture results in abun-dant growth of mosses on the forest floor, tree trunks, and fallen logs in the YYL (Hwang et al., 1996; Chang et al., 2002). The average annual air temperature is 13°C; the highest monthly mean is 18°C in June, and the lowest perature is 5°C in January (Hwang et al., 1996). The tem-perature is rarely lower than 0°C, and snowfall is rare in winter.
Field Study and Data Analysis
Forest composition. Current forest composition,
stud-ied from 1998 to 2001, was obtained from a census of trees in a 0.25-ha plot located 200 m northwest of the lake. Ac-cording to Chou et al. (2000), we recorded the woody stems with a diameter at 130 cm above ground (diameter at breast height, DBH) larger than 1 cm in the 0.25-ha plot. The rela-tive density of a particular species was measured as the proportion of total individuals.
Selecting down logs and census of seedlings. The
boundary of a gap was difficult to define because the canopy layer was not continuous in the YYL forest. The shrub layer and undergrowth surrounding down logs was not obviously different from the forest floor under canopy shade. Data from the plot census revealed that abundant yellow cypress and broadleaf tree seedlings occurred on down logs. These down logs were sampled to investigate regeneration characteristics and processes in YYL forest.
Discontinuous disturbances resulted in different stages of seedling recruitment among gaps of varying ages. In this study, only down logs not covered by other logs, limbs, or living tree trunks were sampled. Although down logs were quite common in YYL, only two down logs met this criterion within the original 0.25-ha plot (namely logs 8 and 9). Therefore, following the inventory, an additional seven down logs were surveyed in a 7-ha lakeside area. The total of nine down logs was sampled for the regen-eration studies in YYL (Table 1).
Segment division of down logs. Each down log was
divided into root, trunk, and branch segments in order to determine whether resource gradient and distribution are different among colonizing tree species. The root segment ran from the root swelling to the trunk base of 150 cm long. The trunk segment ran from the trunk base of 150 cm long to the log’s first branch. The branch segment ran from the first branch to the log’s top. Because these down logs had variable lengths and their diameter near the tips was narrow, five meters of branch segments were studied to reduce the sample error.
All seedlings on down logs were identified. The height, diameter at stem base (basal diameter, BD), locating segment, and distances along segments of each seedling were recorded. For distance, a value of zero was assigned if seedlings grew at the base point of the trunk. If seed-lings occurred between the trunk base and the log tip, the distance to the trunk base was positive. A negative value was obtained when seedlings grew on root swelling of down logs. Seedling composition on down logs was mea-sured as the relative density of all seedling species. A comparison of the forest composition from the 0.25-ha plot, and seedling composition on the down logs, was made to evaluate the succession trend.
Grouping down logs. In order to analyze the
recruit-ment patterns of seedlings, down logs were classified into three groups. The classification was based on the aver-age diameter of all yellow cypress seedlings of each type (Table 1). In years following the fall of a yellow cypress, yellow cypress seedlings were found to recruit on it. Thus, the date of a log’s fall could be estimated from the age of the oldest yellow cypress seedling growing on it. Group I was composed of logs 2, 3 and 4 with an average diam-eter of less than 1 mm, while group II, of logs 7, 8 and 9, had average diameters of 5 to 15 mm, and group III, of logs 1, 5 and 6, had average diameters greater than 15 mm.
Bryophytes and organic matters sampled on down logs.
Micro-habits on the down logs were characterized by cov-erage of epiphytic bryophytes and organic matter. A cir-cular sample (5 cm in diameter) was taken every 50 cm along root segments and at every 100 cm along trunk and branch segments. Living parts of bryophytes were
sepa-rated from dead parts and litter fall. Sampled areas were also recorded to estimate their weights per square centimeter. Bryophyte samples and dead organic matters were dried at 65 °C for two days and weighed.
Data analysis. Average values of basal diameter of all
seedlings and dry weights of bryophytes and organic mat-ters on down logs were analyzed with respect to differ-ences between the three groups identified above. One-way analysis of variance (ANOVA) was performed, followed by Tukey’s test for pairwise comparisons of treatment means.
Results
Comparisons of Floristic Composition
Yellow cypress was the dominant tree species in the YYL forest community (Table 2). A total of 182 individuals with diameters larger than 1cm were recorded in the 0.25-ha plot studied. They constituted the canopy and 0.25-had low relative density and high basal area in the forest, present-ing an inverse J-shaped pattern in their size. Furthermore, 102 yellow cypress seedlings were found on down logs 8 and 9 inside the plot. Therefore, the yellow cypress pop-ulations included plants at various ages, from seedlings to adults.
Most dominant broadleaf species presented inverse J-shaped patterns in size distribution within the plot (Table 3), suggesting a regeneration of these plants. The num-ber of seedling species (with basal diameter smaller than 5.0 cm) found in the sampling plot totaled 42 while stud-ied down logs totaled 41. Thus, both of them shared a simi-lar species composition.
Yellow cypress was the main component on the canopy layer in this area. Only a few broadleaf trees can grow to
Table 2. Comparison of species composition (relative density,
%) on sampled down logs with that in a 0.25-ha plot studied. On down In 0.25-ha
logs plot
Chamaecyparis obtusa var. formosana 59.76 10.07
Schefflera actinophylla 7.62 10.90
Rhododendron formosanum 7.46 20.48
Illicium philippinense 5.00 10.90
Adinandra formosana 2.22 15.55
Rhododendron kawakamii var. flaviflorum 1.67 0.00
Dendropanax pellucidopunctata 1.59 2.10 Trochodendron aralioides 1.51 0.11 Barthea formosana 1.51 10.29 Eurya crenatifolia 1.11 1.72 Vaccinium wrightii 0.95 1.22 Viburnum urceolatum 0.95 0.11 Elaeocarpus japonicus 0.87 1.05 Skimmia arisanense 0.87 3.15 Viburnum sympodiale 0.63 4.26 Neolitsea acuminatissima 0.32 1.11 Ternstroemia gymnanthera 0.24 1.44 Others 5.71 5.54
Total individuals of all species 1260 1807
Table 1. The diameter and length of nine down logs and
aver-age diameter of yellow cypress on each down log in the forest of Yuan-Yang Lake Nature Reserve.
Diameter of Trunk length Average diameter
trunk base (cm) (cm) (mm) Log 1 45 1174 46.3±168.3 Log 2 49 1524 0.9±2.2 Log 3 90 1400 0.7±2.2 Log 4 54 2740 0.3±0.9 Log 5 60 2400 101.0±123.1 Log 6 72 2895 50.0±63.7 Log 7 77 3286 9.5±9.8 Log 8 53 1790 16.0±24.8 Log 9 45 1600 5.1±2.8
reach this layer. Most broadleaf seedlings on down logs were of shrub species. They usually did not reach heights greater than 10 m. Only species such as Elaeocarpus
japonicus and Trochodendron aralioides could reach the
canopy layer. However, they had low relative densities.
Size Classes Distribution of Trees on Down Logs
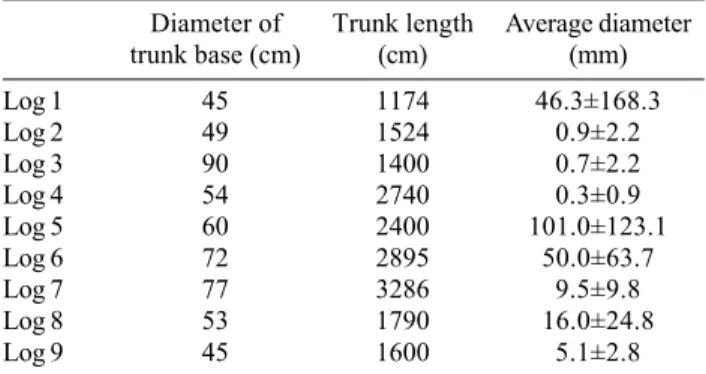
The size class distributions of yellow cypress seedlings on the three groups of down logs (denoted as group I, II, and III) were analyzed. An L-shaped distribution pattern was found for group I (Figure 1a). Abundant seedlings found on new down logs indicated that they provided suit-able open sites for germination of yellow cypress seeds. Most of the seedlings were less than a year old and less than 1 mm in basal diameter. In groups II and III, in
Figure 1. Size class distributions of yellow cypress seedlings
on three groups of down logs. (a): group I, (b): group II, and (c): group III.
Figure 2. Size class distributions of broadleaf individuals
lo-cated on three groups of down logs. (a): group I, (b): group II, and (c): group III.
contrast, most seedlings fell into the 3-9 and 10-20 mm di-ameter classes, respectively (Figure 1b, 1c). As a result, yellow cypress seedlings would establish simultaneously on down logs. Apparently, compared with new down logs, seedlings on older down logs were too crowded to pro-vide space for the germination of new seeds.
The diameter classes of broadleaf trees displayed an in-verse J-shaped distribution on all down log groups, irre-spective of species (Figure 2), suggesting a continuous recruitment of broadleaf trees on down logs. Although there were some variations in the frequency of species present, there was no particular distribution pattern for species found on down logs. Species with a frequency of occur-rence higher than 10 individuals on the studied logs were listed in Table 3.
Distribution Patterns of Species on Down Logs
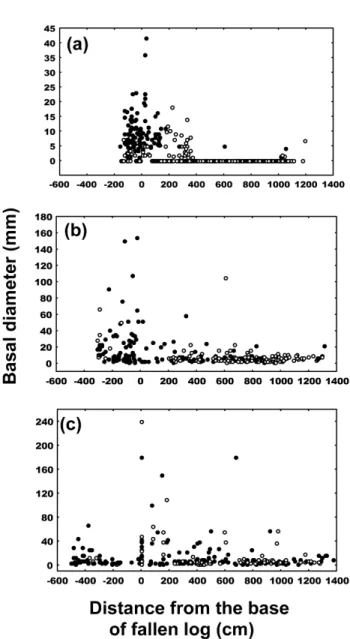
Seedlings of yellow cypress growing on different types of down logs exhibited various temporal and spatial pat-terns (Figure 3). Of group I, seedlings on trunk and branch segments were mostly one-year-old. We found 81, 290, and 184 seedlings of yellow cypress on root, trunk, and branch segments, respectively. Of group II and III, larger and fewer seedlings were found growing on trunk and branch segments. In the former group, we found 12, 41, and 105 seedlings while in the latter we found 16, 12, and 11 seed-lings on root, trunk, and branch segments, respectively. The density of yellow cypress seedlings decreased sig-nificantly from group I to III (Tukey’s test, p<0.01) (Table 4) but increased in size (Tukey’s test, p<0.0001). A num-ber of broadleaf seedlings were found on root segments of group I (Figure 3a). In group II, broadleaf trees were found to invade, particularly on trunk segments (Figure 3b) while they were evenly distributed over different sub-strata in group III (Figure 3c). The average basal diameter of seedlings, irrespective of species, was significantly larger on root segments than on trunk and branch seg-ments in group I (Tukey’s test, p<0.01). However, those grown on groups II and III did not differ significantly. The density of broadleaf species was higher than that of yel-low cypress.
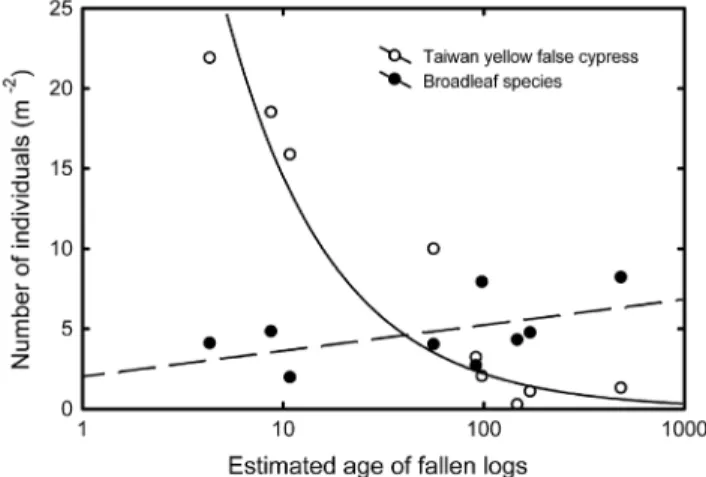
Relation of Seedlings to Age of Down Logs
It was observed that recruitment of yellow cypress seed-lings occurred about one year after the trees fell. Thus, the age of a down log, defined as beginning with a tree’s fall, can be estimated by determining the oldest age of cy-press seedlings grown on it. Analysis of cycy-press density against down log age indicated a negative correlation, sug-gesting high mortality over time (Figure 4). Density of yel-low cypress seedlings decreased from young to old down logs. The regression coefficients declined when the age of down logs increased from 1 to 100 years. The number of seedlings might be higher than 20 per square meter on a ten-year-old down log, but less than four individuals per square meter on a 100-year-old down log.
Figure 3. Spatial distributions of broadleaf species (solid) and
yellow cypress (blank) on three groups of down logs based on their basal diameter. (a): group I, (b): group II, and (c): group III. (The largest yellow cypress, with 780 mm, was eliminated from the diagram of group III down logs).
Table 3. Number of individuals and average basal diameter of broadleaf species occurring on three groups of down logs.
Group I Group II Group III
Species N Diameter (mm) N Diameter (mm) N Diameter (mm)
Rhododendron formosanum 2 13.8±3.0 37 20.7±4.3 55 19.7±3.4 Illicium philippinense 13 9.6±1.5 20 26.8±8.8 30 6.2±2.1 Schefflera taiwaniana 4 13.7±3.2 34 6.7±1.1 58 6.3±0.8 Adinandra formosana 11 10.0±1.7 7 19.9±12.0 10 6.2±1.2 Dendropanax pellucidopunctata 6 20.4±6.4 4 39.7±18.1 10 9.5±3.1 Trochodendron aralioides - - - - 19 13.6±2.4
Rhododendron kawakamii var. laviflorum 21 8.5±1.0 - - -
-Barthea formosana 10 8.1±1.2 4 7.0±1.3 5 8.1±2.9
Vaccinium kengii 1 1.40 5 11.9±4.4 6 14.7±5.7
Elaeocarpus japonicus - - 4 6.8±2.1 7 14.6±6.3
Eurya crenatifolia 2 3.7±0.5 5 7.2±2.0 7 6.1±0.9
Viburnum urceolatum 5 10.3±1.4 3 5.6±2.0 4 11.1±3.6
In contrast to yellow cypress, the density of broadleaf seedlings did not decrease with the age of down logs (Figure 4). In addition, the maximum density of broadleaf seedlings on down logs did not exceed 8 individuals per square meter.
Micro-Habit Variations on Down Logs
Bryophytes and organic matters were unevenly distrib-uted on down logs (Figure 5). Bryophytes generally cov-ered the forest floor and down logs and were scattcov-ered on erect tree trunks. Soon after erect trees fell, bryophytes colonized the new down logs, where they tended to es-tablish dense populations. However, the biomass of bryo-phytes did not differ significantly between different groups of down logs or different segments. The amount of organic matters on a down log might vary, depending upon the segment. The dry weight of organic matters on trunk and branch segments differed significantly in different groups of down logs (Tukey’s test, p<0.01). It was found that the deposition of organic matter on trunk and branch segments proceeded soon after the growth of bryophytes (Figure 5).
No statistical relationship exists between the abundance of bryophytes and average diameter (r2=0.07, p=0.17) or
density (r2<0.01, p=0.95) of woody species on down logs.
The amount of organic matters on down logs, however,
Figure 5. Dry weights of bryophytes (blank) and organic
mat-ter (solid) on three groups of down logs. (a): group I, (b): group II, and (c): group III.
Figure 4. Relationships of densities of broadleaf species (solid)
and yellow cypress (blank) and down log age.
was more closely correlated with average diameter (r2=
0.61, p<0.001), but less with the density (r2=0.17, p<0.05)
of woody species.
Discussion
Since Chamaecyparis is a widely distributed plant and an important timber producer for Taiwan, the development
Table 4. Significance of difference in species number per square meter, density and average diameter between C. obtusa var. formosana
and broadleaf species on three groups of down logs using ANOVA followed by Tukey’s test.
C. obtusa var. formosana Broadleaf species
Species number Density Average Density Average
(Individuals m-2) diameter (mm) (Individuals m-2) diameter (mm)
Group I 1.4±0.4 22.6±5.6 0.5±0.1 3.1±1.6 9.3±0.6
Group II 2.6±0.5 5.9±2.7 4.7±0.5 4.6±1.1 15.2±1.9
Group III 2.7±0.7 1.4±0.5 51.9±21.1 8.3±2.5 10.5±1.0
p value 0.200 <0.01 <0.001 0.139 <0.01
of this temperate forest has received great attention on this island (Su, 1984; Jen, 1995). Yellow cypress is the domi-nant species in the YYL forest. Although it is the main con-tributor to the canopy layer of the forest, broadleaf trees occupy the secondary layer beneath the cypress canopy. In this situation competition between broadleaf trees and cypress, particularly at the seedling stage, might occur, as mentioned by previous authors (Liu and Hsu, 1973). However, the results of our study seem to rule out this possibility because the two plants exhibit quite different types of regeneration. Down logs, particularly those of cy-press trees, provide a habitat especially suitable for the germination and subsequent re-growth of cypress seedlings. As a result, cypress dominates over broadleaf trees in the regeneration process.
Comparisons of seedlings and adult forest composition provide information about forest development (Lertzman, 1992; Poulson and Platt, 1996). In this study, the composi-tion of seedling species corresponded closely to the com-position of adult trees. Thus, replacement of the dominant species by other species seems not to occur. According to pollen analysis (Chen and Wu, 1999), the cypress for-est in YYL Nature Preserve has existed for at least 4,000 years. Thus, we speculate that the vegetation, floristic composition, and canopy structure of this forest will not change in the near future.
Down logs have been reported to promote establish-ment of coniferous seedlings in several forests of the tem-perate climate zone (Christy and Mack, 1984; Veblen, 1986; Harmon and Franklin, 1989; Nakashizuka, 1989; Gray and Spies, 1997). In subtropical areas like Taiwan, no similar study has appeared until now. In YYL Nature Preserve, down logs only occupied ca. 1.3% of the forest floor area (unpublished data). However, they were an important re-generation substrate for yellow cypress. Data from the cen-sus of trees in the study plot indicate that seedlings are abundant on down logs. Analysis of size-class distribu-tion indicates that cypress seeds tend to establish on down logs soon after they fall. Regression analysis revealed that a density-dependent mortality might drive the dynamic growth of yellow cypress seedlings on down logs.
In the forest of YYL, both the yellow cypress and broa-dleaf species co-existed. The predominance of the former plant over the latter one might be related to different gap detecting mechanisms in them, as documented by Swaine and Whitmore (1988). The seedlings of both the yellow cypress and broadleaf trees seem to co-exist on down logs. However, young cypress seedlings grow without compe-tition from broadleaf trees on the trunk and branch seg-ments of new down logs because very few of the latter were found to grow on them. In this case, inter-specific competition is not important for the recruitment of cypress. At the latter stage of recruitment, the yellow cypress might be tall enough to override any invasions by broadleaf trees. As a result, cypress is a stronger competitor than broad-leaf trees on older down logs.
In the forest of YYL, at least two conditions may affect tree recruitment. The first is the light mosaic in the forest.
The growth of yellow cypress in plantations was improved when the light regime was enhanced by pruning or thin-ning the vegetation (Lo-Cho et al., 1999). This indicates that yellow cypress is a shade-intolerant species. In addition, the mound or bole of an uprooted tree provided suitable sites for tree seedlings, preventing competition with bamboo and undergrowth (Nakashizuka, 1989; Abe et al., 2001). Therefore, new down logs might provide a habitat that allows a relatively higher light regime than shaded sites under the canopy. As a result, down logs provide an opportunity for yellow cypress to compete with other species of plants.
The second condition that might affect tree recruitment is the varied microhabitats on a down log. In the forest of YYL, both the epiphytic bryophytes and organic matter on down logs provided a favorable microhabitat for cy-press seedlings. The effects of bryophytes were possi-bly more beneficial than those of organic matter. The high fog-absorbing ability of epiphytic bryophytes was impor-tant for hydrological and nutrient cyclings in this ecosys-tem (Chang et al., 2001). The growth of plants is affected by the water and nutrient contents in their inhabiting sub-strates (Burslem et al., 1995; Burslum and Grubb, 1996). Bryophytes spreading on newly down logs might repre-sent nutrient and hydrological gradients that allow yellow cypress seeds to germinate on trunk and branch segments.
To demonstrate co-existence via niche partitioning in gaps, three premises must be confirmed: the existence of a resource gradient within gaps, tree species performing differently along this gradient, and the contribution of dif-ferences to species coexistence (Burslem, 1996; Brokaw and Busing, 2000). Resource gradients for woody species were not well known on down logs. However, yellow cy-press and broadleaf trees co-existed and showed specific patterns on down logs. Accordingly, two of the three pre-mises hold true for cypress in the forest of YYL. Thus, the niche-partitioning hypothesis is suggested to explain the co-existence of yellow cypress and broadleaf trees on down logs in YYL. Apparently, further study is needed to examine the resource gradients and their possible func-tions on down logs.
Other than the factors mentioned above, the predomi-nance of yellow cypress over broadleaf plants might be related to an allelopathic effect. It is possible that cypress might contain certain kinds of allelochemicals that favor the growth of cypress and inhibit that of other plants. However, further study is necessary.
Gaps are usually filled in by lateral extensive growth of surrounding canopy trees and vertical growth of saplings in the gap (Nakashizuka, 1984). In the forest of YYL, yel-low cypress was the only species creating both canopy gaps and down logs. Gaps were filled almost exclusively by the growth of yellow cypress seedlings under the gaps. Lateral extension of surrounding canopy trees was less important because the canopy layer was not continuous. Therefore, the recovery time of a gap was determined by the growth rate of yellow cypress seedlings. According
to their growth rate, new individuals need about 200 years of growth to overtop the shrub layer and about 500 years to fill up the gap.
Kennedy and Quinn (2001) showed that understory plant species had different establishing abilities on vari-ous substrates. However, in the studied area of YYL, there was no significant difference in the distribution between broadleaf trees on down logs and those on the forest floor. The present study provides an explanation of how impor-tant down logs are to the regeneration of yellow cypress in YYL Nature Preserve.
Acknowledgments. The authors thank the Forest
Conserva-tion and Management AdministraConserva-tion, Veterans Affairs Commission, Executive Yuan, Taiwan, for permission to do this study in Yuan-yang Lake Nature Preserve. The authors want to acknowledge Prof. Horng-Jye Su of the Department of Forestry, National Taiwan University (NTU), Taipei, Taiwan and Prof. Chang-Fu Hsieh in the Department of Botany, NTU for criti-cal reading of the manuscript. We also thank Dr. Shih-Chieh Chang of the Institute of Natural Resources, Dong Hwa University, Hualien and Dr. Chiao-Ping Wang of the Taiwan For-estry Research Institute, Taipei, for their useful discussions. We also appreciate Ms. Hei-Cheng Peng, Shu-Ling Wu, Mr. Po-Chun Cheng, and Ms. Yi-Ling Lai for their help in field experiments.
Literature Cited
Abe, M., H. Miguchi, and T. Nakashizuka. 2001. An interac-tive effect of simultaneous death of dwarf bamboo, canopy gap, and predatory rodents on beech regeneration. Oecologia
127: 281-286.
Arriaga, L. 1988. Gap-dynamics of a tropical cloud forest in Northeastern Mexico. Biotropica 20: 178-184.
Brokaw, N.V.L. 1985. Gap-phase regeneration in a tropical forest. Ecology 66: 682-687.
Brokaw, N. and R. T. Busing. 2000. Niche versus chance and tree diversity in forest gaps. Trends Ecol. Evol. 15: 183-188.
Burselm, D.F.R.P. 1996. Differential responses to nutrients, shade and drought among tree seedlings of lowland tropical forest in Singapore. In M.D. Swaine (ed.), Ecology of Tropi-cal Forest Tree Seedlings. Man and the Biosphere Series. Parthenon, Vol. 17, pp. 211-244.
Burslem, D.F.R.P. and P.J. Grubb. 1996. Response to simulated drought and elevated nutrient supply among shade-tolerant tree seedlings of lowland tropical forest in Singapore. Biotropica 28: 636-348.
Burslem, D.F.R.P., P.J. Grubb, and I.M. Turner. 1995. Re-sponses to nutrient addition among shade-tolerant tree seed-lings of lowland tropical rain forest in Singapore. J. Ecol.
83: 113-122.
Chang, L.M. 1961. Ecological Studies on the Vegetation of Mt. Ta-Yuan. Bull. Taiwan For. Res. Inst. No. 70.
Chang, N.H., Y.R. Hsui, F.W. Horng, H.M. Yu, and F.C. Ma. 2001. Natural seeding and seedlings occurrence in the
Chamaecyparis forest at Chilan Mt. area. Taiwan J. For.
Sci. 16: 321-326.
Chang, S.C., I.L. Lai, and J.T. Wu. 2002. Estimation of fog
depo-sition on epiphytic bryophytes in a subtropical montane forest ecosystem in northeastern Taiwan. Atmospheric Re-search 64: 159-167.
Chen, S.H. and J.T. Wu. 1999. Paleolimnological environment indicated by the diatom and pollen assemblages in an al-pine lake of Taiwan. J. Paleolimnolo. 22: 149-159. Chiu, C.M., C.N. Lo-Cho, and H.H. Chung. 1995. The stem
form and crown structure of natural regeneration stands of
Chamaecyparis taiwanensis in Chi-Lan-Shan Area. Bull.
Taiwan For. Res. Inst. (New Series) 10: 121-130.
Chou, C.H., T.Y. Chen, C.C. Liao, and C.I. Peng. 2000. Long-term ecological research in the Yuanyang Lake forest eco-system I. Vegetation composition and anlysis. Bot. Bull. Acad. Sin. 41: 61-72.
Christy, E.J. and R.N. Mack. 1984. Variation in demography of juvenile Tsuga heterophylla across the substratum mosaic. J. Ecol. 72: 75-91.
Dalling, J.W., S.P. Hubbell, and K. Silvera. 1998. Seed dispersal, seedling establishment and gap portioning among tropical pioneer trees. J. Ecol. 86: 674-689.
Denslow, J.S. 1980. Gap partitioning among tropical rainforest trees. Biotropica 12: 47-55.
Gray, A.N. and T.A. Spies. 1996. Gap size, within-gap posi-tion and canopy structure effects on conifer seedling establishment. J. Ecol. 84: 635-645.
Gray, A.N. and T.A. Spies. 1997. Microsite controls on tree seedling establishment in conifer forest canopy gaps. Ecol-ogy 78: 2458-2473.
Grubb, P.J. 1977. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 52: 107-145.
Harmon, M.E. and J.F. Franklin. 1989. Tree seedlings on logs in Picea-Tsuga forests of Oregon and Washington. Ecology
70: 48-59.
Hung, L.P. 1984. The effect of improvement by selective cut-ting methods for the natural forest of cypress on high moun-tain area in Taiwan. Quar. J. Chin. For. 17: 47-56.
Hwang, Y.H., C.W. Fang, and M.H. Yin. 1996. Primary pro-duction and chemical composition of emergent aquatic macrophytes, Schoenoplectus mucronatus ssp. robustus and
Sparganium fallax, in Lake Yuan-yang, Taiwan. Bot. Bull.
Acad. Sin. 37: 265-273.
Jen, I.A. 1995. Expectation and historical review of cypress (Chamaecyparis spp.) timber production in Taiwan. Bull. Taiwan For. Res. Inst. (New Series) 10: 227-234.
Kennedy, P.G. and T. Quinn. 2001. Understory plant estab-lishment on old-growth stumps and the forest floor in west-ern Washington. For. Ecol. Manage. 154: 193-200. Lawton, R.O. and F.E. Putz. 1988. Natural disturbance and
gap-phase regeneration in a wind-exposed tropical cloud forest. Ecology 69: 764-777.
Lertzman, K.P. 1992. Patterns of gap-phase replacement in a subalpine, old growth forest. Ecology 73: 657-669. Liu, T. 1971. The classification of the Climax Vegetation
Com-munities of Taiwan. II. The Alpine Tundra and Coniferous Forest Formation. Bull. Taiwan For. Res. Inst. No. 203. Liu, T., C.C. Koh, and B.Y. Yang. 1961. Ecological Survey on
Taiwan Important Forest Types. Bull. Taiwan For. Res. Inst. No. 72.
Natural Area Reserve. Bull. Taiwan For. Res. Inst. No. 237. Lo-Cho, C.N., C.M. Chiu, and Y.C. Chen. 1999. Effects of clean-ing and prunclean-ing on natural-regenerated cypress stands. Tai-wan J. For. Sci. 14: 315-321.
Nakashizuka, T. 1984. Regeneration process of climax beech (Fagus crenata Blume) forests IV. Gap formation. Jpn. J. Ecol. 34: 75-85.
Nakashizuka, T. 1989. Role of uprooting in composition and dynamics of an old-growth forest in Japan. Ecology 70: 1273-1278.
Poulson, T.L. and W.J. Platt. 1996. Replacement patterns of beech and sugar maple in Warren Woods, Michigan. Ecol-ogy 77: 1234-1253.
Putz, F.E. 1983. Treefall pits and mounds, buried seeds, and the importance of soil disturbance to pioneer trees on Barro Colorado Island, Panama. Ecology 64: 1069-1074. Ricklefs, R.E. 1977. Environmental heterogeneity and plant
spe-cies diversity: a hypothesis. Amer. Nat. 111: 376-381. Runkle, J.R. 1981. Gap regeneration in some old growth
for-ests of the Eastern United States. Ecology 62: 1041-1051. Runkle, J.R. 1982. Patterns of disturbance in some old-growth
mesic forests of eastern North America. Ecology 63: 1533-1546.
Runkle, J.R. 2000. Canopy tree turnover in old-growth mesic forests of eastern North America. Ecology 81: 554-567. Spies, T.A., J.F. Franklin, and T.B. Thomas. 1988. Coarse woody
debris in Douglas-fir forests of western Oregon and Washington. Ecology 69: 1689-1702.
Su, H.J. 1984. Studies on the climate and vegetation types of the natural forests in Taiwan (II) Altitudinal vegetation zones in relation to temperature gradient. Quar. J. Chin. For. 17: 57-73.
Swaine, M.D. and T.C. Whitmore. 1988. On the definition of ecological species groups in tropical rain forests. Vegetatio
75: 81-86.
Uhl, C., K. Clark, N. Dezzeo, and P. Maquirino. 1988. Vegeta-tion dynamics in Amazonian treefall gaps. Ecology 69: 751-763.
Veblen, T.T. 1986. Treefalls and the coexistence of conifers in subalpine forests of the Central Rockies. Ecology 67: 644-649.
Yamamoto, S. 1988. Seedling recruitment of Chamaecyparis
obtusa and Sciadopitys verticillata in different
microenvi-ronments in an old-growth Sciadopitys verticillata forest. Bot. Mag. Tokyo 101: 61-71.