Lysophospholipids Enhance Matrix Metalloproteinase-2
Expression in Human Endothelial Cells
Wen Ting Wu, Chiung-Nien Chen, Chi Iou Lin, Jiun Hong Chen, and Hsinyu Lee
Department of Life Science (J.H.C., H.L.) and Institute of Zoology (W.T.W., C.I.L., J.H.C., H.L.), National Taiwan University, Taipei, Taiwan 10617; and Department of Surgery (C.-N.C.), National Taiwan University Hospital, Taipei, Taiwan 10051
Lysophosphatidic acid (LPA) and sphingosine 1-phosphate (S1P) are both low-molecular-weight lysophospholipids, which promote cell proliferation, migration, and invasion via interaction with a family of specific G protein-coupled recep-tors. Matrix metalloproteinases (MMPs) are zinc-dependent proteolytic enzymes, which are involved in degradation of the extracellular matrix and play critical roles in endothelial cell migration and matrix remodeling during angiogenesis. Among these MMPs, MMP-2 is known to trigger cell migration. In our present study, we examined the effects of LPA and S1P on MMP-2 expression in human endothelial cells. We showed that LPA and S1P enhanced MMP-2 expression in mRNA, pro-tein levels, and also enzymatic activity of cells of the EAhy926
human endothelial cell line. The enhancement effects oc-curred in concentration- and time-dependent manners. Re-sults from real-time PCR, Western blots, and substrate gels indicated that these enhancement effects were mediated through MAPK kinase/ERK-, nuclear factor-B-, and calcium influx-dependent pathways. Furthermore, we show that en-dothelial cell invasion of the gel was enhanced by lysophos-pholipids, and the induction could be prevented by an MMP inhibitor, GM6001. These observations suggest that LPA and S1P may play important roles in endothelial cell invasion by regulating the expression of MMP-2. (Endocrinology 146: 3387–3400, 2005)
B
IOACTIVE LYSOPHOSPHOLIPIDS (LPLS), including lysophosphatidic acid (LPA) and sphingosine-1-phos-phate (S1P), are membrane-derived lipid mediators, which are generated from phospholipid precursors of membranes and secreted by platelets (1, 2), macrophages, epithelial cells (3–5), and some cancer cells (6 – 8). These lysophospholipids regulate migration, proliferation, and survival of endothelial cells (9 –11). LPA and S1P bind multiple G protein-coupled receptors of the endothelial differentiation gene (Edg) family (9, 12). To date, nine members of the family have been dis-covered in mammalian cells: LPA1 (Edg-2), LPA2 (Edg-4), LPA3(Edg-7), and LPA4are receptors for LPA (13, 14), and S1P1 (Edg-1), S1P2(Edg-5), S1P3 (Edg-3), S1P4(Edg-6), and S1P5(Edg-8) are receptors for S1P (9, 15). Activation of LPL receptors results in a wide spectrum of intracellular events, such as increases in inositol phosphates and intracellular calcium (9), inhibition of adenylyl cyclase (16), and activation of kinases such as protein kinase C and many other signaling pathways (17–19).The major sources of LPA and S1P are activated platelets, injured cells, and cells stimulated by growth factors, sug-gesting potential roles in inflammation, wound healing, and tumor formation (6). The concentration of S1P is estimated to be 200 nm in plasma serum (20), and that of LPA is approx-imately 1–10 m in human serum with variable reports of plasma concentrations from undetectable to 2 nm (21). The growth-promoting activity of LPA has been observed in nu-merous cell lines. The ability of serum to induce mitosis appears to be mediated, in large part, by the lipids LPA and S1P (22).
Endothelial cells are normally quiescent and form a tight monolayer by interacting with the underlying extracellular matrix (ECM) and surrounding endothelial cells (23). Inter-actions between endothelial cells and ECM proteins are im-portant determinants of endothelial cell migration and sig-naling. In addition, endothelial cell migration is important during vascular development and angiogenesis. Both LPA and S1P stimulate endothelial cell migration (22, 24, 25). Matrix metalloproteinases (MMPs) have been reported to play critical roles in this process (26 –28).
MMPs are a family of enzymes that degrade components of the ECM. They are grouped by their substrate preferences and also domain structures: collagenases (MMP-1, MMP-8, and MMP-13) degrade fibrillar collagen; gelatinases (MMP-2 and MMP-9) are potent in nonfibrillar and denatured colla-gen degradation; stromelysins (3, 10, and MMP-11) prefer proteoglycans and glycoproteins as substrates; and membrane-type MMPs (MT1-, MT2-, MT3-, MT4-, and MT5-MMP) contain a C-terminal transmembrane domain direct-ing cell surface localization (26, 29, 30).
The 72-kDa gelatinase A (MMP-2) is the most widely dis-tributed of all the MMPs and is expressed constitutively by First Published Online May 5, 2005
* W.T.W. and C.-N.C. contributed equally to this work
Abbreviations: ECM, Extracellular matrix; Edg, endothelial differen-tiation gene; FBS, fetal bovine serum; GAPDH, glyceraldehyde phos-phate dehydrogenase; HUVEC, human umbilical vein endothelial cell; LPA, lysophosphatidic acid; LPL, lysophospholipid; M199, medium 199; MEK, MAPK kinase; NF-B, nuclear factor-B; MMP, matrix metallo-proteinase; PDTC, pyrrolidinedithiocarbamate; PI3K, phosphatidylino-sitol 3-kinase; PLC, phospholipase C; PP2, 4-amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo(3, 4-d) pyrimidine; S1P, sphingosine 1-phosphate; TIMP, tissue inhibitor of metalloproteinase; uPA, urokinase-type plas-minogen activator.
Endocrinology is published monthly by The Endocrine Society (http:// www.endo-society.org), the foremost professional society serving the endocrine community.
a number of cells, including endothelial cells. It participates in the breakdown of collagen type IV, a major component of subendothelial basement membranes (29). Therefore, MMP-2 production by endothelial or surrounding cells may be vital for the formation of new functional blood vessels, in either the early process of degradation or the later recon-struction of the underlying basement membrane. Paradox-ically, at low levels, the specific physiological MMP-2 in-hibitor, tissue inhibitor of metalloproteinases (TIMP)-2, promotes this activation by forming a membrane complex with MT1-MMP, anchoring the pro-MMP-2 to the cell surface (31–33). A resent study also showed that both TIMP-2 and TIMP-3 enhance the activation of pro-MMP-2 by MT3-MMP (34). It has been shown that MMP-2 and TIMP-2 are required for normal development of zebrafish embryos (35, 36), in-dicating that MMP-2 and TIMP-2 expressions are essential during embryogenesis.
Activities of MMPs are tightly regulated at several levels including gene expression and secretion of proenzymes that require activation by other MMPs or serine proteinases (37). Once MMPs are activated, they are subjected to inhibition by TIMPs, which determine the net enzymatic activity in the extracellular space (26). A balance between MMPs and TIMPs is involved in the morphogenesis of normal tissue as well as mechanisms of tumor invasion and metastasis (37– 40).
Aberrant expression of MMPs has been noted in cancer, angiogenesis (41), arthritis, inflammation, emphysema, multiple sclerosis (42), and chronic wounds (6, 8). In this study, we hypothesized that LPA and S1P may affect en-dothelial cell invasion through regulating MMP-2 expres-sion. We present evidence that LPA and S1P enhance MMP-2 expression in human endothelial cell hybridoma EAhy926 cells in concentration- and time-dependent man-ners. By using chemical inhibitors, we show that LPA and S1P enhance MMP-2 through pathways dependent on MAPK kinase (MEK)/ERK-, nuclear factor-B (NF-B), and calcium influx-mediated signaling. Furthermore, we used a modified Boyden chamber to investigate the inva-sive activity of EAhy926 cells and found that the invainva-sive activity was enhanced by LPL treatments. This enhance-ment was likely due to an increase in MMP-2 expression in EAhy926 cells because the enhancement was prevented by pretreatment with the synthetic MMP inhibitor, GM6001. These results imply that LPL’s enhancement of endothelial cell invasion is likely mediated through in-duction of MMP-2 expression.
Materials and Methods Reagents
S1P, oleoyl-LPA, and proteinase inhibitors, including leupeptin, so-dium vanadate, phenylmethylsulfonyl fluoride, and -glycerophos-phate, pertussis toxin, and pyrrolidinedithiocarbamate (PDTC) were purchased from Sigma (St. Louis, MO). Gelatin was obtained from United States Biochemical Corp. (Cleveland, OH). The chemical inhib-itors, Y27632 dihydrochloride, U73122, PD98059, and wortmannin, were purchased from Tocris Cookson (Ellisville, MO). 4-Amino-5-(4-chloro-phenyl)-7-(t-butyl)pyrazolo(3, 4-d) pyrimidine (PP2) was purchased from Calbiochem (San Diego, CA). Monoclonal mouse antibody against human MMP-2 was obtained from Santa Cruz Biotech (Santa Cruz, CA). Fetal bovine serum (FBS) was obtained from Hyclone Laboratories
(Lo-gan, UT). Medium 199 (M199) and trypan blue stain were purchased from Life Technologies, Inc. (Gaithersburg, MD). TRIzol, penicillin-streptomycin, and trypsin-EDTA were purchased from Invitrogen (Grand Island, NY). The MMP inhibitor, N-[(2S)-2-(methoxycarbonyl-methyl)-4-methylpentanoyl]-l-tryptophan-methylamide, GM6001, was purchased from Chemicon (Temecula, CA).
Cell culture
Human umbilical cords were kindly provided by Dr. S. M. Lee (Chun-Shan Hospital, Taipei, Taiwan). Each umbilical cord was washed several times with 1⫻ cord buffer [136.9 mm NaCl, 4 mm KCl, 10 mm HEPES, and 11.1 mm glucose (pH 7.65)] and then was incubated with 0.1% type IV collagenase (Sigma) in a 37 C incubator for 10 min. After incubation, the endothelial cells were washed out with serum-free M199 and cul-tured with 20% endothelial cell growth medium (Cell Applications Inc., San Diego, CA), in M199 with 5 mm l-glutamine, 1.5 g/liter sodium bicarbonate, 4.5 g/liter glucose, 1.0 mm sodium pyruvate, penicillin (100 IU/ml), streptomycin (100 IU/ml), and 10% FBS. Culture dishes were maintained in a humidified 5% CO2at 37 C. After 16 h, nonadherent cells
were removed, and the adherent cells were harvested. Experiments were performed with cells that had undergone no more than five passages. Endothelial cell hybridoma Eahy926 cells were kindly provided by Dr. Cora-Jean S. Edgell (Department of Pathology, University of North Carolina, Chapel Hill, NC) (43). Eahy926 cells were cultured in M199 supplemented with 10% FBS, penicillin (100 IU/ml), streptomycin (100 IU/ml), and glutamine (5 mm) in a humidified 5% CO2at 37 C. Cells
were cultured in six-well plates (Greiner Bio-One, Kremsmu¨nster, Aus-tria) for the experiments.
LPL stimulation
LPA and S1P were prepared in chloroform and methanol (1:9) and stored at⫺20 C. Endothelial cells were cultured to 70–80% confluence and starved in serum-free medium for 16 –24 h. LPA and S1P treatments were prepared in serum-free M199 with 0.1% fatty acid-free BSA as a carrier.
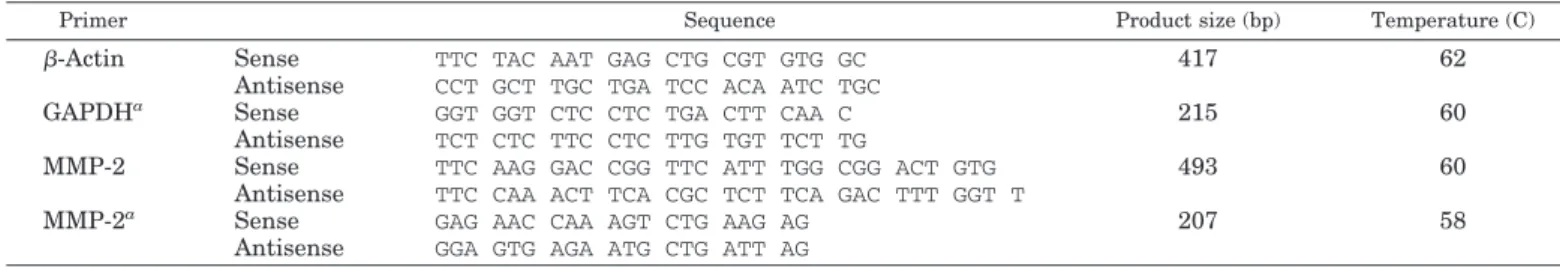
RNA extraction, RT-PCR, cloning, and DNA sequencing Total RNA from endothelial cells was isolated using the TRIzol re-agent (Invitrogen), and 1g of total RNA was reverse-transcribed using reverse transcriptase enzyme (New England Biolabs, Beverly, MA). The generated cDNAs were subjected to PCR using primer sets for MMP-2 and edg receptors. Coamplification of-actin or glyceraldehyde phos-phate dehydrogenase (GAPDH) was performed as the internal control. Sequences of the oligonucleotide primers, and the expected size and annealing temperature for the reactions are summarized in Tables 1 and 2. PCR products were resolved on 1.5% agarose gels, stained with ethidium bromide, and photographed. All data were quantified with TotalLab software (version 2.01; Newcastle upon Tyne, UK). PCR prod-uct was extracted from agarose gel and cloned by TA-cloning kit (In-vitrogen, Carlsbad, CA). Cloned cDNAs were sequenced by Mission Biotech Inc. (Taipei, Taiwan) to verify the identities of the sequences. Real-time PCR
Real-time PCR was performed using the iCycler iQ real-time detec-tion system (Bio-Rad Laboratories, Hercules, CA) with SYBR-green I (stock solution⫻ 25,000, diluted at 1:1000) as the fluorescent dye en-abling real-time detection of PCR products according to the manufac-turer’s protocol. Gene-specific primers were used, and the specificity was tested under normal PCR conditions. The cDNA was subjected to real-time PCR using the primer pairs as given in Table 1. Cycling con-ditions were 95 C for 3 min, followed by 40 cycles of 94 C for 30 sec, 62 C for 30 sec, and 72 C for 60 sec. For quantification, the target gene was normalized to the internal standard GAPDH gene. Oligonucleotide primers for PCR were designed using Beacon Designer2 software (Pre-mier Biosoft International, Palo Alto, CA).
Western blotting
Endothelial cell lysates were prepared as previously described. Briefly, control and LPA- or S1P-treated cells were washed with ice-cold
PBS twice and lysed with 200 l of radioimmunoprecipitation assay buffer [50 mm Tris (pH 8.0), 150 mm NaCl, 1% Triton X-100, 0.5% Na deoxycholate, and 0.1% sodium dodecyl sulfate] in the presence of proteinase inhibitors (10g/ml leupeptin, 2 mm sodium vanadate, 1 mm phenylmethylsulfonyl fluoride, and 2 mm-glycerophosphate). Lysates were collected with a cell scraper and clarified by centrifugation (10,000⫻ g for 10 min at 4 C). Protein concentrations were determined using the Bio-Rad protein assay.
Equal amounts of protein (60g) were separated by 4–10% SDS-PAGE and then transferred to Immobilon-nitrocellulose transfer mem-branes (Millipore, Billerica, MA). Transferred blots were blocked with 5% BSA in TBS and Tween 20 [10 mm Tris, 150 mm NaCl, and 0.1% Tween-20 (pH 7.4)] for 2 h (room temperature) before incubation with mouse monoclonal antibody against human MMP-2 (1:1000 dilution; Santa Cruz Biotechnology). Membranes were then washed and incu-bated with horseradish peroxidase-conjugated goat antimouse antibody (1:2000). The blot was then incubated in ECL substrate (Santa Cruz chemiluminescence) for 1 min at room temperature, and visualized by autoradiography.
Blots were stripped and reprobed with an antibody against human -actin to demonstrate uniform loading of proteins. All data were quan-tified with TotalLab version 2.01 software.
Gelatin zymography
EAhy926 cells and human umbilical vein endothelial cells (HUVECs) were serum starved for at least 16 h before LPL treatment in serum-free medium. Conditioned media collected from both control (untreated) and LPL-treated cells were diluted 1:1 in nonreducing sample buffer and analyzed for proteinase activity by substrate gel zymography. Total protein concentrations were determined using the Dc protein assay (Bio-Rad). Identical amounts of supernatant (10 l) were electropho-resed under nonreducing conditions using a 10% polyacrylamide gel containing 0.1% gelatin for 120 min at 110 V. After electrophoresis, the gel was washed twice in 2.5% Triton X-100 for 30 min to remove sodium dodecyl sulfate. After incubation at 37 C in developing buffer [50 mm Tris-HCl (pH 7.5), 50 mm NaCl, 10 mm CaCl2䡠 2H2O, and 0.02% NaN3]
for 16 h, the gels were stained for 1 h with 0.25% Coomassie blue R 250
and destained with 7% acetic acid. The presence of gelatinolytic activity was identified as clear bands on a uniform blue background. Incubation of the gel with 5 mm EDTA in developing buffer significantly inhibited the enzymatic activity of the gelatinase, indicating that the responsible gelatinase was a metalloproteinase. To confirm that detected activities were zinc-dependent gelatinases, some zymogram gels were incubated with 1 mm 1, 10-O-phenanthroline (Calbiochem), a broad-spectrum in-hibitor of metalloproteinases.
Immunoprecipitation
EAhy926 cells were serum starved for at least 16 h before LPL treat-ment in serum-free medium. Conditioned media collected from both control (untreated) and LPL-treated cells were centrifuged at 5000 rpm for 10 min at 4 C. Anti-MMP-2 antibodies (1:500 dilution; Santa Cruz Biotechnology) were added to 0.5 ml of supernatants and incubated for 24 h at 4 C. The immunocomplexes were separated by incubating with 30l of Protein G Plus/Protein A agarose suspension (Calbiochem) for 4 h at 4 C followed by centrifugation at 5000 rpm for 5 min at 4 C. The pellets were mixed with nonreducing sample buffer and processed for gelatin zymography.
Invasion assay
The invasive properties of endothelial cells were assayed in a mod-ified Boyden chamber with a membrane filter. In brief, the upper surface of polycarbonate filters with 8-m pores (Neuro Probe, Gaithersburg, MD) was precoated with 2% gelatin and dried at room temperature. The prepared filters (gelatin-coated filters) were extensively rehydrated with PBS before use. Confluent EAhy926 cells were starved with serum-free M199 for 16 h. Cells were then trypsinized with trypsin-EDTA, followed by incubating with M199 in the presence of 1% FBS to inhibit trypsin activity. Cell pellets were then resuspended in serum-free media to a concentration of 1⫻ 106cells/ml. Control and different-concentration
LPL treatments were loaded in the lower chambers of the modified Boyden chamber as a chemotaxis inducer. Cells (2⫻ 105cells/ml at 50
l per well) were plated in the upper chambers and allow to invade through the 8-m porous 2% gelatin-coated filters at 37 C for 4 h. TABLE 2. Human Edg receptor primer sets
Primers Sequences Product size (bp) Temperature (C)
edg1 S1P1 5⬘-GACTCTGCTGGCAAATTCAAGCGAC 352 62 3⬘-ACCCTTCCCAGTGCATTGTTCACAG edg5 S1P2 5⬘-CTCTCTACGCCAAGCATTATGTGCT 512 62 3⬘-TCAGACCACCGTGTTGCCCTC edg3 S1P3 5⬘-CAAAATGAGGCCTTACGACGCCA 701 61.2 3⬘-TCCCATTCTGAAGTGCTGCGTTC edg6 S1P4 5⬘-AGCCTTCTGCCCCTCTACTC 339 62 3⬘-GTAGATGATGGGGTTGACCG edg8 S1P5 5⬘-GGAGTAGTTCCCGAAGGACC 236 59.7 3⬘-TCTAGAATCCACGGGGTCTG
edg2 LPA1 5⬘-CGGAGACTGACTGTCAGCAC 397 62
3⬘-GGTCCAGAACTATGCCGAGA
edg4 LPA2 5⬘-AGCTGCACAGCCGCCTGCCCCGT 888 53
3⬘-TGCTGTGCCATGCCAGACCTTGTC
edg7 LPA3 5⬘-TTAGCTGCTGCCGATTTCTT 392 62
3⬘-ATGATGAGGAAGGCCATGAG TABLE 1. Human primer sets
Primer Sequence Product size (bp) Temperature (C)
-Actin Sense TTC TAC AAT GAG CTG CGT GTG GC 417 62
Antisense CCT GCT TGC TGA TCC ACA ATC TGC
GAPDHa Sense GGT GGT CTC CTC TGA CTT CAA C 215 60
Antisense TCT CTC TTC CTC TTG TGT TCT TG
MMP-2 Sense TTC AAG GAC CGG TTC ATT TGG CGG ACT GTG 493 60
Antisense TTC CAA ACT TCA CGC TCT TCA GAC TTT GGT T
MMP-2a Sense GAG AAC CAA AGT CTG AAG AG 207 58
Antisense GGA GTG AGA ATG CTG ATT AG aPrimer sets designed for real-time PCR.
Noninvading cells on the upper surface of the filter were wiped off with a cotton swab, and migrating cells adherent to the lower surface were fixed with 20% formaldehyde and stained with a 1% crystal violet solution in 5% ethanol. Cells that had invaded to the lower surface were quantified by colorimetric measurement using crystal violet staining. Adhesion assay
The assays were modified and performed as previously described (44). Briefly, untreated polystyrene 96-well flat-bottom microtiter plates (Greiner Bio-One) were coated with 2% gelatin in the presence or ab-sence of GM6001. Wells coated at 37 C for 1 h were washed with PBS [PBS: 137 mm NaCl, 2.7 mm KCl, and 10 mm Na2HPO4(pH 7.4)].
EAhy926 cells were starved overnight in serum-free medium and then treated with control and 1m of LPA and S1P as indicated for 4 h. Cells were trypsinized and than coincubated with media in the presence or absence of 10m GM6001 in 1 h gelatin-precoated 96 wells for 45 min. As a negative control, media alone were loaded into the wells. Nonad-herent cells were vigorously washed by PBS three times. The attached cells were fixed and stained with a crystal violet solution [1% (wt/vol) crystal violet, 10% (vol/vol) ethanol] for 30 min at room temperature. After washing with PBS, adherent cells were solubilized by solubiliza-tion buffer [50% (vol/vol) 0.1 m NaH2PO4(pH 4.5) and 50% (vol/vol)
ethanol] overnight at room temperature. Adherence was determined by absorption at 595 nm in a microplate reader (Bio-Rad Laboratories). The data were reported as the mean absorbance of triplicate wells⫾ se minus the mean absorbance of media alone loaded wells.
GM6001 treatments
Cells were then trypsinized with trypsin-EDTA, followed by incu-bating with M199 in the presence of 1% FBS to inhibit trypsin activity. Cell pellets were then resuspended in serum-free media to a concen-tration of 1⫻ 106cells/ml. The suspended cells were incubated in control
medium or 10m GM6001, a broad MMP inhibitor, at 37 C for 1 h. Serum-free M199 control, 1m LPA, and 1 m S1P treatments were loaded in the lower chambers of the modified Boyden chamber. Cells (5⫻ 104/well, in either control medium or 10m GM6001) were plated
in the upper chambers and allowed to invade through the 8-m porous filters coated with 2% gelatin for 4 h. Nonmigrating cells were removed, and the filter was fixed with 20% formaldehyde and stained with 1% crystal violet solution in 5% ethanol.
Chemical inhibition
A specific inhibitor of Gi (PTx, 15 ng/ml), a phospholipase C (PLC) inhibitor (U73122, 1m), a MEK/ERK inhibitor (PD98059, 10 m), a phosphatidylinositol 3-kinase (PI3K) inhibitor (LY 294002, 20m), an NF-B inhibitor (PDTC, 100 m), a Src kinase inhibitor (PP2, 10 m), and an inhibitor of receptor-mediated calcium entry (SKF 96365, 10m) were used in our assays to determine the signaling pathways involved. Cells were cultured to around 80% confluency and starved overnight in se-rum-free medium. All inhibitors were applied 1 h before LPL treatments. Statistical analysis
Significant differences between control and treatment groups were tested using one-way ANOVA followed by Fisher’s protected least-significant differences test (StatView, Abacus Concept, Berkeley, CA) and two-way ANOVA followed by Duncan’s new multiple range test. Each experiment was repeated at least three times. A value of P⬍ 0.05 was considered statistically significant.
Results
LPA and S1P enhance MMP-2 mRNA levels in human endothelial cells
We first determined the Edg receptor expression profile of EAhy926 cells by RT-PCR (Fig. 1). Our results indicated that these cells express S1P1, S1P5, LPA1, and LPA3receptors and therefore can be used as a good model to study LPL’s effects in human endothelial cells.
Dose response. LPLs have multiple effects in many different cell types, such as promoting cell migration, which requires protease. Therefore, we were interested in determining whether LPLs regulate MMP-2 expression in endothelial cells. We first investigated the effects of LPLs on MMP-2 mRNA levels in HUVECs. HUVECs were treated with 1m of LPA and S1P for 4 h. RNAs from treated cells were isolated and subjected to RT-PCR using specific primer sets for hu-man MMP-2 and -actin. MMP-2 mRNA levels were ele-vated after the treatments, especially after treatment with LPA (Fig. 2A, left panel). Expression patterns of-actin, which was used as the loading control, did not significantly differ in either LPA- or S1P-treated samples (Fig. 2A, right panel). To further confirm that the observed bands were MMP-2 mRNA, PCR products from agarose gels were cloned and sequenced. Sequencing results verified the bands are human MMP-2. We then checked whether the same effects could be detected in EAhy926 cells. Cells were treated with different concentrations of LPA and S1P. RNA from treated EAhy926 cells was harvested and subjected to RT-PCR using specific primer sets for human MMP-2 or GAPDH. The expression patterns of GAPDH, which was used as the loading control, did not significantly differ in either LPA- or S1P-treated samples. LPA and S1P also up-regulated MMP-2 mRNA expression in EAhy926 cells in concentration-dependent manners (Fig. 2B). The MMP-2 mRNA level increased at 0.1 m, and the expression peaked at 1 m of S1P treatment and around 0.5–1m of LPA. Therefore, HUVECs and EAhy926 cells have similar effects after LPL treatment. HUVECs can undergo only five passages. On the other hand, EAhy926 cells can passage through up to 20 generations and have been shown to express S1P1, S1P5, LPA1, and LPA3 receptors (Fig. 1). Therefore, we chose EAhy926 cells as a suitable model for the following experiments.
Time course. In the dose-response experiments, we showed that the enhancement of MMP-2 mRNA expression by both LPA and S1P in EAhy926 cells was concentration dependent. There-fore, we intended to investigate whether the effects of LPA and S1P on MMP-2 mRNA levels in EAhy926 cells were time de-pendent. EAhy926 cells were incubated with LPA (1m) and S1P (1m) for different time intervals as indicated, and RNA FIG. 1. Edg receptor expression profile in EAhy926 cells. cDNA of EAhy926 cells was analyzed by RT-PCR to determine the Edg receptor (LPL receptor) expression profile. GAPDH was used to confirm the cDNA quality. PCR products were separated by 1.5% agarose gel electrophoresis. The expected PCR products were 352, 397, 392, 236, and 215 bp for Edg 1, 2, 7, 8, and GAPDH, respectively. EAhy926 cells express Edg 1 and 8 (S1P 1 and 5) and Edg 2 and 7 (LPA 1 and 3). M, 100-bp marker; G, GAPDH.
was subjected to RT-PCR using specific primer sets for human MMP-2 or GAPDH. Enhanced MMP-2 mRNA in EAhy926 cells was first observed as early as 1 h after treatments were initiated, peaked at 4 h, and then declined thereafter. We show that LPA and S1P up-regulated MMP-2 mRNA expression in EAhy926 cells in a time-dependent manner (Fig. 2C).
LPA and S1P enhance MMP-2 protein levels in human endothelial cells
Dose response. Because the mRNA levels were increased by LPLs, we further investigated whether the elevated mRNA levels of MMP-2 were also correlated with protein expres-sion levels. Cells were treated with different concentra-tions of LPA and S1P, and total protein was collected and
detected by Western blot analysis. After a 4-h treatment, LPA and S1P both enhanced MMP-2 protein expression in concentration-dependent manners (Fig. 3A). Human -ac-tin Western blot-ac-ting was used to show that the amount of sample loading did not significantly differ between con-trol and treated samples. Consistent with the RT-PCR re-sults, the enhancement effects of LPA and S1P on MMP-2 expression in EAhy926 cells peaked at 1 m. For LPA treatment, MMP-2 protein expression appeared to peak at 0.5m and dropped at 5 m, whereas for S1P, the peak was around 0.5–1m, then it dropped at 10 m. Similar results were also observed in HUVECs (data not shown). Time course. In the dose-response experiments, we showed that enhancement of MMP-2 protein levels by both LPA and FIG. 2. LPA and S1P enhance MMP-2 mRNA levels in
HUVECs and EAhy926 cells. A, RNA was isolated from HUVECs treated with 1Mof LPA and S1P in serum-free medium and subjected to RT-PCR using specific primer sets for human MMP-2 or-actin. PCR products were separated by 1.5% agarose gel electrophoresis. The expected PCR products were 493 bp for MMP-2 and 417 bp for -actin. B, EAhy926 cells were serum starved overnight and incubated with LPA or S1P for 4 h at various concentrations as indicated. C, EAhy926 cells were starved overnight and incubated with S1P (1M) and LPA (1M) for different time durations. RNA from treated cells was harvested and subjected to RT-PCR using specific primer sets for human MMP-2 or GAPDH. The reaction products were separated on a 1.5% agarose gel and photographed. A 100-bp DNA ladder was used as a marker. The expected PCR products were 207 bp for MMP-2 and 215 bp for GAPDH. Similar experiments were repeated at least three times, and a representative result is shown in the figure. Each bar of the histogram represents a minimum of three experiments, and data are presented as the mean⫾SD. *, Statistically different, compared with the level in control cells (P⬍ 0.05).
S1P in EAhy926 cells was concentration dependent. There-fore, we wanted to test whether the effects of LPA and S1P on MMP-2 protein expression were time dependent. EAhy926 cells were incubated with LPA (1m) and S1P (1 m) for different time durations as indicated, and total pro-tein was subjected to Western blotting using an anti-MMP-2 antibody. A human-actin antibody was used to show that the amount of sample loading did not significantly differ between control and treated samples. These enhanced MMP-2 protein levels in EAhy926 cells were first observed as early as 1 h after treatment was initiated, peaked at around 4 h for S1P treatment and 4 – 8 h for LPA treatment, and declined thereafter. With overnight treatment, MMP-2 ex-pression remained high in S1P-treated but returned to basal levels in LPA-treated samples. Therefore, we have shown that LPA and S1P up-regulate MMP-2 protein expression in EAhy926 cells in a time-dependent manner (Fig. 3B). LPA and S1P enhance MMP-2 enzymatic activity in human endothelial cells
Dose response. Because protein levels were increased by LPLs, we further investigated whether the elevated protein levels of MMP-2 were also correlated with enzymatic activity. We next investigated the effects in both HUVECs and EAhy926
cells. Cells were treated with different concentrations of LPA and S1P, and conditioned medium was collected and re-solved on a substrate gel (containing 0.1% gelatin). After staining, increased gelatinolytic activity was detected, and we found that LPLs induced MMP-2 activity in a concen-tration-dependent manner both in HUVECs (Fig. 4A) and EAhy926 cells (Fig. 4B). These results were consistent with the results of mRNA levels. HUVECs and EAhy926 cells showed similar results after LPL treatment in these assays. MMP-2 enzymatic activity first increased at 0.5m, and the activities increased with the concentration. The effect of S1P was more significant than that of LPA in regulating MMP-2 enzymatic activity. These results indicated that LPA and S1P induce endothelial cell proteolytic activity, which might stimulate the invasive capacity of endothelial cells. Incuba-tion of the gel with EDTA significantly inhibited the enzy-matic activity of the gelatinase, indicating that the respon-sible gelatinase was a metalloproteinase (data not shown). Time course. In the dose-response experiments, we show that the enhancement of MMP-2 enzymatic activity by both LPA and S1P in EAhy926 cells was concentration dependent. We further investigated whether the effects of LPA and S1P on MMP-2 enzymatic activity were time dependent or due to starvation. Cells were simultaneously subjected to LPL treat-FIG. 3. LPA and S1P enhance MMP-2 protein
expression in EAhy926 cells in concentration-and time-dependent manners. A, EAhy926 cells were starved overnight and incubated with S1P and LPA for 4 h at various concen-trations as indicated. B, EAhy cells were in-cubated with S1P (1M) and LPA (1M) for different time durations. Total cell lysates were collected, and MMP-2 protein expression levels were monitored by Western blotting us-ing a human monoclonal MMP-2 anti-body. A human-actin antibody was used as the loading control. Similar experiments were repeated at least three times, and a represen-tative result is shown in the figure. Each bar of the histogram represents a minimum of three experiments, and data are presented as the mean⫾SD.
ments or serum-free medium for different time durations. The conditioned media, including LPL-treated and control media, were collected and assayed on a substrate gel (con-taining 0.1% gelatin). LPLs induced MMP-2 activity in a time-dependent manner (Fig. 4C). The enhancement effects of LPA and S1P were first observed at 2 h after ligand treat-ment. Although control medium showed a slight increase in MMP-2 activity, LPL treatment generated a more-significant induction of MMP-2 enzymatic activity. Therefore, we sub-tracted the control value to quantify the increased multiples of MMP-2 activity stimulated by LPLs. It should be noted that in addition to starvation before treatment, cells in the overnight treatment were incubated in a serum-free condi-tion for 48 h. Such a stressful situacondi-tion may have caused a dramatic increase in cell secretion in general. That might be the reason that the conditioned medium collected in the overnight treatment showed significantly increased MMP-2 gelatinolytic activity.
Enhancement of MMP-2 gelatinolytic activity by LPL treatments is suppressed by GM6001 and 1, 10-O-phenanthroline. Because the LPLs showed enhancement effects on gelatinolytic ac-tivity in endothelial cells, we further confirmed whether LPLs enhance gelatinolytic activity by up-regulating MMP-2 enzymatic activity in endothelial cells. Cells were pretreated with GM6001 for 1 h, followed by control and 1m of LPA and S1P treatments for 4 h. Conditioned media were col-lected and resolved on a substrate gel. We found that LPL induction of MMP-2 enzymatic activity of the gelatinase was partially blocked by GM6001 (Fig. 5A). In addition, incuba-tion of the gel with 1 mm of 1, 10-O-phenanthroline, a che-lator of zinc (45), also significantly inhibited the enzymatic activity of MMP-2 (Fig. 5B), indicating that the responsible gelatinase was MMP-2. To further confirm that the observed bands in the 62-kDa gelatinolytic activities reflected MMP-2 activation, conditioned media harvested from Eahy926 cells were immunoprecipitated with the anti-MMP-2 antibody. FIG. 4. LPA and S1P enhance MMP-2 activity
in HUVECs and EAhy926 cells in concentra-tion-dependent manners. HUVECs (A) and EAhy926 cells (B) were starved overnight in serum-free medium and incubated with S1P and LPA at various concentrations as indi-cated for 4 h. (C) EAhy926 cells were starved overnight in serum-free medium and incu-bated with 1M of S1P (left panel) or LPA (right panel) for different time durations. We simultaneously treated cells with LPA and S1P or control (serum-free) medium. Condi-tioned media were collected and subjected to substrate gel. After staining with 0.25% Coo-massie blue R 250, increased gelatinolytic ac-tivity was detected. Similar experiments were repeated at least three times, and a represen-tative result is shown in the figure. Each bar of the histogram represents a minimum of three experiments, and data are presented as the mean⫾SD. *, Statistically different, com-pared with the level in control cells (P⬍ 0.05).
Consistent with the gelatin zymography results, conditioned media immunoprecipitated with the anti-MMP-2 antibody but not with normal mouse IgG generate a sharp band at the size of 62 kDa in gelatin zymography (Fig. 5C). These results confirmed LPA and S1P enhance gelatinolytic activity in human endothelial cells through up-regulating MMP-2 en-zymatic activity
LPLs effects on MMP-2 mRNA expression are involved in MEK/ ERK-, NF-B-, and calcium influx-dependent signaling pathways. Because LPA and S1P are ligands for Edg receptors and at least four different Edg receptors are expressed on EAhy926 cells (Fig. 1), we further investigated whether the effects of
LPLs on MMP-2 expression are mediated through these re-ceptors. To investigate the signaling pathway involved in the induction of MMP-2 by LPA and S1P, we used several chem-ical inhibitors known to impede certain signaling pathways. Cells were pretreated with pertussis toxin (a Gi blocker), PD98059 (a MEK blocker), U73122 (a PLC blocker), LY 294002 (a PI3K blocker), PDTC (an NF-B blocker), PP2 (an Src family kinase blocker), and SKF 96365 (a receptor-mediated calcium influx blocker) for 1 h and then were treated with control, 1m LPA, and 1 m S1P for 4 h. After the reverse transcription reaction, we used real-time PCR to detect mod-ulation of MMP-2 mRNA levels by these inhibitors as en-hanced by LPLs. Pretreatment using chemical inhibitors for 1 h had no significant effect on the GAPDH loading control. However, the stimulatory effects of 1 m LPA and S1P on MMP-2 mRNA expression in EAhy926 cells were signifi-cantly suppressed by PD98059, PDTC, and SKF96365. On the other hand, pretreatment with U73122, LY 294002, PP2, and pertussis toxin had no effect on either control or LPL-en-hanced MMP-2 expression (Fig. 6). These results suggested that the enhancement effects of LPLs on MMP-2 mRNA expression are MEK, NF-B, and calcium influx dependent but independent of the function of Gi, PLC, PI3K, and Src family kinases.
FIG. 5. Enhancement of MMP-2 gelatinolytic activity by LPA and S1P treatments is partially suppressed by GM6001 and 1, 10-O-phenanth-roline. A, EAhy926 cells were starved overnight in serum-free medium and pretreated with GM6001 for 1 h and then treated with control and 1Mof LPA and S1P for 4 h. Conditioned media were collected and subjected to gelatin zymographic analysis. B, EAhy926 cells were starved overnight in serum-free medium and treated with control and 1 M of LPA and S1P for 4 h. Conditioned media were collected and subjected to substrate gel. Gels were incubated with developing buffer containing with or without 1 mMof MMP inhibitor 1, 10-O-phenanth-roline. After staining with 0.25% Coomassie blue R 250, increased gel-atinolytic activity was detected. C, Gelatin zymographic analysis of MMP-2 immunoprecipitated with the specific anti-MMP-2 antibody or normal mouse IgG from collected conditioned media as indicated.
FIG. 6. LPA and S1P induction of MMP-2 mRNA expression in EAhy926 cells is mediated through MEK-, NF-B- and calcium influx-dependent mechanisms. EAhy926 cells were pretreated with control media alone, 15 ng/ml of PTx (a Gi inhibitor), 10Mof PP2 (an Src family kinase inhibitor), 20Mof LY 294002 (a PI3K inhibitor), 100 Mof PDTC (an NF-B inhibitor), 1 Mof U73122 (a PLC inhibitor), 10 MPD98059 (a MEK/ERK inhibitor), and 10 MSKF96365 (a calcium influx inhibitor) for 1 h. Inhibitor-treated cells were then treated with 1Mof LPA or S1P for 4 h. MMP-2 mRNA expression levels were monitored by real-time PCR. Histograms represent quan-tification by real-time analysis. Multiple increases, compared with untreated cells, are shown. The average value for each sample was normalized with the amount of GAPDH. Each bar of the histogram represents a minimum of three experiments, and data are presented as the mean⫾SD. These results were analyzed by two-way ANOVA. A level of P ⬍ 0.01 was accepted as significant. **, Statistically different compared with the level in control cells (P⬍ 0.01).
LPLs effects on MMP-2 protein expression are involved in MEK/ ERK-, NF-B-, and calcium influx-dependent signaling pathways. Because the enhancement effects on mRNA levels by LPLs were blocked by the above mentioned chemical inhibitors, we further investigated whether these inhibitors also affected MMP-2 protein expression induced by LPLs. Cells were pre-treated with the inhibitors mentioned above for 1 h, followed by control and 1 m of LPA and S1P treatments for 4 h. Human -actin Western blotting was used to indicate that sample loadings were even. Consistent with the mRNA re-sults, we found that LPL induction of MMP-2 protein ex-pression was blocked by PDTC, PD98059, and SKF96365 but not pertussis toxin. These results suggested that the enhance-ment effects of LPLs on MMP-2 protein expression are MEK, NF-B, and calcium influx dependent but independent of the function of Gi (Fig. 7).
Enhancement of MMP-2 gelatinolytic activity by LPL treatments is partially suppressed by chemical inhibitors. Because the en-hancement effects on mRNA and protein levels by LPLs were blocked by certain chemical inhibitors, we further investi-gated whether the effects of these inhibitors were also cor-related with enzymatic activity. Cells were pretreated with the inhibitors mentioned above for 1 h, followed by control and 1m of LPA and S1P treatments for 4 h. Conditioned media were collected and resolved on a substrate gel. We found that LPL induction of MMP-2 activity was signifi-cantly blocked by SKF96365 but partially suppressed by PDTC and PD98059, implying that calcium mobilization, MEK/ERK, and NF-B signaling were involved. (Fig. 8). These results are consistent with the above-mentioned mRNA and protein results.
LPA and S1P induce MMP-dependent endothelial cell invasion. We further investigated whether enhancements of MMP by LPLs were responsible for the cell invasive activity. We tested the hypothesis that LPL treatments induce endothelial cell invasion by an in vitro Boyden chamber invasion assay. No significant cell invasion was observed under control con-ditions, whereas dramatic enhancement was seen in the pres-ence of LPLs. The enhancement effects occurred in a con-centration-dependent manner (Fig. 9A). The maximal effects were about 4- and 5-fold increases by S1P and LPA treat-ments, respectively.
To further demonstrate that LPL-induced EAhy926 cell invasion was due to the induction of MMPs, we used GM6001, a chemically synthesized MMP inhibitor in the invasion experiments. We found that LPL-induced EAhy926 cell invasion was significantly inhibited by GM6001 (Fig. 9B). The amounts of invading cells were also monitored by mi-croscopy (Fig. 9C). Together, these results suggested that the LPL-induced endothelial cell invasive capacity is MMP dependent.
GM6001 inhibited basal or LPL-induced endothelial cell adhe-sion to gelatin. Because endothelial cell invaadhe-sion of gelatin gel was enhanced by LPLs through up-regulating MMP-2 enzymatic activity, we further investigated whether these enhancement effects were due to increase of cell adhesion. We tested the hypothesis by an in vitro adhesion assay. Treatments with 1m LPA or S1P had limited effects on
Eahy926 cell adhesion to gelatin gel. However, significant suppression of cell adhesion was observed in both control and treated Eahy926 cells coincubated with 10m GM6001 (Fig. 10). These results indicated that LPA and S1P strongly enhanced endothelial cell invasion but had lim-ited effects on endothelial cell adhesion. On the other hand, GM6001, a broad MMP inhibitor, showed strong suppression effects on both endothelial cell invasion and adhesion. These results indicated that MMP-2 participates in both endothelial cell invasion of and adhesion to gelatin gel. In addition, the inhibitory effects of GM6001 on en-dothelial cell adhesion might also be responsible for the suppression of endothelial cell invasion induced by LPLs. FIG. 7. Enhancement of MMP-2 protein expression by LPA and S1P treatments are suppressed by the chemical inhibitors. EAhy926 cells were pretreated with control media alone, 15 ng/ml of PTx (Gi inhibitor), 10MPD98059 (MEK/ERK inhibitor), 100M of PDTC (NF-B inhibitor), and 10 MSKF96365 (calcium influx inhibitor) for 1 h. Inhibitor-treated cells were then treated with control medium alone or 1 M of LPA or S1P for 4 h. Total cell lysates were collected, and MMP-2 protein expression levels were monitored by Western blotting using a human monoclonal anti-MMP-2 antibody. A human-actin antibody was used as the load-ing control. The results were scanned, quantified, and shown in bar graph. Each bar of the histogram represents a minimum of three experiments, and data are presented as the mean ⫾ SD. These results were analyzed by two-way ANOVA. A level of P⬍ 0.01 was accepted as significant. **, Statistically different, compared with the level in control cells (P⬍ 0.01).
Discussion
Our current study provides evidence that LPA and S1P produced by some cancer cells or secreted by activated plate-lets might enhance MMP-2 expression in surrounding dothelial cells. The up-regulation of MMP-2 in human en-dothelial cells is directly correlated with its invasion potential. LPA levels in ovarian cancer patients are elevated in both plasma and ascites, suggesting that this bioactive lipid may contribute to early events that promote cancer cell dissemination. This idea was supported by the observation that LPA induces urokinase secretion by ovarian cancer cells (8, 46). MMP and urokinase-type plasminogen activator (uPA) systems are mutually related and are both involved in carcinoma progression through adjacent extracellular deg-radation. LPA and uPA are both present at elevated levels in ovarian cancer, and uPA has been reported to be linked to
malignant transformation in ovarian cells (47). A previous study (48) also showed that uPA can activate several MMPs. Our preliminary data showed that LPLs also induce uPA activity by casein zymography assays (data not shown). These results strongly suggest that LPLs play critical roles in endothelial cell invasion.
Previous work suggested that S1P induces MMPs and integrin-dependent HUVEC invasion and lumen formation (49). In this report, we present the molecular mechanism of LPL regulation of MMP-2 expression in EAhy926 cells. Fur-thermore, the concentration- and time-dependent activation of MMP-2 by LPLs is also consistent with a receptor-medi-ated mechanism. Cell migration and proteolysis are two es-sential processes during tumor invasion and metastasis, and MMPs are prime candidates for these activities. Furthermore, there is accumulating evidence that elevated expression of FIG. 8. Enhancement of MMP-2 gelatinolytic activity
by LPA and S1P treatments is partially suppressed by chemical inhibitors. A, EAhy926 cells were starved over-night in serum-free medium and pretreated with per-tussis toxin (a Gi blocker), PD98059 (a MEK blocker), U73122 (a PLC blocker), LY 294002 (a PI3K blocker), PDTC (an NF-B blocker), PP2 (an Src family kinase blocker), and SKF 96365 (a receptor-mediated calcium influx blocker) for 1 h and then treated with control and 1Mof LPA and S1P for 4 h. Conditioned media were collected and subjected to gelatin zymographic analysis. After staining with 0.25% Coomassie blue R 250, in-creases in gelatinolytic activity was detected. B, Similar experiments were repeated three times, and a represen-tative result is shown in the figure. These results were analyzed by two-way ANOVA. A level of P⬍ 0.05 was accepted as significant. *, Statistically different, com-pared with the level in control cells (P⬍ 0.05).
MMPs is observed in various cancer tissues (43, 50). It is now clearer that MMP substrates are not limited to the ECM but include other proteinases, proteinase inhibitors, clotting fac-tors, chemotactic molecules, latent growth facfac-tors, growth factor-binding proteins, cell surface receptors, and cell-cell adhesion molecules (39). MMPs are also known to solubilize cell surface and matrix-bound factors that can then act in an autocrine or paracrine manner to influence cellular proper-ties such as growth, apoptosis, and migration. In prostate cancer cells, LPA-induced ERK activation involves MMPs, which cause the release of active epithelial growth factor receptor ligands and may induce cancer cell proliferation (51). This is an example of MMP-mediated transactivation of cell surface receptors by proteolytically cleavage of plasma membrane-anchored proteins to produce catalytically active hormones. Moreover, it has also been shown that LPA pro-motes MMP activation and MMP-dependent invasion in ovarian cancer cells (52). These results suggest that the effects of LPLs on cell invasion might be partially mediated through enhancement of MMP-2 expression.
LPA and S1P bind to Edg receptors and activate multiple G proteins including Gi, Gq, and G12/13. These activated G ments, and the data are presented as the mean⫾SD. These results were analyzed by two-way ANOVA. A level of P⬍ 0.05 was accepted as significant. a, Significant differences between control and LPL treatment groups; b, significant differences between LPL treatment and inhibitor treatment groups. C, Amounts of invading cells were monitored by microscopy. a, Untreated control-1; b, S1P, 1M; c, S1P, 5M; d, untreated control-2; e, LPA, 1M; f, LPA, 5M; g, control pretreated with GM6001; h, LPA, 1Mpretreated with GM6001; and i, S1P, 1 M pretreated with GM6001. Similar experiments were repeated three times, and a representative result is shown in the figure. Bar, 300m.
FIG. 9. LPA and S1P increase invasion of EAhy926 cells, and the effect is blocked by the MMP inhibitor. EAhy926 cells were starved overnight in serum-free medium. A, Control and 0.5, 1, and 5Mof S1P and LPA treatments were loaded in the lower chamber as a chemotaxis inducer. Cells at 5⫻ 104/well were plated in the upper
chamber and allowed to migrate through 8-m porous filters coated with 2% gelatin for 4 h. Noninvading cells were removed, and the filter was fixed and stained with crystal violet. B, Suspended cells were pretreated with GM6001, a broad MMP inhibitor, for 1 h. GM6001 significantly suppressed endothelial cell invasion enhanced by LPLs. Each bar of the histogram represents a minimum of three
experi-FIG. 10. GM6001 blocked basal or LPL induced-endothelial cell ad-hesion to gelatin. EAhy926 cells were starved overnight in serum-free medium and then treated with control and 1Mof LPA and S1P as indicated for 4 h. Cells were trypsinized and then coincubated with media in the presence (open bars) or absence (solid bars) of 10M GM6001 in gelatin precoated 96-wells for 45 min. Adhered Eahy926 cells were washed and stained by crystal violet and quantified by OD595. Histograms represent quantification results of adherent Eahy926 cells. Similar experiments were repeated three times, and a representative result is shown in the figure. *, Statistically differ-ent, compared with the level in control cells (P⬍ 0.05).
proteins turn on downstream signaling events including MAPK phosphorylation, Ca2⫹mobilization, and cytoskele-ton reorganization (24). Herein, we report that the effect of LPA and S1P on endothelial cell invasion activity can be inhibited by MEK/ERK, NF-B, and calcium influx inhibi-tors (Fig. 8). MMP-13 induction in human osteoblastic cells and the IL-1- and TNF␣-induced MMP-1 secretion in hu-man pancreatic cell are involved in the MEK/ERK pathway (53). Additionally, calcium influx has been suggested to modulate expression of MMP-2 (54). Collectively, these re-sults suggest that the effects of LPA and S1P on endothelial cell MMP-2 expression are mediated by Edg receptors and multiple G protein-activated downstream effectors.
The enhancement effects of MMP-2 gelatinolytic activity by LPA and S1P were only partially suppressed by PD98059, PDTC, and SKF96365. This was possibly due to the fact that the enzymatic activity of MMP-2 was regulated by a com-plicated and delicate mechanism, including posttranscrip-tion, secretion regulaposttranscrip-tion, proteolytic activaposttranscrip-tion, and proteo-lytic inhibition. Therefore, gelatinoproteo-lytic activities do not reveal complete inhibition effects as those on mRNA levels. Another possible reason is that the quantification of gelatin zymographic analyses is not as precise.
MMPs are up-regulated in many types of human cancer, and their expression is often associated with poor survival (55). Several studies have reported the up-regulation of MMP-2 in cancer cells, including breast, colon, ovarian, pan-creatic, and liver cancers (37, 56). Whereas some MMPs (for example, MMP-1, MMP-2, and MMP-7) are expressed by cancer cells, other MMPs (for example, MMP-2 and MMP-9) are also synthesized by tumor stromal cells, including fibro-blasts, myofibrofibro-blasts, inflammatory cells, and endothelial cells. Animal studies have also shown that in prostate can-cers, only cells expressing high levels of MMP-2 and MMP-9 have the ability to metastasize (57). Metastasis is a major cause of death among cancer patients, which requires several sequential events, such as changes in cell-ECM interactions, the disconnection of intercellular adhesion and separation of single cells from solid tumor tissue, degradation of the ECM, and the movement of tumor cells into the ECM. Relatively benign cells acquire malignant properties when MMP activ-ity is increased or TIMP activactiv-ity diminished (58, 59). This is consistent with our findings that the enhancement of MMP-2 is related to the invasive properties of endothelial cells, sug-gesting that LPLs might contribute to cancer invasion by enhancement of MMP-2 production from an endothelial or-igin. MMP-2-deficient mice showed slightly smaller body weight but displayed significant reduction in both the pri-mary tumor mass and metastatic spread (60). MMP-2 (and other MMPs as well) therefore are tempting targets for ther-apeutic intervention strategies in diseases such as cancer. The phenotypes of some MMP-deficient mice suggest, however, that this approach may give rise to severe side effects. MMP inhibitor therapy has yielded spectacular results in animal models, but unfortunately not in humans (61).
During angiogenesis, endothelial cells experience loosen-ing of the matrix and intercellular adhesion; degradation of the subendothelial matrix; and the migration, proliferation, and formation of new tubes (62– 64). MMPs play an impor-tant role in these steps by degrading the ECM so that cells can
move across tissues into nearby stroma (65). S1P-induced invasion may facilitate the guidance of endothelial cells cre-ating an initial sprouting and may also contribute to the ability of endothelial cells (49). These results are consistent with our current observation that LPLs might be important regulators for endothelial cells by modulating MMP-2 expression.
The broad-spectrum MMP inhibitor, GM 6001 (galardin, Ilomastat), is a hydroxamic acid originally synthesized as an inhibitor of human skin collagenase, and has been shown to block MMP-1, MMP-2, MMP-3, and MMP-9 (66). In our study, we showed that LPL-induced human endothelial cells invasion was blocked by this inhibitor. Using trypan blue staining, which distinguishes viable from nonviable cells, we confirmed that GM6001’s inhibition of endothelial invasion was not due to the reagent having a lethal effect at the indicated concentration (data not shown). Koike et al. (67) group observed that aged human microvascular endothelial cell lines exhibited poor formation of tubular, capillary-like structures in vitro, and diminished expression of active MMP-2 and also showed that GM6001 decreased tubulogen-esis in endothelial cells. Another study showed that GM6001 blocked LPA-induced epithelial growth factor receptor transactivation in rat-1 cells (68). Moreover, Bayless’ group used GM6001 to inhibit S1P-induced MMP-2 activation (49). Those results are consistent with our current observations. We found that both LPA and S1P up-regulate MMP-2 protein expression levels as little as 1 h. Previous studies indicated that some proinflammatory cytokines such as TNF-induced MMP-9 expression through modulating inhibitory B␣ phosphorylation, ubiquitination, and degradation (69). These results suggest that LPLs might induce MMP-2 ex-pression through these processes and result in rapid changes of MMP-2 protein expression.
In summary, our study clearly indicates that LPLs increase MMP-2 mRNA, protein, and enzymatic activities in concen-tration- and time-dependent manners, and these enhance-ments are consistent with a receptor-mediated mechanism. The induction is mediated by MEK/ERK-, NF-B-, and cal-cium influx-dependent signaling pathways. Furthermore, the enhancement effects of LPA and S1P on MMP-2 expres-sion in EAhy926 cells are responsible for the induction of the invasive activity. These results suggest that LPA and S1P might be important regulators of the interaction of cells and the extracellular matrix. Both lipids might play critical roles in regulating angiogenesis and cancer metastasis through the regulation of MMP-2.
Acknowledgments
We thank Dr. S. M. Lee (Chung Shang Hospital) for providing um-bilical cords for endothelial cell preparation and Dr. Cora-Jean S. Edgell (Department of Pathology, University of North Carolina, Chapel Hill, NC) for providing endothelial cell hybridoma Eahy926 cells. We thank Ms. Ting-En Su for technical assistance.
Received December 22, 2004. Accepted April 26, 2005.
Address all correspondence and requests for reprints to: Hsinyu Lee, De-partment of Life Science and Institute of Zoology, National Taiwan University, Taipei, Taiwan 10617, Republic of China. E-mail: [email protected].
This work was supported by National Science Council Grants 90-2316-B-002-019, 91-2311-B-002-071, and 92-2311-B-002-095 (to H.L.),
Tai-wan, and 91-B-FA09-2-4 supported by Ministry of Education Program for promoting Academic Excellence of Universities Grant (to H.L.), Taiwan.
References
1. Gerrard JM, Robinson P 1989 Identification of the molecular species of ly-sophosphatidic acid in platelets stimulated by thrombin. Biochim Biophys Acta 1001:282–288
2. Gaits F, Fourcade O, Le Belle F, Gueguen G, Gaige B, Gassama-Diagne A,
Fauvel J, Salles J-P, Mauco G, Simon M-F, Chap H1997 Lysophosphatidic acid as a phospholipid mediator: pathways of synthesis. FEBS Lett 410:54 –58 3. Fourcade O, Simon MF, Viode C, Rugani N, Leballe F, Ragab A, Fournie B,
Sarda L, Chap H1995 Secretory phospholipase A2 generates the novel lipid mediator lysophosphatidic acid in membrane microvesicles shed from acti-vated cells. Cell 80:919 –927
4. Lochter A, Galosy S, Muschler J, Freedman N, Werb Z, Bissell MJ 1997 Matrix metalloproteinase stromelysin-1 triggers a cascade of molecular alterations that leads to stable epithelial-to-mesenchymal conversion and a premalignant phenotype in mammary epithelial cells. J Cell Biol 139:1861–1872
5. Lee HY, Liao JJ, Graeler M, Huang MC, Goetzl EJ 2002 Lysophospholipid regulation of mononuclear phagocytes. Biochim Biophys Acta 1582:175–177 6. Xu Y, Shen Z, Wiper DW, Wu M, Morton RE, Elson P, Kennedy AW, Belinson
J, Markman M, Casey G1998 Lysophosphatidic acid as a potential biomarker for ovarian and other gynecologic cancers. JAMA 280:719 –723
7. Shen Z, Belinson J, Morton RE, Xu Y 1998 Phorbol 12-myristate 13 acetate stimulates lysophosphatidic acid secretion from ovarian and cervical cancer cells but not from breast or leukemia cells. Gynecol Oncol 71:364 –368 8. Pustilnik TB, Estrella V, Wiener JR, Mao M, Eder A, Watt MV, Bast Jr RC,
Mills GB1999 Lysophosphatidic acid induces urokinase secretion by ovarine cancer cells. Clin Cancer Res 5:3704 –3710
9. An SZ, Bleu T, Zheng YH, Goetzl EJ 1998 Recombinant human G protein-coupled lysophosphatidic acid receptors mediate intracellular calcium mobi-lization. Mol Pharmacol 54:880 – 888
10. Spiegel S, Merrill Jr AH 1996 Sphingolipid metabolism and cell growth regulation. FASEB J 10:1388 –1397
11. Xie Y, Gibbs TC, Meier KE 2002 Lysophosphatidic acid as an autocrine and paracrine mediator. Biochim Biophys Acta 1582:270 –281
12. Moolenaar WH 1999 Bioactive lysophospholipids and their G protein-coupled receptors. Exp Cell Res 253:230 –238
13. Noguchi K, Ishii S, Shimizu T 2003 Identification of p2y9/GPR23 as a novel G protein-couple receptor for lysophosphatidic acid, structurally distant from the Edg family. J Biol Chem 278:25600 –25606
14. Ishii I, Fukushima N, Ye X, Chun J 2004 Lysophospholipid receptors: sig-naling and biology. Ann Rev Biochem 73:321–354
15. Moolenaar WH, Kranenburg O, Postma FR, Zondag G 1997 Lysophospha-tidic acid: G protein signaling and cellular responses. Curr Opin Cell Biol 9:168 –173
16. Tigyi G, Fischer DJ, Sebok A, Marshall F, Dyer DL, Miledi R 1996 Lyso-phosphatidic acid-induced neurite retraction in PC12 cells: neurite-protective effects of cyclic AMP signaling. J Neurochem 66:549 –558
17. Moolenaar WH 1995 Lysophosphatidic acid signalling. Curr Opin Cell Biol 7:203–210
18. Stahle M, Veit C, Bachfischer U, Schierling K, Skripczynski B, Hall A,
Gierschik P, Giehl K2003 Mechanisms in LPA-induced tumor cell migration: critical role of phosphorylated ERK. J Cell Sci 15:3835–3846
19. Seewald S, Schmitz U, Seul C, Ko Y, Sachinidis A, Vetter H 1999 Lysophos-phatidic acid stimulates protein kinase C isoforms␣, , ␦ and in a pertussis toxin sensitive pathway in vascular smooth muscle cells. Am J Hypertens 12:532–537
20. Yatomi Y, Ozaki Y, Ohmori T, Igarashi Y 2001 Sphingosine 1-phosphate: synthesis and release. Prostaglandins 64:107–122
21. Pages C, Simon M, Valet P, Saulnier-Blache JS 2001 Lysophosphatidic acid synthesis and release. Prostaglandins Other Lipid Mediat 64:1–10 22. Fang X, Gaudette D, Furui T, Mao M, Estrella V, Eder A, Pustilnik T,
Sasagawa T, Lapushin R, Yu S, Jaffe RB, Wiener JR, Erickson JR, Mills GB
2000 Lysophospholipid growth factors in the initiation, progression, metas-tases, and management of ovarian cancer. Ann NY Acad Sci 905:188 –208 23. Bazzoni G, Dejana E, Lampugnani MG 1999 Endothelial adhesion molecules
in the development of the vascular tree: the garden of forking paths. Curr Opin Cell Biol 11:573–581
24. Lee H, Geotzl EJ, An S 2000 Lysophosphatidic acid and sphingosine 1-phos-phate stimulate endothelial cell wound healing. Am J Physiol 47:C612–C617 25. Spiegel S, Milstien S 2003 Sphingosine-1-phosphate: an enigmatic signaling
lipid. Nat Rev Mol Cell Biol 4:397– 407
26. Nagase H, Woessner Jr JF 1999 Matrix metalloproteinases. J Biol Chem 274: 21491–21494
27. Ellis V, Murphy G 2001 Cellular strategies for proteolytic targeting during migration and invasion. FEBS Lett 506:1–5
28. Abecassis I, Olofsson B, Schmid M, Zalcman G, Karniguian A 2003A induces MMP-9 expression at CD44 lamellipodial focal complexes and pro-motes HMEC-1 cell invasion. Exp Cell Res 291:363–376
29. Strongin AY, Collier I, Bannikov G, Marmer BL, Grant GA, Goldberg GI 1995 Mechanism of cell surface activation of 72-kDa type IV collagenase. J Biol Chem 270:5331–5338
30. Murphy G, Gavrilovic J 1999 Proteolysis and cell migration: creating a path? Curr Opin Cell Biol 11:614 – 621
31. Hernandez-Barrantes S, Toth M, Bernardo MM, Yurkova M, Gervasi DC,
Raz Y, Sang QA, Fridman R2000 Binding of active (57 kDa) membrane type 1-matrix metalloproteinase (MT1-MMP) to tissue inhibitor of metalloprotein-ase (TIMP)-2 regulates MT1-MMP processing and pro-MMP-2 activation. J Biol Chem 275:12080 –12089
32. Hotary K, Allen E, Punturieri A, Yana I, Weiss SJ 2000 Regulation of cell invasion and morphogenesis in a three dimensional type collagen I matrix by membrane type matrix metalloproteinases 1, 2 and 3. J Cell Biol 149:1309 –1323 33. Seiki M 2002 The cell surface: the stage for matrix metalloproteinase regulation
of migration. Curr Opin Cell Biol 14:624 – 632
34. Zhao H, Bernardo MM, Osenkowski P, Sohail A, Pei D, Nagase H,
Kashi-wagi M, Soloway PD, DeClerck YA, Fridman R2004 Differential inhibition of membrane type 3 (MT3)-matrix metalloproteinase (MMP) and MT1-MMP by tissue inhibitor of metalloproteinase (TIMP)-2 and TIMP-3 regulates pro-MMP-2 activation. J Biol Chem 279:8592– 8601
35. Zhang J, Bai S, Tanase C, Nagase H, Sarras Jr MP 2003 The expression of tissue inhibitor of metalloproteinase 2 (TIMP-2) is required for normal development of zebrafish embryos. Dev Genes Evol 213:382–389
36. Zhang J, Bai S, Zhang X, Nagase H, Sarras Jr MP 2003 The expression of gelatinase A (MMP-2) is required for normal development of zebrafish em-bryos. Dev Genes Evol 213:456 – 463
37. Sternlicht MD, Werb Z 2001 How matrix metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol 17:463–516
38. Greene J, Wang M, Liu YE, Raymond LA, Rosen C, Shi YE 1996 Molecular cloning and characterization of human tissue inhibitor of metalloproteinase 4. J Biol Chem 271:30375–30380
39. Friedl P, Wolf K 2003 Tumor-cell invasion and migration: diversity and escape mechanisms. Nat Rev Cancer 3:362–374
40. Porter JF, Shen S, Denhardt DT 2004 Tissue inhibitor of metalloproteinase-1 stimulates proliferation of human cancer cells by inhibiting a metalloprotein-ase. Br J Cancer 90:463– 470
41. Bergers G, Brekken R, McMahon G, Vu TH, Itoh T 2000 Matrix metallo-proteinase-9 triggers the angiogenic switch during carcinogenesis. Nat Cell Biol 2:737–744
42. Lusis AJ 2000 Atherosclerosis. Nature 407:233–241
43. Edgell C, McDonald C, Graham J 1983 Permanent cell line expressing human factor VIII-related antigen established by hybridisation. Proc Natl Acad Sci USA 80:3734 –3737
44. Schnapp LM, Hatch N, Ramos DM, Klimanskaya IV, Sheppard D, Pytela R 1995 The human integrin␣81 functions as a receptor for tenascin, fibronectin, and vitronectin. J Biol Chem 270:23196 –23202
45. Szklarczyk A, Lapinska J, Rylski M, McKay RD, Kaczmarek L 2002 Matrix metalloproteinase-9 undergoes expression and activation during dendritic remodeling in adult hippocampus. J Neurosci 22:920 –930
46. Huang MC, Lee HY, Yeh CC, Kong Y, Zaloudek CJ, Goetzl EJ 2004 Induction of protein growth factor systems in the ovaries of transgenic mice overex-pressing human type 2 lysophosphatidic acid G protein-coupled receptor (LPA2). Oncogene 23:122–129
47. Sier CF, Stephens R, Bizik J, Mariani A, Bassan M, Pedersen N, Frigerio L,
Ferrari A, Dano K, Brunner N, Blasi F1998 The level of urokinase-type plasminogen activator receptor is increased in serum of ovarian cancer pa-tients. Cancer Res 58:1843–1849
48. Rabbani SA, Mazar AP 2001 The role of the plasminogen activation system in angiogenesis and metastasis. Surg Oncol Clin North Am 10:393– 415 49. Bayless KJ, Davis GE 2003 Sphingosine-1-phosphate markedly induces matrix
metalloproteinase and integrin-dependent human endothelial cell invasion and lumen formation in three-dimensional collagen and fibrin matrices. Bio-chem Biophys Res Commun 312:903–913
50. Menashi S, Serova M, Ma L, Vignot S, Mourah S, Calvo F 2003 Regulation of extracellular matrix metalloproteinase inducer and matrix metalloprotein-ase expression by amphiregulin in transformed human breast epithelial cells. Cancer Res 15:7575–7580
51. Daaka Y 2002 Mitogenic action of LPA in prostate. Biochim Biophys Acta 1582:265–269
52. Fishman DA, Liu Y, Ellerbroek SM, Stack MS 2001 Lysophosphatidic acid promotes matrix metalloproteinase (MMP) activation and MMP-dependent invasion in ovarian cancer cells. Cancer Res 61:3194 –3199
53. Yang CM, Chien CS, Yao CC, Hsiao LD, Huang YC, Wu CB 2004 Mechanical strain induces collagenases-3 (MMP-13) expression in MC3T3–E1 osteoblastic cells. J Biol Chem 279:22158 –22165
54. Kohn EC, Jacobs W, Kim YS, Alessandro R, Stetler-Stevenson WG, Liotta LA 1994 Calcium influx modulates expression of matrix metalloproteinase-2 (72-kDa type IV collagenase, gelatinase A). J Biol Chem 269:21505–21511 55. Zucker S, Lysik RM, Zarrabi MH, Moll U 1993 M(r) 92, type IV collagenase
is increased in plasma of patients with colon cancer and breast cancer. Cancer Res 53:140 –146
56. Chang C, Werb Z 2001 The many faces of metalloproteases: cell growth, invasion, angiogenesis and metastasis. Trends Cell Biol 11:S37–S43 57. Lokeshwar BL 1999 MMP inhibition in prostate cancer. Ann NY Acad Sci
878:271–289
58. Johnsen M, Lund LR, Ru`mer J, Almholt K, Danu` K 1998 Cancer invasion and tissue remodeling: common themes in proteolytic matrix degradation. Curr Opin Cell Biol 10:667– 671
59. Stamenkovic I 2000 Matrix metalloproteinases in tumor invasion and metas-tasis. Seminars Cancer Biol 10:415– 433
60. Itoh T, Tanioka M, Yoshida H, Yoshioka T, Nishimoto H, Itohara S 1998 Reduced angiogenesis and tumor progression in gelatinase A-deficient mice. Cancer Res 58:1048 –1051
61. Coussens LM, Fingleton B, Matrisian LM 2002 Matrix metalloproteinase inhibitors and cancer: trials and tribulations. Science 295:2387–2392 62. Risau W 1997 Mechanisms of angiogenesis. Nature 386:671– 674
63. Griffioen AW, Molema G 2000 Angiogenesis: potentials for pharmacologic intervention in the treatment of cancer, cardiovascular diseases, and chronic inflammation. Pharmacol Rev 52:237–268
64. Langlois S, Gingras D, Beliveau R 2004 Membrane type 1-matrix
metallo-proteinase (MT1-MMP) cooperates with sphingosine 1-phosphate to induce endothelial cell migration and morphogenic differentiation. Blood 103:3020 – 3028
65. Seo DW, Li H, Guedez L, Wingfield PT, Diaz T, Salloum R, Wei BY,
Stetler-Stevenson WG2003 TIMP-2 mediated inhibition of angiogenesis: an MMP-independent mechanism. Cell 114:171–180
66. Agren MS, Mirastschijski U, Karlsmark T, Saarialho-Kere UK 2001 Topical synthetic inhibitor of matrix metalloproteinases delays epidermal regeneration of human wounds. Exp Dermatol 10:337–348
67. Koike T, Vernon RB, Gooden MD, Sadoun E, Reed MJJ, Gerontol A 2003 Inhibited angiogenesis in aging: a role for TIMP-2. Biol Sci Med Sci 58:B798 – B805
68. Santiskulvong C, Rozengurt E 2003 Galardin (GM 6001), a broad-spectrum ma-trix metalloproteinase inhibitor, blocks bombesin- and LPA-induced EGF receptor transactivation and DNA synthesis in rat-1 cells Exp Cell Res 290:437–446 69. Takada Y, Fang X, Jamaluddin MS, Boyd DD, Aggarwal BB 2004 Genetic
deletion of glycogen synthase kinase-3 abrogates activation of IB␣ kinase, JNK, Akt, and p44/p42 MAPK but potentiates apoptosis induced by tumor necrosis factor. J Biol Chem 79:39541–39554
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving the endocrine community.