Chemical Composition and Bioactivities of the Marine Alga
Isochrysis galbana CCMP 1324 from Taiwan
Chi-Cheng Yua, Hsiao-Wei Chenb, Mao-Jing Chenb, Yu-Ching Changb, Shih-Chang Chien c, Yuen-Hsiung Kuod, Feng-Ling Yange, Shih-Hsiung Wue, Jie Chenf, and Louis Kuop-Ping Chaof*
a Greenlink Biotech Inc. 12 F, No. 182, Sec. 4, Chengde Rd., Shilin Dist., Taipei, Taiwan 111
bChemistry and Environment Labs., Taiwan Power Research Institute. 84, Da-ani Rd., Shulin, Taipei country, Taiwan 238
cSchool of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University, Taichung, Taiwan d Tsuzuki Institute for Traditional Medicine, College of Pharmacy, China Medical University, Taichung, Taiwan 404
e
Institute of Biological Chemistry, Academia Sinica, Taipei 115,
f Department of Cosmeceutics, China Medical University, Taichung, Taiwan 404
*To whom correspondence should be addressed. [email protected]; [email protected]
Received: Aug 18th, 2010; Accepted:
The present study investigated the chemical composition of Isochrysis galbana, a species of marine microalgae which is frequently and widely used as a feedstock in aquaculture. The I. galbana alga contained hot-water extractable polysaccharides (11.17%); after hydrolysis the carbohydrate components were treated with trimethylsilane for derivatization and subsequent GC-MS analysis. The mono-sugar compositions of I. galbana were 2.07% fucose, 2.50% rhamnose, 2.72% arabinose, 8.49% xylose, 15.7% mannose, 32.73% galactose and 35.79% glucose. Using the murine macrophage cell line J774A.1, we found that polysaccharides of I. galbana were able to induce prointerleukin-1β (pro-IL-1β) protein expression. Furthermore, the composition of I. galbana was determined to consist of 8.58% ash, 38.16% extractable by 95% alcohol and 21.75% extractable by alcohol/hexane (1/1). Five kinds of chlorophyll and one sterol were separated from the alcoholic extractives of I. galbana, including pheophorbide-a, ethyl pheophorbide-a, 10S-10-hydroxypheophytin-a, 10R-10-10S-10-hydroxypheophytin-a, (132-R)-pheophyin-a, and brassicasterol. In addition, the major soluble components of alcohol/hexane were found to be 9-octadecenoic acid (E) (38.41%), hexadecanoic acid (23.34%), tetradecanoic acid (15.70%), and octadecanoic acid (7.16%) – but only a few polyunsaturated fatty acids such as 9,12,15-octadecatrienoic acid (1.95%), 9,12-octadecadienoic acid (Z,Z) (3.44%), and docosahexaenoic acid (0.21%). This is the first occasion that polysaccharides from I. galbana have been demonstrated to exert immunomodulatory properties by the induction of IL-1 within macrophages.
Keywords: Isochrysis galbana, extractives, chemical compositions, polysaccharides, bioactivity, pro-IL-1β.
Marine microalgal biomasses play an important role as primary producers in the animal food chain. Isochrysis
galbana, a golden-brown flagellate marine microalga, is
widely used as an aquaculture feed for young fish and in bivalve hatcheries owing to its being rich in PUFA [1]. Many studies have focused on the relationship between fatty acids and algal growth [2-4]. In addition, because of the high content of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA), I. galbana is recognized for its beneficial effects on human health, and is considered to be a good substitute for fish oil in the human diet. However, only a few studies have shown the detailed chemical composition of I. galbana. The purpose of the present study was to analyze the chemical composition of
I. galbana, including: hexane/alcohol extracts; alcohol
extracts; total carbon, hydrogen, oxygen, nitrogen and sulfur; and total polysaccharides.
First we analyzed the ratio of carbon, hydrogen, oxygen, nitrogen, sulfur and ash of I. galbana cultured in the laboratory. The results showed that I. galbana contains: C (56.35%), H (7.55%), O (20.29%), N (6.25%), S (0.97%) and ash (8.59%), based on dry weight. I. galbana also has a high content of hexane/alcohol (1/1) extracts, as well as extracts of alcohol only which amount to 21.75% and 38.16%.
NPC
Natural Product Communications
Vol. 5
2010
No. 0
1 - 4
2 Natural Product Communications Vol. 5 (0) 2010 Yu et al.
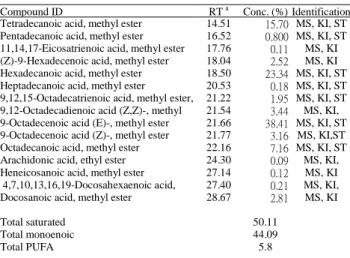
Sixteen components of fatty acids from I. galbana are identified and presented in Table 1, where all compounds are listed in order of their elution from the DB-5HT column. The major component of fatty acids is 9-octadecenoic acid (E) (38.41% of total fatty acids), followed by hexadecanoic acid (23.34%), tetradecanoic acid (15.70%), octadecanoic acid (7.16%), and 9-octadecenoic acid (Z) (3.16%). The compositions of fatty acids from I. galbana in this study are not similar to an earlier study, which showed higher DHA (22:6n-3) (7.91%) [5]. Also, some differences were found from the study by Lin et al., who determined DHA to be a major fatty acid at every growth phase [4]. It is interesting that we found a few fatty acids of odd carbons such as pentadecanoic acid, heptadecanoic acid, and heneicosanoic acid, which had not been previously found [4]. This could be a result of different growth conditions.
Table 1: Chemical composition of L. galbana, alcohol/hexane soluble.
Compound ID RT a Conc. (%) Identification
b
Tetradecanoic acid, methyl ester 14.51 15.70 MS, KI, ST
Pentadecanoic acid, methyl ester 16.52 0.800 MS, KI, ST
11,14,17-Eicosatrienoic acid, methyl ester 17.76 0.11 MS, KI (Z)-9-Hexadecenoic acid, methyl ester 18.04 2.52 MS, KI Hexadecanoic acid, methyl ester 18.50 23.34 MS, KI, ST
Heptadecanoic acid, methyl ester 20.53 0.18 MS, KI, ST
9,12,15-Octadecatrienoic acid, methyl ester, 21.22 1.95 MS, KI, ST
9,12-Octadecadienoic acid (Z,Z)-, methyl 21.54 3.44 MS, KI, 9-Octadecenoic acid (E)-, methyl ester 21.66 38.41 MS, KI, ST
9-Octadecenoic acid (Z)-, methyl ester 21.77 3.16 MS, KI,ST
Octadecanoic acid, methyl ester 22.16 7.16 MS, KI, ST
Arachidonic acid, ethyl ester 24.30 0.09 MS, KI, Heneicosanoic acid, methyl ester 27.14 0.12 MS, KI 4,7,10,13,16,19-Docosahexaenoic acid, 27.40 0.21 MS, KI, Docosanoic acid, methyl ester 28.67 2.81 MS, KI
Total saturated 50.11
Total monoenoic 44.09
Total PUFA 5.8
a Retention time on a DB-5 HT column with reference to n-alkanes. b MS, NIST and Wiley library spectra and the literature; RI, Retention
index; ST, authentic standard compounds.
In general, chlorophyll compounds are used for estimating the biomass of microalgae and can be used to measure growth. Few existing studies have focused on the variety of pigments from I. galbana. Herein we have separated and identified the pigments from 1.65 g of alcoholic extracts of I. galbana. As shown in Figure 1, I.
galbana contains at least five kinds of chlorophyll
compounds. They include (132-R)-pheophytin-a (1.5 mg; RT = 8.03 min), pheophorbide-a (1 mg; RT = 15.82 min), 10R-10-hydroxypheophytin-a (3 mg; RT = 22.54 min), 10S-10-hydroxypheophytin-a (1 mg; RT = 23.51 min), and ethyl pheophorbide-a (4 mg; RT = 34.92 min). We also found brassicasterol (2 mg; RT = 15.10 min). In 1981, Volkman et al. [6] were the first to report that 24-methyl-22-dehydrocholesterol was the dominant sterol from Isochrysis galbana [6]. Our experimental results were similar, in that brassicasterol was found to be the principal sterol from I. galbana. Park et al. (2002) [7]
demonstrated that autotrophically grown I. galbana contains three major sterols (24-oxocholesterol acetate, ergost-5-en-3-ol, and cholest-5-en-24-1,3-(acetyloxy)-,3-ol), with 24-methylcholesta-5,22-dien-3-ol as a minor sterol [7]. Meanwhile they found that the total sterol content will clearly decrease during dark culture, with such decreases being particularly evident in two major sterols, 24-oxocholesterol acetate and ergost-5-en-3-ol.
Figure 1. The chemical structure of alcoholic extracts from I. galbana.of
(1). Pheophorbide-a (2). Ethyl pheophorbide-a (3). (132
-R)-pheophyin-a (4). 10R-10-Hydroxypheophytin-a (5). 10S-10-Hydroxypheophytin-a (6). Brassicasterol
Our study shows that the mono-sugar compositions of Isochrysis polysaccharide (IP) from I. galbana were 2.07% fucose, 2.50% rhamnose, 2.72% arabinose, 8.49% xylose, 15.7% mannose, 32.73% galactose and 35.79% glucose. Furthermore, no cytotoxic effect was observed after J7774A.1 cells were treated with various concentrations of IP for a period of 24 h, as measured by MTT assay (Figure 2).
Figure 2: Effect of Isochrysis polysaccharide (IP) on cell viability. J774A.1
macrophages (5 × 103/well) were treated with IP or DMSO
(control) for 24 h, followed by incubating with MTT reagent The absorbance (A550-A690) was measured by
spectrophotometry. Data were expressed as mean ± SE from three separate experiments
An IP-induced IL-1 precursor, pro-IL-1, was detected in the whole cell lysates after I. galbana alga (IP) stimulation by means of Western-blotting analysis. In this time course study, the expression of pro-IL-1 protein within IP-stimulated cells was detected at 6 h
post-Chemical Composition and Bioactivities of the Isochrysis galbana Natural Product Communications Vol. 5 (0) 2010 3
stimulation. In addition, the expression of pro-IL-1 increased with increasing IP concentrations in a dosage-dependent manner (Figure 3). Such results demonstrate that IP stimulates pro-IL-1 expression, a result that would appear to be similar to the ability of polysaccharides isolated from algae of Rhizoclonium riparium or
Chlorella pyrenoidosa to stimulate pro-IL-1 expression
within murine macrophages [8-9]. Fabregas et al. (1999) found that endocellular extracts from I. galbana could clearly inhibit viral hemorrhagic septicemia virus (VHSV) replication at a dose of 20 µg/mL, and that I. galbana contained sulfated soluble exopolysaccharides [10].
Figure 3: IP induces pro-IL-1 protein expression in J774A.1 cells. Dose
response of Western-blotting analyses of pro-IL-1 protein expression in IP-treated cells: cells were treated with various concentrations of IP as indicated (0 to 800 µg/mL) for 6h, and whole cell lysates were analyzed by Western blotting (LPS as positive control). Pro-IL-1 and actin (as an internal control) are indicated by arrows on the right.
Experimental
Algal culture and collection:I. galbana alga used in this
study was obtained from the Tungkang Biotechnology Research Center, Fisheries Research Institute, Republic of China. PurifiedI. galbana CCMP 1324 (0.9 L, 680 nm,
OD 1.10) was inoculated and cultured in Walne’s medium [9]. This included nutrient solution, vitamin solution and trace metal solution in a 10 L PET tank containing 8.1 L seawater autoclaved at 120 °C for 20 min, to which was added 9.0 mL of nutrient solution and 0.9 mL of vitamin solution. The culture medium was agitated gently by bubbling air with a flow rate of 4.7 L/min and a culture time of 7 d. Continuous illumination at an irradiance of 5900 lx was provided by fluorescent lamps. The culture medium was then centrifuged (Himac CR22-GII, Hitachi, Japan) continuously at 12000 rpm, 25 ºC. After lyophilization the yields of the alga were 0.1 g/L (dry weight) culture medium.
Total C、、、、H、、、、O、、、、N、、、、S : Total carbon, hydrogen,
oxygen, nitrogen and sulfur contents were determined by CHN elemental analysis. Freeze-dried samples (15 g each) were combusted in a 2400 CHN/O elemental analyzer (Perkin-Elmer, Waltham MA, USA) [11].
Extraction and compositions of hexane/alcohol extracts of I. galbana: Ten g of the sample was extracted in a
Soxhlet apparatus with n-hexane:alcohol (95% v/v) (50:50) for 48 h. The solution was rotary-evaporated at 65
ºC to provide hydroalcoholic extracts (HAE) of 2.175 g. HAE (0.5 g) was tested for further trimethylsilylation with Sylon HTP (HMDS/TMCS/pyridine, 3:1:9) trimethylsilylation reagent (Supelco, Bellefonte PA, USA). The final trimethylsilylated (TMS) derivatives were kept in n-hexane for gas chromatography–mass spectrometry (GC-MS) analysis. A Hewlett-Packard HP 6890 gas chromatograph equipped with a DB-5HT fused silica capillary column (30 m x 0.25 mm x 0.25 µm film thickness; Agilent Technologies, Santa Clara CA, USA) and a FID detector was used for quantitative determination of the components. Oven temperature was programmed as follows: 100 ºC for 2 min, rising to 275 ºC at 5 ºC/min; injector temperature, 270 ºC; carrier gas, He with a flow rate of 1 mL/min; detector temperature, 250 ºC; split ratio 50.1:1. One µL sample was injected. Identification of the oil components was based on their retention indices and MS results. The GC analysis parameters listed above and the MS were obtained (full scan mode; scan time, 0.3 s; mass range, MHz 30–500) in the electron impact (EI) mode at 70 eV.
Extraction, purification and
identification
of alcoholic extracts of I. galbana: Dry alga (5 g) was treated withethanol (95% v/v for 10 d, repeated 3 times) at room temperature; then the extract was concentrated to provide alcoholic extract (AE). AE (1.65 g) was applied on a silica gel column (Si 60) for separation. Then it was eluted with acetone/n-hexane to give 43 subfractions. Each eluted volume of the fraction was 150 ml. The chlorophyll compounds were purified by preparative HPLC (KNAUER RI detector 2400, pump 100; KNAUER, Germany) on a Merck (Germany) Hibar Fertigsaule RT column Si 60 (25 cm length, 1 cm i.d., 5.0
µm). The separation conditions were as follows: flow rate
4 mL/min; mobile phase, acetone/hexane = 1/9. The sterol was separated by a Phenomenex Luna silica (2) column (25 cm length, 1 cm i.d., 5.0 µm) under the following conditions: flow rate 4 mL/min; mobile phase, acetone/hexane = 1/15. The structures of the compounds were confirmed by comparison of physical and spectral data (including optical rotation, EI-MS, 1H-NMR) with previously reported values.
Extraction of the polysaccharides from I. galbana: Five
g dry alga were extracted by hexane/alcohol, ground into a fine powder, and then suspended in 100 mL of distilled water. After autoclaving at 121 °C for 30 min, the extracts were filtered by a 0.2 µm membrane. The extracts were then vacuum-concentrated at 50 °C, giving a final volume of 30 mL to which five volumes of 95% ethanol was added slowly at 4 °C. Then the mixture was centrifuged to produce precipitates of ca. 1.910 g (d/w). Sixty mg of the precipitates were treated further with 3 mg proteinase K for removal of the peptide part, and
4 Natural Product Communications Vol. 5 (0) 2010 Yu et al.
dialyzed against H2O (Spectra/Por membrane, molecular
weight cutoff 1000 Da), resulting in 17.5 mg polysaccharide of I. galbana (IP).
Sugar compositions analysis: Sugar composition was
determined by GC-MS. Polysaccharide content of I.
galbana was determined by methanolysis with 0.5 M
methanolic HCl at 80 °C for 16 h and trimethylsilylation with Sylon HTP. The final trimethylsilylated (TMS) derivatives were kept in n-hexane for GC-MS analysis [12]. Carbohydrate analysis was done with inositol as the internal standard; its integrated peak area was used to establish the relative amounts of the constituents. The compounds were identified by comparing their mass spectrometric fragmentation patterns with those of authentic standards, and the quantity of compounds was obtained by integrating the peak area of the spectrograms.
Microculture tetrazolium (MTT) assay for cell viability:
J774A.1 macrophages were seeded in 96-well plates at a density of 5 × 103 cells/well. Cells were incubated with IP for 24 h. Cell viability was determined using colorimetric MTT assays.
Cell cultures: Murine J774A.1 macrophages were
obtained from the American Type Culture Collection (ATCC) (Rockville MD, USA). All cells were propagated
in RPMI-1640 medium supplemented with 10% heat-inactivated FCS and 2 mM L-glutamine (Life Technologies, Carlsbad CA, USA), and cultured in a 37
°C, 5% CO2 incubator [8,9].
Western Blotting: Whole cell lysates were separated by
12% SDS-PAGE and electrotransferred to a PVDF membrane. The membrane was incubated in blocking solution (5% nonfat milk in PBS with 0.1% Tween 20) at room temperature for 1 h. The membrane was incubated with anti-IL-1β antibody at room temperature for 2 h. After washing three times in PBS with 0.1% Tween 20, the membrane was incubated with an HRP-conjugated secondary antibody directed against the primary antibody. The membrane was developed by an enhanced chemiluminescence Western-blotting detection system (DuPont NEN Research Products, Boston MA, USA) according to the manufacturer’s instructions [8,9].
Statistical analysis: All values are given as mean ± SE.
Data analysis involved one-way ANOVA with subsequent Scheffé test.
Acknowledgment - This work was supported by a grant
form Taiwan Power Research Institute, Contract/grant number: TPC-546-2517-9802 for this investigation.
References
[1] Wikfors GH, Ferris GE, Smith BC. (1992) The relationship between gross biochemical composition of cultured algal foods and growth of the hard clam, Mercenaria mercenaria (L.). Aquaculture, 108: 135-154.
[2] Sánchez S,, Mart´ınez M, Espinola F. (2000) Biomass production and biochemical variability of the marine microalga Isochrysis galbana in relation to culture medium. Biochemical Engineering Journal, 6: 13-18.
[3] Pernet F, Tremblay R, Demers E, Roussy M. (2003) Variation of lipid class and fatty acid composition of Chaetoceros muelleri and Isochrysis sp. Grown in a semicontinuous system. Aquaculture, 221: 393–406.
[4] Lin YH, Chang FL, Tsao CY, Leu JY. (2007) Influence of growth phase and nutrient source on fatty acid composition of Isochrysis galbana CCMP 1324 in a batch photoreactor. Biochemical Engineering Journal, 37: 166-176.
[5] Cho JY, Jin HJ, Lim HJ, Whyte JNC, Hong YK. (1999) Growth activation of the microalga Isochrysis galbana by the aqueous extract of the seaweed Monostroma nitidum. Journal of Applied Phycology, 10: 561-567.
[6] Volkman JK, Smith DJ, Eglinton G, Forsberg TEV, Corner. EDS (1981) Sterol and fatty acid composition of four marine Haptophycean algae. Journal of the Marine Biological Association of the United Kingdom, 61: 509-527.
[7] Park DW, Jo Q, Lim HJ, Véron B. (2002) Sterol composition of dark-grown Isochrysis galbana and its implication in the seed production of Pacific oyster, Crassostrea gigas. Journal of Applied Phycology, 14: 351-355.
[8] Hsu HY, Hua KF, Su YC, Chu LC, Su SC, Chiu HW, Wong CH, Chen ST, Shieh CW, Yang SS, Chen YM, Chao LK. (2006) Study on regulation of cytokine gene expression in macrophages with an alkali-soluble polysaccharide of Rhizoclonium riparium alga. Journal of Agricultural and Food Chemistry, 54, 3558-3565.
[9] Hsu HY, Jeyashoke N, Yeh CH, Song YJ, Hua KF, Chao LK. (2010) Immunostimulatory bioactivity of algal polysaccharides from Chlorella pyrenoidosa activates macrophages via Toll-like receptor 4. Journal of Agricultural and Food Chemistry, 58, 927-936 [10] Fabegas J, García D, Fernandez-Alonso M, Rocha AI, Gómez-Puertas P, Escribano JM, Otero A, Coll JM. (1999) In vitro inhibition
of the replication of haemorrhagic septicaemia virus (VHSV) and African swine fever virus (ASFV) by extracts from marine microalgae. Antiviral Research, 44, 67-73.
[11] Barbarino E, Louren SO. (2005) An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. Journal of Applied Phycology, 17: 447-460.
[12] Su Y, Tsay SS, Zou W, Wu SH. (2004) Structural determination of the polar glycoglycerolipids from thermophilic bacteria Meiothermus taiwanensis. European journal of biochemistry / FEBS, 271: 4545-4551.