Colorectal Cancer and Genetic Polymorphism of DNA Double-Strand
Break Repair Gene XRCC4 in Taiwan
Da-Tian Bau2,3,Mei-Due Yang1, Yung-An Tsou3, Song-Shei Lin4, Cheng-Nan Wu5,
Hao-Hsueh Hsieh2,6, Rou-Fen Wang2, Chia-Wen Tsai2,3, Wen-Shin Chang2, Hsiu-Min
Hsieh2 and Ru-Yin Tsai2
Department of 1 Surgery and 2 Terry Fox Cancer Research Lab, China Medical University
Hospital, Taichung, Taiwan R.O.C. 3
Graduate Institute of Chinese Medical Science, and 6 Basic Medical Science, China
Medical University, Taichung, Taiwan, R.O.C. 4
Department of Medical Imaging and Radiological Sciences, Central-Taiwan University
of Science and Technology, Taichung, Taiwan, R.O.C. 5
Institute of Medical Bioscience, Central-Taiwan University of Science and Technology,
Taichung , Taiwan , R.O.C.
Correspondence to: Da-Tian Bau and Ru-Yin Tsai, Terry Fox Cancer Research Lab,
Department of Medical Research, China Medical University Hospital, 2 Yuh-Der Road,
Taichung, 404 Taiwan, R.O.C. Tel: +886 422052121 Ext 1523, Fax: +886 422052121,
e-mail: [email protected]; [email protected]
Abstract The DNA repair gene X-ray repair complementing defective
repair in Chinese hamster cells 4 (XRCC4) is thought to play a major role in the caretaking of the whole genome via double strand-break repair. However, the association of polymorphic variants of XRCC4 with colorectal cancer susceptibility has never been reported. In this hospital-based case control study, the association of XRCC4 polymorphisms C-1622T (rs7727691), G-1394T (rs6869366), G-652T (rs2075685), C-571T (rs2075686), intron 3 DIP (rs28360071), S247A (rs3734091) and intron 7 DIP (rs28360317) with colorectal cancer risk in a Taiwanese population was investigated. The genotypes of XRCC4 of 370 patients with colorectal cancer and 370 age- and gender-matched healthy controls were determined by polymerase chain
reaction-restriction fragment length polymorphism(PCR-RFLP) analysis.
We found significant differences in the genetic and allelic frequencies of the XRCC4 G-1394T between the colorectal cancer and control groups
(P=0.0003 and 8.32×10-5, respectively). The distributions of other genetic
polymorphisms between cases and the control group were not significantly different. We conclude that the G allele of XRCC4 G-1394T may contribute to colorectal carcinogenesis and may be useful for early
detection of colorectal cancer.
Colorectal cancer (CRC) is a leading cause of cancer-related morbidity and mortality worldwide. The incidence and age-adjusted mortality of CRC have drastically continued to increase in Taiwan. In recent years, the incidence and mortality of CRC has taken the third place among the common types of cancer. Etiological studies have attributed more than 85% of CRC to environmental factors (1, 2). Meat consumption, cigarette smoking and exposure to carcinogenic aromatic amines are commonly recognized environmental factors for CRC (3-5) and these DNA damage-inducing factors may induce various types of DNA adducts, including double-strand breaks (DSBs) (6). DSBs may lead to severe genome instability, which is closely related to carcinogenesis (7, 8). Two distinct DNA repair pathways are responsible for DSB repair. Homologous recombination (HR) and the non-homologous end-joining (NHEJ) (8). Components involved in NHEJ include the DNA ligase IV— XRCC4 complex, Ku70, Ku80, the catalytic subunit PKcs and artemis (9). Genetic polymorphisms in NHEJ genes influence DNA repair capacity and confer predisposition to several types of cancer, including skin (10), breast (11-13), gastric (14), and oral (15). Among these reports, the study investigating the association of XRCC4 G-1394T polymorphism with
gastric cancer found that those who had G/T or G/G at G-1394T showed a 3 to 79-fold increased risk of gastric cancer compared to those with T/T (15). Since both the stomach and intestine originate from endoderm during embryonic development, we proposed that this genetic polymorphism may also be associated with CRC susceptibility. To the best of our knowledge, there is no report studying the association of XRCC4 with CRC risk. The present study aimed at investigating the potential association of XRCC4 polymorphisms, including C-1622T (rs7727691), G-1394T (rs6869366), G-652T (rs2075685), C-571T (rs2075686), intron 3 DIP (rs28360071), S247A (rs3734091) and intron 7 DIP (rs28360317), with CRC susceptibility.
Materials and Methods
Study population and sample collection. The study population consisted
of 370 case patients and 370 cancer-free control volunteers. Three hundred and seventy patients diagnosed with CRC were recruited at the outpatient clinics of general surgery during 2002-2009 at the China Medical University Hospital, Taichung, Taiwan. The clinical characteristics of patients include histological details were all graded and
defined by expert surgeons (Dr. Yang’s team). All patients voluntarily participated, completed a self-administered questionnaire and provided peripheral blood samples. An equal number of healthy volunteers as controls were selected by matching for age, gender and some indulgences after initial random sampling from the Health Examination Cohort of the hospital. The exclusion criteria for the control group included previous malignancy, metastasized cancer from other or unknown origin, and any familial or genetic diseases. Our study was approved by the Institutional Review Board of the China Medical University Hospital and written-informed consent was obtained from all participants.
Genotyping conditions. Genomic DNA was prepared from peripheral blood leukocytes using a QIAamp Blood Mini Kit (Blossom, Taipei, Taiwan, R.O.C.) and further processed according to our previous papers (15-19). The polymerase chain reaction (PCR) cycling conditions were:
one cycle at 94oC for 5 min; 35 cycles of 94oC for 30 s, 55oC for 30 s, and
72oC for 30 s; and a final extension at 72oC for 10 min. Pairs of PCR
primer sequences and restriction enzymes for each DNA product are all listed in Table I.
Statistical analyses. Only those matches with all DNA polymorphism data (case/control=370/370) were selected for final analysis. To ensure that the controls used were representative of the general population and to exclude the possibility of genotyping error, the deviation of the genotype frequencies of XRCC4 single nucleotide polymorphism (SNPs) in the controls from those expected under the Hardy-Weinberg equilibrium was assessed using the goodness-of-fit test. Pearson’s chi-square test or Fisher’s exact test (when the expected number in any cell was less than five) was used to compare the distribution of the XRCC4 genotypes between cases and controls. Data were recognized as significant when the statistical P-value was less than 0.05.
Results
The frequencies of the genotypes for XRCC4 C-1622T, G-1394T, G-652T, C-571T, intron 3 DIP, S247A and intron 7 DIP for controls and CRC patients are shown in Table II. Genotype distribution of various genetic polymorphisms of G-1394T was significantly different between the CRC and control groups (P=0.0003), while those for C-1622T, G-652T, C-571T, S247A and intron 3 DIP and intron 7 DIP were not different
(P>0.05) (Table II).
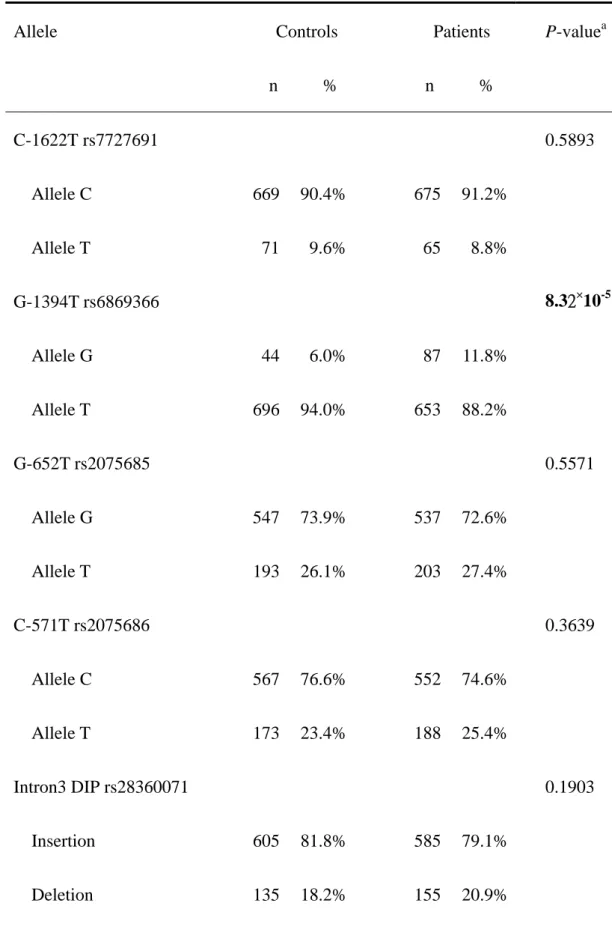
The frequencies of the alleles for the XRCC4 C-1622T, G-1394T, G-652T, C-571T, intron 3 DIP, S247A and intron 7 DIP between controls and CRC patients is shown in Table III. Allelic frequency distribution of the XRCC4 promoter -1394 was significantly different between the CRC
and control groups (P=8.32×10-5), while that of the other were not (Table
III). The conclusion deduced from Table II and III is that XRCC4 promoter -1394 G allele seems to be associated with a higher risk for CRC.
Discussion
To our knowledge, there are no studies investigating the role of XRCC4, which plays an important role in the NHEJ pathway, in CRC. According to our results, the G-1394T polymorphism of the XRCC4 gene was found for the first time to be associated with CRC (Tables II and III). According to previous study, XRCC4 is such an essential protein in taking care of our genome that critical mutations of XRCC4 will lead to cell apoptosis and possibly fatality during the embryonic stage (20). The variant polymorphism of G-1394T of XRCC4 may slightly alter the normal
expression level of XRCC4. These changes may be very subtle initially, and cause no obvious pathological change in the human body without alerting the cell via its apoptosis checking system, such as the cell cyclins and their related kinase system. If the cells undergo severe DNA damage, the cell cycle checking points will arrest resulting in either DNA repair or apoptosis. The effects may only lead to a lower capacity of the NHEJ in which the XRCC4 gene is involved. After birth, exposure to environmental carcinogens increases the possibility for genomic instability, and the need for the intracellular repair system to stem this DNA damage will increase too. In those people whose NHEJ repair system cannot perform at its normal efficiency, the raised instability of their genome can be expected. With time, these genetic deficiencies will increase the number of abnormal cells, and thus raise the risk of CRC.
It is known that DSBs are one of the most severe types of DNA damage, and if the DSBs cannot be repaired before the duplication of the genome, they will result in irreversible cellular injuries, which increases the possibility of CRC carcinogenesis, and perhaps of other types of cancer too. Some reports showed that variations of the XRCC4 gene are
the G allele of XRCC4 gene is a risk factor for CRC is consistent with the case for gastric cancer (14). A previous study showed that intestinal-type gastric carcinoma shared common pathways with CRC, indicating that the clinical subtypes of gastric cancer and CRC are both important in genetic study of cancer (22). Moreover, defects in XRCC4 function and failures of the DNA DSB repair pathway may serve as common biomarkers for early carcinogenesis detection. The phenotypes related to DNA DSB repair capacity, and the correlation between genotype and phenotype need further investigation.
In this study, we have screened seven SNPs of the XRCC4 gene and investigated their associations with susceptibility to CRC. We found that the G-1394T polymorphism of the XRCC4 gene appears to be associated with CRC risk and that the G allele is a risk factor. However, the effects of these SNPs of XRCC4 gene need further study, such as promoter assays, to reveal the role of each section or even each nucleotide in the subtle regulation of the transcriptional, translational, and post-translational expression of the XRCC4 gene. In addition, a larger population size for stratification of factors such as gender, anatomic distributions of CRC, and genomic-environmental combinatorial studies
could provide a better understanding of the complex relationship between XRCC4 and CRC carcinogenesis.
Acknowledgements
This study was supported by research grants from the China Medical University and Hospital (DMR-99-049 and CMU-97-333), Terry Fox Cancer Research Foundation and the National Science Council (NSC 98-2320-B-039-010-MY3).
References
1 Doll R and Peto R: The causes of cancer: quantitative estimates of avoidable risks of cancer in the United States today. J Natl Cancer Inst 66: 1191-1308, 1981.
2 Thomas HJ: Familial colorectal cancer. BMJ 307: 277-278, 1993.
3 Giovannucci E, Colditz GA, Stampfer MJ, Hunter D, Rosner BA, Willett WC and Speizer FE: A prospective study of cigarette smoking and risk of colorectal adenoma and colorectal cancer in U.S. women. J Natl Cancer Inst 86: 192-199, 1994.
4 Giovannucci E, Rimm EB, Stampfer MJ, Colditz GA, Ascherio A, Kearney J and Willett WC: A prospective study of cigarette smoking and risk of colorectal adenoma and colorectal cancer in U.S. men. J Natl Cancer Inst 86: 183-191, 1994.
5 Heineman EF, Zahm SH, McLaughlin JK and Vaught JB: Increased risk of colorectal cancer among smokers: results of a 26-year follow-up of US veterans and a review. Int J Cancer 59: 728-738, 1994.
6 Pryor WA, Hales BJ, Premovic PI and Church DF: The radicals in cigarette tar: their nature and suggested physiological implications. Science 220: 425-427, 1983.
7 Goode EL, Ulrich CM and Potter JD: Polymorphisms in DNA repair genes and associations with cancer risk. Cancer Epidemiol Biomarkers Prev 11: 1513-1530, 2002.
8 Karran P: DNA double-strand break repair in mammalian cells. Curr Opin Genet Dev 10: 144-150, 2000.
9 Burma S, Chen BP and Chen DJ: Role of non-homologous end joining (NHEJ) in maintaining genomic integrity. DNA Repair (Amst) 5: 1042-1048, 2006. 10 Chang CH, Chiu CF, Wang HC, Wu HC, Tsai RY, Tsai CW, Wang RF, Wang
CH, Tsou YA and Bau DT: Significant association of ERCC6 single nucleotide polymorphisms with bladder cancer susceptibility in Taiwan. Anticancer Res 29: 5121-5124, 2009.
11 Bau DT, Fu YP, Chen ST, Cheng TC, Yu JC, Wu PE and Shen CY: Breast cancer risk and the DNA double-strand break end-joining capacity of nonhomologous end-joining genes are affected by BRCA1. Cancer Res 64: 5013-5019, 2004.
12 Bau DT, Mau YC, Ding SL, Wu PE and Shen CY: DNA double-strand break repair capacity and risk of breast cancer. Carcinogenesis 28: 1726-1730, 2007. 13 Chiu CF, Wang HC, Wang CH, Wang CL, Lin CC, Shen CY, Chiang SY and
Bau DT: A new single nucleotide polymorphism in XRCC4 gene is associated with breast cancer susceptibility in Taiwanese patients. Anticancer Res 28: 267-270, 2008.
14 Chiu CF, Wang CH, Wang CL, Lin CC, Hsu NY, Weng JR and Bau DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with gastric cancer susceptibility in Taiwan. Ann Surg Oncol 15: 514-518, 2008. 15 Chiu CF, Tsai MH, Tseng HC, Wang CL, Wang CH, Wu CN, Lin CC and Bau
DT: A novel single nucleotide polymorphism in XRCC4 gene is associated with oral cancer susceptibility in Taiwanese patients. Oral Oncol 44: 898-902, 2008.
16 Bau DT, Tseng HC, Wang CH, Chiu CF, Hua CH, Wu CN, Liang SY, Wang CL, Tsai CW and Tsai MH: Oral cancer and genetic polymorphism of DNA double-strand break gene Ku70 in Taiwan. Oral Oncol 44: 1047-1051, 2008. 17 Chiu CF, Tsai MH, Tseng HC, Wang CL, Tsai FJ, Lin CC and Bau DT: A
novel single nucleotide polymorphism in ERCC6 gene is associated with oral cancer susceptibility in Taiwanese patients. Oral Oncol 44: 582-586, 2008. 18 Hsu CF, Tseng HC, Chiu CF, Liang SY, Tsai CW, Tsai MH and Bau DT:
Association between DNA double-strand break gene Ku80 polymorphisms and oral cancer susceptibility. Oral Oncol 45: 789-793, 2009.
Wang HC, Chiu CF and Bau DT: Interaction of EXO1 genotypes and smoking habit in oral cancer in Taiwan. Oral Oncol 45: e90-94, 2009.
20 Gao Y, Ferguson DO, Xie W, Manis JP, Sekiguchi J, Frank KM, Chaudhuri J, Horner J, DePinho RA and Alt FW: Interplay of p53 and DNA-repair protein XRCC4 in tumorigenesis, genomic stability and development. Nature 404: 897-900, 2000.
21 Han J, Colditz GA, Samson LD and Hunter DJ: Polymorphisms in DNA double-strand break repair genes and skin cancer risk. Cancer Res 64: 3009-3013, 2004.
22 Shimada S, Tashima S, Yamaguchi K, Matsuzaki H and Ogawa M: Carcinogenesis of intestinal-type gastric cancer and colorectal cancer is commonly accompanied by expression of brain (fetal)-type glycogen phosphorylase. J Exp Clin Cancer Res 18: 111-118, 1999.
Table I. The primer sequences, polymerase chain reaction and restriction fragment length
polymorphism (PCR-RFLP) conditions for XRCC4 gene polymorphisms.
Polymorphism
(location)
Primer sequence (5’->3’) Restriction
enzyme SNP sequence DNA fragment size (bp) C-1622T rs77277691 F: AAGATACTGAGACACTAATC R: CACAACATAACTAAGGATGA Fnu4H I T C 218 bp 32 + 186 bp G-1394T rs6869366 F: GATGCGAACTCAAAGATACTGA R: TGTAAAGCCAGTACTCAAACTT Hinc II T G 300 bp 200 + 100 bp G-652T rs2075685 F: GCTAGACACCACTCCAATAA R: GGCTACGTAGATTATGTGTG Mbo II T G 326 bp 127 + 199 bp C-571T rs2075686 F: GGCTACTGACTAAACAGATG R: TAACACGTTGGCTACGTAGA Mnl I C T 197 bp 69 + 128 bp Intron 3 rs28360071 F: TCCTGTTACCATTTCAGTGTTAT R: CACCTGTGTTCAATTCCAGCTT Insertion Deletion 139 bp 109 bp Codon 247 rs3734091 F: GCTAATGAGTTGCTGCATTTTA R: TTTCTAGGGAAACTGCAATCTGT Bbs I C A 308 bp 204 + 104 bp
Intron 7 rs28360317 F1 (CCT-positive): ATACTGTGTTTGGAACTCCT F2 (CCT-negative): ATACTGTGTTTGGAACTAGA R: TATCCTATCATCTCTGGATA CCT - 239 bp No product
Table II. Distribution of XRCC4 genotypes among colorectal cancer patient and control groups. Controls Patients Genotype n % n % P-valuea C-1622T rs7727691 0.6352 CC 299 80.8% 305 82.4% CT 71 19.2% 65 17.6% TT 0 0.0% 0 0.0% G-1394T rs6869366 0.0003 GG 0 0.0% 3 0.8% GT 44 11.9% 81 21.9% TT 326 88.1% 286 77.3% G-652T rs2075685 0.8140 GG 191 51.6% 183 49.5% GT 165 44.6% 171 46.2% TT 14 3.8% 16 4.3% C-571T rs2075686 0.6529 CC 216 58.4% 204 55.1% CT 135 36.5% 144 38.9% TT 19 5.1% 22 6.0% Intron3 DIP rs28360071 0.4096 II 246 66.5% 229 61.9% ID 113 30.5% 127 34.3% DD 11 3.0% 14 3.8%
Codon247 rs3734091 0.2597 AA Ser/Ser 0 0.0% 0 0.0% AC Ser/Ala 67 18.1% 74 20.0% CC Ala/Ala 303 81.9% 269 80.0% Intron7 DIP rs28360317 0.5605 II 182 49.2% 168 45.4% ID 154 41.6% 163 44.1% DD 34 9.2% 39 10.5% a
Table III. Distribution of XRCC4 alleles among the colorectal cancer patient and control groups. Allele Controls n % Patients n % P-valuea C-1622T rs7727691 0.5893 Allele C 669 90.4% 675 91.2% Allele T 71 9.6% 65 8.8% G-1394T rs6869366 8.32×10-5 Allele G 44 6.0% 87 11.8% Allele T 696 94.0% 653 88.2% G-652T rs2075685 0.5571 Allele G 547 73.9% 537 72.6% Allele T 193 26.1% 203 27.4% C-571T rs2075686 0.3639 Allele C 567 76.6% 552 74.6% Allele T 173 23.4% 188 25.4% Intron3 DIP rs28360071 0.1903 Insertion 605 81.8% 585 79.1% Deletion 135 18.2% 155 20.9%
Codon247 rs3734091 0.5954 Allele A (Ser) 67 9.1% 74 10.0% Allele C (Ala) 673 90.9% 666 90.0% Intron7 DIP rs28360317 0.2868 Insertion 518 70.0% 499 67.4% Deletion 222 30.0% 241 32.6% a