of August 5, 2012.
This information is current as
Conditions
Polarizing
−
Lymphocytes under IL-17
T Cells Disarm or Delete Cytotoxic T
+
CD4
Shen and Hsin-Wei Chen
Ann-Joy Cheng, Pele Chong, Ching-Liang Chu, Chia-Rui
Chen, Shih-Jen Liu, Joseph T. Chang, Chun-Ta Liao,
Jy-Ping Tsai, Meng-Hua Lee, Shu-Ching Hsu, Mei-Yu
http://www.jimmunol.org/content/189/4/1671
doi: 10.4049/jimmunol.1103447
July 2012;
2012; 189:1671-1679; Prepublished online 13
J Immunol
References
http://www.jimmunol.org/content/189/4/1671.full#ref-list-1
, 22 of which you can access for free at:
cites 46 articles
This article
Subscriptions
http://jimmunol.org/subscriptions
is online at:
The Journal of Immunology
Information about subscribing to
Permissions
http://www.aai.org/ji/copyright.html
Submit copyright permission requests at:
Email Alerts
http://jimmunol.org/cgi/alerts/etoc
Receive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606.
Immunologists, Inc. All rights reserved.
Copyright © 2012 by The American Association of
9650 Rockville Pike, Bethesda, MD 20814-3994.
The American Association of Immunologists, Inc.,
is published twice each month by
The Journal of Immunology
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
CD4
+
T Cells Disarm or Delete Cytotoxic T Lymphocytes
under IL-17–Polarizing Conditions
Jy-Ping Tsai,*
,1Meng-Hua Lee,
†,‡,1Shu-Ching Hsu,*
,x,{Mei-Yu Chen,* Shih-Jen Liu,*
,‖Joseph T. Chang,
#Chun-Ta Liao,** Ann-Joy Cheng,
†Pele Chong,*
,‖Ching-Liang Chu,*
,2Chia-Rui Shen,
†,‡and Hsin-Wei Chen*
,‖Previous studies have shown that TGF-b acts cooperatively with IL-6 to elicit a high frequency of IL-17–secreting CD4+T cells
(termed Th17) and an elevated CD8+IL-17+T cell population (termed Tc17). These CD8+cells fail to behave like most cytotoxic T lymphocytes that express IFN-g and granzyme B, but they exhibit a noncytotoxic phenotype. Although a significant increase in the number of these Tc17 cells was found in tumors, their role and interaction with other cell types remain unclear. In this study, we demonstrate that the presence of CD4+CD252T cells, but not the CD4+CD25+(regulatory T [Treg]) cell population, signif-icantly reduced the elicitation of Tc17 cells, possibly as a result of the induction of apoptotic signals. Importantly, these signals may be derived from soluble mediators, and the addition of anti–IL-2 restored the reduction of Tc17 cells in the presence of CD4+ CD252 T cells. Finally, the elicited Tc17 and Treg cells exhibited a close association in patients with head and neck cancer, indicating that the surrounding Treg cells might maintain the survival of the Tc17 cells. Taken together, these results reveal an intriguing mechanism in which Tc17 cells are controlled by a finely tuned collaboration between the different types of CD4+T cells in distinct tumor microenvironments. The Journal of Immunology, 2012, 189: 1671–1679.
C
D4+T cells play important roles in orchestrating immune responses. Early work has shown that CD4+T cells play a crucial role in the generation of active effector CTLs (1, 2). Several models have been proposed that depict the mechanisms by which CD4+T cells help CTL responses (3–5). Sakaguchi et al. (6) demonstrated that in contrast to activated Th cells, regulatory T (Treg) cells constitutionally express CD25. The development and function of CD4+CD25+ Treg cells are controlled by the transcription factor Foxp3 (7, 8). Several pioneering studies have revealed that the depletion of CD25+T cells enhances the functionof CTLs and promotes the clearance of viruses (9) and tumors (10, 11). Furthermore, Treg cells suppress CTL effector functions by inhibiting CTL degranulation (12, 13), and they modulate CTL expansion and effector differentiation (14–16). It is clear that the function and differentiation of CD8+T cells are critically depen-dent on the subset of CD4+T cells with which they interact.
Conventional CTLs express CD8 molecules as coreceptors that recognize and respond to peptides that are presented by MHC class I molecules. The CTLs act as the guided missiles of the immune system. When CTLs are aroused from a naive state, they can target and kill Ag-specific target cells. In general, CTLs are involved in the host defense during acute and chronic infections with various pathogens, and they also contribute to the elimination of trans-formed cells (5). The fate and function of CTLs depend largely upon the local cytokine milieu at the time of the initial interaction with the immunogen. In a previous study (17), we showed that TGF-b and IL-6 induced IL-17–secreting CD8+ T cells with a noncytotoxic phenotype (Tnc17). IFN-g and IL-10 were not pro-duced in IL-17–secreting CD8+ T cells. These IL-17–secreting CD8+T cells are fundamentally different from conventional CTLs and represent a distinct subset of CD8+T cells. Because our initial presentation of our findings, several independent research groups reported that IL-17–secreting CD8+T cells are negative for cy-tolytic activity and express low levels of the CTL markers (18–21). These IL-17–secreting CD8+T cells are now termed Tc17 cells.
Recently, we found that there were high levels of TGF-b, IL-6, and PGE2in the malignant effusions of cancer patients (22).
TGF-b and IL-6 are the key factors in the development of the Th17 (23‑25) and Tc17 (18–21) cell lineages. It has been demonstrated that PGE2stimulates IL-23 expression (26, 27). IL-23 is an
im-portant cytokine for Th17 survival and expansion (25), and this environment should favor the development of IL-17–producing T cells. Indeed, the frequency of IL-17–producing cells was sig-nificantly elevated in the blood, bone marrow, and spleen of B16 melanoma-bearing mice (28). Furthermore, high numbers of CD4+
IL-17+and CD8+IL-17+T cells were detected in human head and
*National Institute of Infectious Diseases and Vaccinology, National Health Research Institutes, Miaoli 350, Taiwan;†Department of Medical Biotechnology and
Labora-tory Science, College of Medicine, Chang Gung University, Taoyuan 333, Taiwan;
‡
Graduate Institute of Biomedical Sciences, College of Medicine, Chang Gung Uni-versity, Taoyuan 333, Taiwan;xGraduate Institute of Life Sciences, National Defense Medical Center, Taipei 114, Taiwan;{Ph.D. Program in Tissue Engineering and Regenerative Medicine, National Chung Hsing University, Taichung 402, Taiwan;
‖Graduate Institute of Immunology, China Medical University, Taichung 402, Taiwan; #Department of Radiation Oncology, Chang Gung Memorial Hospital, Taoyuan 333,
Taiwan; and **Department of Otorhinolaryngology, Head and Neck Surgery, Chang Gung Memorial Hospital, Taoyuan 333, Taiwan
1J.-P.T. and M.-H.L. contributed equally to this work.
2Current address: Graduate Institute of Immunology, National Taiwan University,
College of Medicine, Taipei, Taiwan.
Received for publication November 30, 2011. Accepted for publication June 15, 2012.
This work was supported by Grants VC-098-PP-01, VC-099-PP-01, and VC-100-PP-01 from the National Health Research Institutes (to H.-W.C.) and Grants BMRP440 and EMRPD140041 from Chang Gung Memorial Hospital and Chang Gung Univer-sity (to C.-R.S.).
Address correspondence and reprint requests to Dr. Hsin-Wei Chen or Dr. Chia-Rui Shen, National Institute of Infectious Diseases and Vaccinology, National Health Research Institutes, Miaoli 350, Taiwan (H.-W.C.) or Department of Medical Bio-technology and Laboratory Science, Chang Gung University, Taoyuan 333, Taiwan (C.-R.S.). E-mail addresses: [email protected] (H.-W.C.) and [email protected]. edu.tw (C.-R.S.)
Abbreviations used in this article: HNSCC, head and neck squamous cell carcinoma; Treg, regulatory T.
CopyrightÓ 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00 www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103447
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
neck cancers, melanoma, prostate cancer, and sarcoma tissues (28). However, the relationship between the IL-17–producing T cells and tumor development is still controversial (29–32).
Although the effects of CD4+ T cells on conventional CTL development and memory have been widely studied, the modu-lation of Tc17 cells by CD4+ T cells is not well understood. Therefore, this study focuses on the role of CD4+T cells in the differentiation pathway of Tc17 cells. We observed that CD4+ T cells inhibited the development of Tc17 cells. This inhibitory effect is mediated by CD4+CD252T cells but not by CD4+CD25+ T cells. In contrast, Treg cells maintain the survival and expansion of Tc17 cells. We also noticed that Tc17 and Treg cells were enriched in patients with head and neck squamous cell carcinoma (HNSCC). All the information pointed to a strong positive cor-relation between the frequency of Tc17 and Treg cells. In this study, we provide evidence that Treg cells paralyze CTLs by fa-cilitating their differentiation into Tc17 cells, which may be a common mechanism for tumor escape.
Materials and Methods
Mice
Female C57BL/6 mice were purchased from the National Laboratory Animal Center (Taipei, Taiwan). All the mice were housed at the Laboratory Animal Center of the National Health Research Institutes. OT-1 (OVA-specific CD8+TCR transgenic) mice were bred at the Laboratory Animal
Center of the National Health Research Institutes. All the animal studies were approved and were performed in compliance with the guidelines of the Animal Committee of the National Health Research Institutes.
Reagents and Abs
The FITC-, PE-, allophycocyanin-, and PerCP-conjugated Abs specific to CD4, CD8, CD25, IFN-g, granzyme B, and IL-17 were purchased from BD Biosciences (San Diego, CA) or eBioscience (San Diego, CA). The puri-fied anti-CD3 (145-2C11), anti-CD28 (37-51), anti–IFN-g (XMG-1.2), and anti–IL-4 (11B11) mAbs were purchased from eBioscience. The recom-binant human TGF-b and the recomrecom-binant mouse IL-6 were purchased from R&D Systems (Minneapolis, MN). The CellTracker Orange CMTMR fluorescent probe was purchased from Molecular Probes (Eugene, OR). The mouse cells were cultured in complete RPMI 1640 medium (Hyclone) sup-plemented with 5% FBS, 100 U/ml penicillin, 100 mg/ml streptomycin, 2 mM glutamine, 10 mM HEPES, and 50 mM 2-ME.
Isolation of lymphocyte subsets
Murine splenocytes were harvested from 5- to 12 wk-old C57BL/6 mice. The CD4+, CD8+, and CD42CD82 T cell subsets were isolated using
FACS (BD FACSAria Flow Cytometer). In some experiments, we used flow cytometry-sorted CD4+CD252, CD4+CD25+, CD8+, and CD42CD82
T cell subsets. Each subset was.98% pure.
In vitro cocultures and Tc17 cell polarization
Purified murine CD8+T cells (23 105/well), CD42CD82cells (83 105/ well), and other cell subsets were cocultured in 96-well flat-bottom cell culture plates at indicated ratios. The T cells were stimulated with anti-CD3 (4 mg/ml) plus anti-CD28 (2 mg/ml) Abs under IL-17–polarizing conditions for 3 d. The IL-17–polarizing conditions were defined by the initiation of an activation media containing recombinant human TGF-b (1 ng/ml), re-combinant mouse IL-6 (100 ng/ml), purified anti–IFN-g (8 mg/ml), and anti–IL-4 (8 mg/ml) mAbs. In some experiments, the culture medium was replaced with fresh medium that did not contain exogenous cytokine or Ab but was complemented with recombinant human IL-2 (40 U/ml), and in-cubation was continued for an additional 3 d. Half of the culture medium was removed from each well and was replaced with fresh medium con-taining anti-CD3 (4 mg/ml) and anti-CD28 (2 mg/ml) Abs. After an addi-tional 2-d incubation, the cells were harvested for further analysis.
Millicell hanging cell culture system
Purified CD8+T cells were cocultured with CD42CD82cells at the in-dicated ratios in 24-well tissue culture plates. CD4+T cells were seeded in Millicell hanging cell culture inserts (polyethylene terephthalate mem-branes with 0.4-mm pores) (Millipore, Bedford, MA) to prevent their direct contact with the underlying cells.
Intracellular staining of murine cells
The cells were stimulated with 10 ng/ml PMA (Sigma-Aldrich) and 1 mg/ml ionomycin (Sigma-Aldrich) in the presence of brefeldin A for 4 h at 37˚C prior to staining with fluorescence-conjugated anti-CD4 or anti-CD8 Abs. The cells were fixed and were permeabilized using a cytofix/cytoperm kit (eBioscience) in accordance with the manufacturer’s instructions. Intra-cellular staining procedures were performed by staining with the appro-priate Abs.
Cytotoxicity assays
A standard [51Cr] release assay was performed to evaluate cytotoxic
ac-tivity. In brief, purified OT-1 CD8+T cells (23 105/well), CD42CD82 cells (83 105/well), and other cell subsets were cocultured in 96-well flat-bottom cell culture plates at indicated ratios. Effector T cells were stim-ulated with anti-CD3 (4 mg/ml) plus anti-CD28 (2 mg/ml) Abs under neutral or IL-17–polarizing conditions for 3 d. EL-4 target cells were pulsed with OVA peptide (SIINFEKL; 10 mg/ml) and Na2[
51
Cr]O4(100
mCi) for 1 h. After three washes, these effector and target cells (5 3 103
cells/well) were plated in round-bottom 96-well plates at proper E:T ratios and incubated for 5 h. The amount of released [51Cr] in supernatants was
counted, and the percentage of specific target cell killing was calculated according to the formula [(sample release2 spontaneous release)/(total release2 spontaneous release)] 3100.
Monitoring of cell division
Flow cytometry-sorted CD8+T cells were suspended in 1 ml serum-free
medium in the presence of the CellTracker Orange CMTMR probe at a concentration of 10 mM. The cells were incubated at 37˚C for 10 min and were washed with serum-free RPMI 1640 medium. The CD8+T cells were then cocultured with other cell subsets. After 3 d of culture, IL-17 intra-cellular staining was performed to visualize the division of the IL-17+ CD8+cells.
Annexin V staining
The cells were stimulated with 10 ng/ml PMA and 1 mg/ml ionomycin for 4 h at 37˚C, followed by direct surface staining with PerCP-conjugated anti-CD8 and PE-conjugated anti–IL-17 Abs. The Annexin V-FITC Apo-ptosis Detection kit (eBioscience) was used in accordance with the man-ufacturer’s instructions.
Effect of IL-2 on Tc17 cells
Anti-mouse IL-2, anti-mouse IL-27p28 Abs, and recombinant mouse IL-2 were purchased from R&D Systems (Minneapolis, MN). The anti-mouse IL-2 and anti-mouse IL-27p28 Abs were added to the cocultured cells at a concentration of 5 mg/ml. At the beginning of the cell stimulation 10, 1, or 0.1 ng/ml murine IL-2 was added to each sample.
Clinical samples
The human research study protocol was approved by the Ethical Committee of Linkuo Chang Gung Memorial Hospital (Taipei, Taiwan), and written informed consent was obtained from all the participants. Peripheral blood was obtained by vein puncture. Healthy volunteers served as a control group. In total, 151 samples were collected from 25 healthy volunteers and from 126 untreated HNSCC patients. The patients were divided into two groups: those in the early stage of the disease (n = 44) corresponding to stages I and II according to the Tumor Node Metastasis staging for HNSCC, and those in the advanced stage of the disease corresponding to stages III and IV (n = 82). Biopsies from the tumors and the grossly normal mucosal tissues were obtained from the subjects. Total RNA was extracted from the biopsy specimens and was subjected to real-time PCR analysis to determine the IL-6, IL-17, and TGF-b mRNA levels, respectively.
Real-time PCR
The expression of IL-6, IL-17, TGF-b, and GAPDH was analyzed by real-time PCR, using iQ SYBR Green and a BioRad iCycler iQ5 (Bio-Rad, Hercules, CA). GAPDH (forward primer, 59-ATGCTGGCCTGAGTAC-39; reverse primer, 59-TGAGTCCTTCCACGATAC-39) was used as a house-keeping gene. IL-6, IL-17, and TGF-b were amplified using the following primers: IL-6, forward primer, 59-TAGCCGCCCCACACAGACAG-39, and reverse primer, 59-GGCTGGCATTTGTGGTTGGG-39; IL-17, forward primer, 59-AATCTCCACCGCAATGAGGA-39, and reverse primer, 59-AC-GTTCCCATCAGCGTTGA-39; TGF-b, forward primer, 59-ACCGGCCT-TTCCTGCTTCTCA-39, and reverse primer, 59-CGCCCGGGTTATGCTG-GTTGT-39. The reaction parameters for GAPDH and the above cytokines
1672 CTLs PARALYZED UNDER IL-17–POLARIZING CONDITIONS
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
were as follows: an initial denaturation at 95˚C for 3 min, followed by 40 cycles at 95˚C for 3 s, 60˚C for 45 s, and 72˚C for 45 s.
Intracellular staining of human cells
PBMC were obtained from heparinized peripheral blood by density gradi-ent cgradi-entrifugation using Ficoll-Paque Plus (GE Healthcare BioSciences). PBMCs (23 106) were stimulated with 50 ng/ml PMA and 1 mg/ml ionomycin in RPMI 1640 medium (Life Technologies) containing 10% FBS (HyClone), and the cells were incubated in 5% CO2humidified air at
37˚C for 4 h. Brefeldin A (BD Biosciences) (10 mg/ml) was added to the cells 3 h before the cells were harvested. After harvesting, the cells were aliquoted into tubes, were spun, and were washed once with PBS. For the IL-17–producing cells, the cells were incubated with a FITC-conjugated anti-human CD4 or anti-CD8 mAb at 4˚C for 30 min. After fixation and permeabilization were performed in accordance with the manufacturer’s instructions, the surface-labeled cells were stained with PE-conjugated anti-human IL-17 and/or APC anti-anti-human FOXP3. Isotype controls were added to confirm the Ab specificity.
Flow cytometry experiments
The data were acquired using the CellQuest Pro software on a BD FACSCalibur flow cytometer and were analyzed using the FACS 3 software.
Statistical analyses
Statistical analyses were performed using the GraphPad Prism software, version 5.02 (GraphPad Software). The statistical significance of the differences between the groups was assessed using a one-tailed Student t test. The differences with p, 0.05 were considered statistically sig-nificant.
Results
CD4+T cells inhibit Tc17 cell development
To investigate the role of CD4+T cells on the development of Tc17 cells, CD8+T cells that were purified by FACS were stimulated with anti-CD3/anti-CD28 Abs in the presence or absence of in-creasing numbers of purified CD4+T cells under IL-17–polarizing conditions for 3 d. The CD8+T cells were gated and shown in Fig. 1A. A substantial number of Tc17 cells were induced in the ab-sence of CD4+T cells. More than 98% of the IL-17–producing CD8+T cells did not secrete IFN-g. The percentage of Tc17 cells was significantly diminished in the presence of CD4+T cells. The percentage of CD8+IL-17+T cells was reduced by.50% in the presence of CD4+T cells at a CD4+/CD8+ratio of 1:1. Moreover, this suppressive effect was enhanced as the number of CD4+T cells increased. The results from three independent experiments are sum-marized in Fig. 1B.
We next examined if cell-to-cell contact was required for the inhibition of Tc17 cells development by CD4+T cells. To this end, the direct cell-to-cell contact between CD4+and CD8+T cells was blocked using a Transwell culture system. The inhibition of Tc17 cell development occurred in the absence of cellular interaction, and an inhibition of∼40% was observed (36.3 versus 22.5%; Fig. 1C). The results from four independent experiments are summa-rized in Fig. 1D. The observation that direct cell contact did not play a significant role in the inhibition of Tc17 cell differentiation suggests that soluble factor(s) are involved in the suppression of Tc17 cell development.
The inhibition of Tc17 cell development is mediated by CD4+ CD252T cells and IL-2
A separate set of experiments was performed to elucidate if the natural Treg cells (CD4+CD25+) mediated the inhibition of Tc17 cell development. We purified CD4+CD25+ and CD4+CD252 T cells, which represent the natural Treg and non-Treg subsets, respectively. The two CD4+cell subsets were cultured with CD8+ T cells at different ratios (Fig. 2A). The cocultured cells were stimulated with anti-CD3/anti-CD28 Abs under IL-17–polarizing
conditions for 3 d. Tc17 cells were detected by intracellular staining. The generation of CD8+IL-17+T cells was∼45% in the presence of CD4+CD25+T cells, which was comparable to that obtained with CD8+ T cells cultured alone (47%). This result demonstrates that CD4+CD25+ T cells do not prevent the de-velopment of Tc17 cells. In contrast, CD4+CD252T cells sig-nificantly suppressed the production of Tc17 cells in a dose-dependent manner (Fig. 2A). We confirmed this finding in four independent experiments (Fig. 2B). In addition, the majority of Tc17 cells expressed low levels of granzyme B (Fig. 2A). We further performed the [51Cr] release assay to evaluate the cyto-toxicity of CD8+T cells. As shown in Fig. 2C, little cytotoxicity was detected when CD8+ T cells were activated under IL-17– polarizing conditions. In contrast, remarkable killing activities were induced in the neutral conditions. These results suggest that under the experimental conditions used in this study, naive CD8+ T cells differentiate into Tc17 cells that exhibit a limited cytotoxic phenotype.
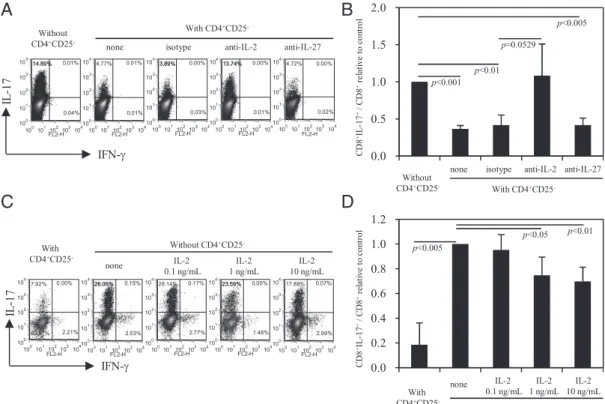
IL-2 (33) and IL-27 (34, 35) have been shown to inhibit the Th17 cell differentiation, and our results demonstrate that soluble factors are involved in the suppression of Tc17 cell development (Fig. 1C, 1D). We next assessed the role of IL-2 and IL-27 in the suppression of Tc17 cell differentiation. CD8+T cells were
cul-tured with CD4+CD252T cells at a ratio of 1:2. The cocultured
FIGURE 1. The inhibition of Tc17 cell development is mediated by CD4+T cells and does not require cell-to-cell interaction. Sorted CD4+ and CD8+T cells from naive C57BL/6 splenocytes were activated with anti-CD3/anti-CD28 Abs under optimal IL-17–polarizing conditions for 3 d. Intracellular cytokine staining was performed 4 h after restimulation of the cells with PMA–ionomycin/brefeldin A. (A) CD8+
T cells were gated, and the expression of IL-17 and IFN-g was analyzed. The numbers indicate the percentages of cytokine-positive cells in each quadrant. The results from a representative experiment are shown. (B) The means and SDs obtained from three independent experiments are shown. Significant differences were determined using Student t test, and the p values are indicated. (C) Sorted CD8+T cells were directly or indirectly cocultured
with sorted CD4+ T cells (at a 1:2 ratio) using a Transwell system to
separate the CD4+cells from the CD8+T cells. The expression of IL-17
and IFN-g by CD8+T cells was analyzed after 3 d of culture. The numbers
represent the percentages of cytokine-positive cells in each quadrant. The results from a representative experiment are shown. (D) The means and SDs obtained from four independent experiments are shown. Significant differences were determined using Student t test, and the p values are indicated.
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
cells were stimulated under IL-17–polarizing conditions for 3 d, as described previously. CD8+T cells cultured alone served as a control. Anti–IL-2 or anti–IL-27 Abs were added at the beginning of the stimulation process to neutralize the IL-2 and IL-27, re-spectively. As shown in Fig. 3A, CD8+IL-17+T cell expression was∼15% when cultured with CD8+T cells alone, whereas in the presence of CD4+CD252T cells, the percentage of CD8+IL-17+ cells increased from 4.8 to 13.7% following the addition of the anti–IL-2 Ab. This restoration was not observed following the
addition of anti–IL-27 or the isotype control Abs. Fig. 3B sum-marizes the results from three independent experiments.
Because the anti–IL-2 Ab reversed the suppression that was mediated by the CD4+CD252T cells, we next examined the effect of exogenous rIL-2 on the Tc17 cell differentiation pathway. rIL-2 (10, 1, or 0.1 ng/ml) was added to CD8+T cells at the start of the cell activation process under IL-17–polarizing conditions for 3 d. CD8+T cells cocultured with CD4+CD252 T cells served as a control. As shown in Fig. 3C, the generation of Tc17 cells was suppressed by.30% when exogenous rIL-2 (10 ng/ml) was added, and this suppression was dose dependent. The results from four independent experiments are summarized in Fig. 3D. These results demonstrate that IL-2 is a soluble factor that potentially mediates the inhibition of Tc17 cell development.
CD4+CD252T cells promote the apoptosis of Tc17 cells To study how CD4+CD252T cells contributed to the suppression of Tc17 cell differentiation programming, CD8+ T cells were labeled with CellTracker Orange and were activated with anti-CD3/anti-CD28 Abs in the presence of different ratios of CD4+ CD252 or CD4+CD25+ T cells. The results showed that the frequency of CD8+IL-17+cells was reduced by 70% (from 41.3 to 12.6%) in the presence of CD4+CD252T cells at a 1:1 ratio in a ratio-dependent manner (Fig. 4A). No noticeable effect on CD8+IL-17+was observed in the presence of CD4+CD25+T cells (Fig. 4A). These results clearly demonstrate that CD4+CD252 T cells, but not CD4+CD25+T cells, play a suppressive role in the generation of Tc17 cells. However, we were surprised to observe that the proliferative capacity of CD8+ T cells, as assessed by CellTracker Orange dilution, was not limited by CD4+CD25+ cells or CD4+CD252T cells (Fig. 4A). Therefore, we speculated that CD4+CD252T cells eliminate Tc17 cells by increasing the apoptosis of these cells. Because IL-17 is also expressed on the cell surface (36), we combined IL-17 surface staining and FITC-conjugated annexin V labeling to evaluate the apoptosis of Tc17 cells. Gated CD8+IL-17+and CD8+IL-172cells are shown in the upper panels and lower panels of Fig. 4B, respectively. Within the CD8+IL-17+T cell subset, in the absence of CD4+T cells, 24%
of the cells were stained positive for annexinV. When CD8+ T cells were cocultured with CD4+CD25+T cells, the percentage of CD8+IL-17+annexinV+ cells was 28.1%, which was compa-rable to that obtained with CD8+T cells cultured alone. In con-trast, the percentage of CD8+IL-17+annexinV+ cells increased significantly when CD8+ T cells were cocultured with CD4+ CD252 T cells, and the increase was a function of the CD4+ CD252/CD8+ ratio. Interestingly, the percentage of apoptotic annexinV+cells did not change significantly within the CD8+ IL-172 T cell subset. These results were confirmed in three inde-pendent experiments (Fig. 4C, 4D).
Next, we analyzed the expression profiles of the anti-apoptotic Bcl-2 protein and the active form of the apoptosis-related caspase 3 in CD8+IL-17+T cells (Fig. 5). As shown in Fig. 5A, in the ab-sence of CD4+T cells, 94.5% of CD8+IL-17+T cells exhibited a Bcl-2+caspase 32 phenotype. When CD4+CD25+cells were
cocultured with CD8+T cells at a 1:1 ratio, the percentage of CD8+IL-17+T cells with the Bcl-2+caspase 32 phenotype in-creased to 95.8%. In contrast, in the presence of CD4+CD252 T cells, the number of CD8+IL-17+ T cells decreased as the number of CD4+CD252T cells increased. Interestingly, there was a higher percentage of CD8+IL-17+T cells with a Bcl-22caspase 3+phenotype in the presence of CD4+CD252T cells than in the
absence of CD4+T cells. This enhancement was dependent on the number of CD4+CD252 T cells. When CD8+T cells were
cocultured with CD4+CD25+T cells, the percentage of CD8+
IL-FIGURE 2. CD4+CD252T cells, but not CD4+CD25+T cells, mediate
the inhibition of Tc17 cell development. Sorted CD4+CD25+(Treg cells)
or CD4+CD252(non-Treg cells) T cells were cocultured with sorted CD8+ T cells at the indicated cell ratios. The cells were stimulated with anti-CD3/anti-CD28 Abs under optimal IL-17–polarizing conditions for 3 d. (A) The expression of IL-17 and granzyme B by CD8+T cells was as-sessed by intracellular staining. The numbers represent the percentages of cytokine-positive cells in each quadrant. A representative experiment is shown. (B) The means and SDs obtained from four independent experi-ments are shown. Significant differences were determined using Student t test, and the p values are indicated. (C) Standard [51
Cr] release assays were used to evaluate cytotoxicity. Effector cells stimulated with anti-CD3/anti-CD28 Abs alone (neutral conditions) were used as controls. The means and SDs obtained from triplicate wells are shown. A representative of three experiments is shown.
1674 CTLs PARALYZED UNDER IL-17–POLARIZING CONDITIONS
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
17+Bcl-22caspase 3+cells was 0.18%, and this was comparable to the percentage observed with CD8+ T cells cultured alone. The data from three independent experiments are summarized in Fig.
5B and 5C. Taken together, these results suggest that CD4+CD25+ T cells maintain the survival of Tc17 cells, whereas CD4+CD252 T cells promote the apoptosis of Tc17 cells.
FIGURE 3. The soluble factor IL-2 contributes to the inhibition of Tc17 cell development in the presence of CD4+CD252T cells. Sorted CD8+T cells were cocultured with or without sorted CD4+CD252T cells at a ratio of 1:2. The cells were stimulated with anti-CD3/anti-CD28 Abs under IL-17–po-larizing conditions for 3 d. The expression of IL-17 and IFN-g on CD8+T cells was assessed by intracellular staining. (A) Anti-mouse 2, anti-mouse IL-27, or isotype control Abs were added to the culture medium at a concentration of 5 mg/ml at the beginning of the stimulation process. The numbers indicate the percentages of positive cells in each quadrant. A representative experiment is shown. (B) The means and SDs obtained from three independent experiments are shown. Significant differences were determined using Student t test, and the p values are indicated. (C) The percentages of CD8+
T cells positive for intracellular IL-17 and IFN-g following culture in the presence of 0, 0.1, 1, or 10 ng/ml exogenous recombinant mouse IL-2 are shown. The results from a representative experiment are shown. (D) The collective results derived from four independent experiments are shown. The results are represented as the mean6 SD. Statistical significance was determined using Student t test, and the p values are indicated.
FIGURE 4. CD4+CD252 T cells promote the
apo-ptosis of Tc17 cells. (A) Sorted CD8+T cells were
in-cubated with 10 mM CellTracker Orange in serum-free RPMI 1640 medium for 10 min. The cells were washed with serum-free RPMI 1640 medium before coculturing with CD4+CD252or CD4+CD25+T cells at the indi-cated ratios for 3 d in the presence of anti-CD3/anti-CD28 Abs under IL-17–polarizing conditions. The CD8+T cells were gated to evaluate their proliferative capacity as judged by the fluorescence intensity of the CellTracker Orange probe. The results from one of two representative experiments are shown. (B) Sorted CD8+
T cells were cocultured with sorted CD4+CD252 or CD4+CD25+T cells and were stimulated under IL-17– polarizing conditions for 3 d. The cells were restimu-lated with PMA–ionomycin for 4 h before staining for surface IL-17. Annexin V labeling was performed in accordance with the manufacturer’s instructions. The percentage of apoptotic cells was assessed using Annexin V staining of CD8+IL-17+T cells (top panels)
and CD8+IL-172T cells (bottom panels). A
represen-tative experiment is shown. (C) and (D) represent the results obtained from three independent apoptosis ex-periments with CD8+IL-17+and CD8+IL-172T cells, respectively. The data are expressed as the mean6 SD. Significant differences were determined using Student t test, and the p values are indicated.
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
CD4+CD25+T cells maintain the survival or expansion of Tc17 cells
To elucidate the role of CD4+T cells on Tc17 cell survival, we determined the percentages of Tc17 cells after secondary stimu-lation. FACS-purified CD8+ T cells were activated using anti-CD3/anti-CD28 Abs in the presence or absence of CD4+T cells under IL-17–polarizing conditions for 3 d. The culture super-natants were removed and were replaced with fresh medium containing IL-2 (40 U/ml) for 3 additional days before cells were restimulated with the same Abs. The expression of IL-17 and IFN-g in the CD8+
T cells was determined by intracellular staining. As shown in Fig. 6A, the majority of the CD8+T cells maintained an IL-17+IFN-g2 phenotype. In the absence of CD4+ T cells, the percentage of CD8+IL-17+T cells was 15.1%. The presence of CD4+CD25+ T cells increased this percentage significantly to 27.4%. In contrast, the presence of CD4+CD252 T cells led to
a marked reduction in the percentage of CD8+IL-17+ T cells (8.69%). The experiments were repeated three times, and these results are summarized in Fig. 6B. The results suggest that CD4+ CD252 and CD4+CD25+ T cells exert different effects on the survival of Tc17 cells.
The frequency of Tc17 cells correlates with the frequency of Treg cells in the peripheral blood of patients with HNSCC In a previous study, we found that high levels of TGF-b and IL-6 were present in the malignant effusions from cancer patients (22).
These results imply that T cell responses may favor the differ-entiation of IL-17–producing cells at tumor sites. In the current study, we used RT-PCR to examine the expression of TGF-b, IL-6, and IL-17mRNA levels in tumor tissue and in adjacent normal tissue from patients with HNSCC. Fig. 7A shows that in all of the samples tested, the TGF-b and IL-6 mRNA levels were higher in malignant tissues than in normal tissues. Consistent with our expectations, high levels of IL-17 mRNA were detected in ma-lignant tissues but not in normal tissues. The levels of cytokine mRNA were normalized to GAPDH levels, and the results are summarized in Fig. 7B. The results from this clinical study sup-port the concept that T cell differentiation is skewed toward a type 17 programming.
Next, we analyzed peripheral blood samples that were obtained from 25 healthy volunteers and 126 patients with HNSCC before any treatment. Flow cytometry was used to evaluate the frequencies of CD8+IL-17+and CD4+CD25+FOXP3+T cells in PBMCs from the untreated HNSCC patients and the healthy volunteers. The frequencies of Tc17 cells observed in the untreated patients were significantly higher than in the healthy volunteers. The represen-tative results are shown in Fig. 7C. Moreover, a significant in-crease in the frequency of Tc17 cells in PBMCs was observed as the disease progressed (Fig. 7D). A similar trend was observed for the frequency of regulatory CD4+CD25+FOXP3+T cells in PBMCs (Fig. 7E). In addition, we found a strong positive corre-lation between the frequencies of Tc17 and CD4+CD25+FOXP3+ T cells (r = 0.1968; p = 0.0154; Fig. 7F) in PBMCs from the HNSCC patients.
FIGURE 5. The expression of Bcl-2 and the active form of caspase 3 in CD8+IL-17+T cells was assessed to determine the role of CD4+CD252 T cells in promoting Tc17 apoptosis. Sorted CD8+T cells were cocultured with sorted CD4+CD252or CD4+CD25+T cells at the indicated ratios following activation with anti-CD3/anti-CD28 under IL-17–polarizing conditions for 3 d. Intracellular staining was performed 4 h after re-stimulation with PMA–ionomycin/brefeldin A. (A) CD8+
IL-17+T cells were gated, and the expression of Bcl-2 and active caspase 3 was ana-lyzed. The numbers indicate the percentages of positive cells in each quadrant. The results from a representative experiment are shown. In (B) and (C), the percentages of Bcl-2+caspase 32and Bcl-22caspase 3+cells
in CD8+IL-17+T cells, respectively, are shown. The data from three
in-dependent experiments are expressed as the mean 6 SD. Significant differences were determined using Student t test, and the p values are indicated.
FIGURE 6. CD4+CD25+ T cells contribute to the maintenance and
survival of Tc17 cells. (A) Sorted CD8+ T cells were cocultured with
sorted CD4+CD252or CD4+CD25+T cells at the indicated ratios in the
presence of anti-CD3/anti-CD28 Abs and under IL-17–polarizing con-ditions for 3 d. The cells were harvested, were washed with fresh culture medium, and were reseeded in the presence of recombinant human IL-2 (40 IU/ml). After an additional 3 d of culture, the cells were restimulated with anti-CD3/anti-CD28 Abs for 2 more days. The expression of IL-17 and IFN-g in the CD8+T cells was determined by intracellular staining. The numbers represent the percentages of positive cells in each quadrant. The results from a representative experiment are shown. (B) The collective results derived from four independent experiments are shown. The results are expressed as the mean6 SD. Statistical significances were determined using Student t test, and the p values are indicated.
1676 CTLs PARALYZED UNDER IL-17–POLARIZING CONDITIONS
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
Discussion
Th cells and Treg cells play opposite roles in the regulation of immune responses. It is accepted that Th cells facilitate CTL development and that Treg cells suppress CTL function. In this study, we show that CD4+T cells also inhibit the development of Tc17 cells (Fig. 1A, 1B). In addition, cell-to-cell contact is not an absolute requirement for the suppression of Tc17 cell differenti-ation (Fig. 1C, 1D). These findings imply that soluble factors mediate this suppressive effect. Several earlier studies have dem-onstrated that the suppressive effects of Treg cells depend on di-rect cell–cell interactions (37, 38). In marked contrast, our study reveals that the inhibition of the Tc17 cell developmental pathway is not mediated by Treg cells but by non-Treg cells. This obser-vation was confirmed by comparing the differentiation pathways of activated CD8+T cells cocultured either with Treg cells (CD4+ CD252) or with non-Treg cells (CD4+CD25+) under IL-17–po-larizing conditions (Fig. 2). Interestingly, it has been reported that human CD8+IL-17+ T cells from psoriasis skin plaques are
re-fractory to control by peripheral blood Treg cells (39). Taken to-gether, these results clearly demonstrate that the inhibition of Tc17 cell development is mediated by non-Treg cells.
Recent studies have shown that IL-2 (33) and IL-27 (34, 35) downregulate the generation of Th17 cells through STAT5- and STAT1-dependent mechanisms, respectively. We have found that Tc17 cell differentiation was restored when IL-2 but not IL-27, activity was neutralized by cytokine-specific Abs (Fig. 3A, 3B). In addition, the production of Tc17 cells was suppressed when rIL-2 was added to activated CD8+ T cells under IL-17–polarizing conditions in the absence of CD4+CD252 T cells (Fig. 3C, 3D). These results suggest that non-Treg cells produce IL-2 after their initial activation, which can limit the development of Tc17 cells. Apoptosis is a tightly regulated process that is crucial for T cell homeostasis and immune responses. The survival of T cells is modulated by the balance between proapoptotic and antiapoptotic proteins, such as active caspase 3 and Bcl-2, respectively (40). We found that the percentage of CD8+IL-17+T cells undergoing
ap-FIGURE 7. Positive correlation be-tween the frequencies of Tc17 and Treg cells in PBMCs from HNSCC patients. (A) The expression of IL-6, IL-17, and TGF-b mRNA in tumor (T) tissues compared with paired grossly normal mucosal tissue (N) from patients with HNSCC was de-termined using RT-PCR. The expres-sion of GAPDH mRNA served as an internal control. (B) The data obtained from real-time PCR normalized to GAPDH are shown. (C) The repre-sentative graphs of the frequencies of Tc17 cells observed in the untreated patients and the healthy volunteer are shown. In (D) and (E), the percentages of circulating Tc17 (CD8+IL-17+)
cells and Treg (CD4+CD25+FOXP3+)
cells in HNSCC patients compared with healthy donors were determined using flow cytometry. (F) The gener-ation of Tc17 cells correlates with the increase in Treg cells.
FIGURE 8. Illustration of the proposed tumor escape mechanism. Naive CD8+T cells in tumor-bearing hosts differentiate into Tc17 cells under IL-17–polarizing conditions. High levels of IL-2 is secreted by activated non-Treg cells (CD4+CD252)
during CD8+T cell activation, and the continued
differentiation and apoptosis of Tc17 cells are deleterious to the development of the CTL effec-tors. Thus, CTL clones are depleted. In contrast, Treg cells (CD4+CD25+) consume IL-2, which
may maintain or promote Tc17 cells differentia-tion. These Tc17 cells are disarmament CTLs that may endow a proinflammatory effect, but they at-tenuate cytotoxicity, and the outcome favors tumor growth.
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
optosis increased in the presence of CD4+CD252T cells. How-ever, this percentage of apoptotic CD8+IL-172 T cells was not altered in the presence of CD4+T cells (Fig. 4B–D). In addition, we found that CD4+CD252T cells, but not CD4+CD25+T cells, upregulated the expression of active caspase 3 and downregulated the expression of Bcl-2 in CD8+IL-17+T cells (Fig. 5). This ob-servation demonstrates that CD4+CD252 T cells promote apo-ptosis in CD8+IL-17+T cells. Therefore, CD8+ T cells are de-pleted under IL-17–polarizing conditions, whereas CD8+T cells are persistently activated in the presence of CD4+CD25-T cells. In contrast, CD4+CD25+T cells upregulated the expression of Bcl-2 but not the expression of active caspase 3 in CD8+IL-17+T cells (Fig. 5). These results suggest that Treg cells maintain the survival and expansion of Tc17 cells. The significant increase in the per-centage of Tc17 cells observed after the immune reactivation in the presence of CD4+CD25+T cells is consistent with this finding (Fig. 6). It has been demonstrated that Treg cells deplete IL-2 from immunological sites through two mechanisms, the suppression of IL-2 production by activated T cells (38) or the consumption of IL-2 to limit the activated T cell exposure to IL-2 (41, 42). Our results explain why Treg cells do not inhibit the development of Tc17 cells but support their survival and expansion.
In 2007, we identified IL-17–producing CD8+T cells with a noncytotoxic phenotype (17). Subsequently, several reports (18– 21) have characterized murine IL-17–secreting CD8+ T cells. These reports demonstrate that IL-17–polarizing conditions en-hance retinoic acid receptor-related orphan receptor-gt and reti-noic acid receptor-related orphan receptor-a expression and sup-press Eomes exsup-pression, thereby augmenting IL-17 production by CD8+T cells. The loss of cytotoxic function associated with the production of IL-17 under IL-17–polarizing conditions results in a marked reduction of granzyme B and perforin expression. In agreement with other studies in mice, Kuang et al. (43) showed that IL-17–producing CD8+ T cells accumulate in human he-patocellular carcinomas. These IL-17–producing CD8+ T cells exhibited the reduced expression of granzyme B, perforin, and CD107a when compared with IFN-g–producing CD8+T cells (43). These results also confirm that Tc17 cells are unable to me-diate significant cytotoxicity. In the current study, regardless of whether CD8+T cells are activated in the presence of Treg cells (CD4+CD25+) or non-Treg cells (CD4+CD252) under IL-17–po-larizing conditions, IL-17–secreting CD8+T cells do not produce IFN-g (Figs. 1, 3, 6), and they express low levels of granzyme B with little cytotoxicity (Fig. 2). Therefore, activated CD8+T cells maintain a noncytotoxic phenotype or an attenuated cytotoxic potential under IL-17–polarizing conditions.
However, the adoptive transfer of tumor-specific Tc17 cells has been shown to control tumor growth (44). The recruitment of neutrophils or other effector cells may mediate the Tc17 cell mechanism that controls tumor growth. Importantly, the effec-tiveness of Tc17 cells to control tumor growth is∼50-fold less than that of Tc1 cells. These results also suggest that Tc17 cells are not effective for the control of tumor growth. The role of IL-17 and IL-17–producing T cells in malignancy is currently under debate (29–32). Currently, very little is known about the nature and regulation of Tc17 cells in cancer development. Recently, accumulating evidence has revealed that Tc17 cells are frequently enriched in mouse tumor models (28) and in various types of human cancer (28, 43, 45, 46). In this study, we show that the frequency of Tc17 cells was markedly increased in the peripheral blood of patients with HNSCC. Patients with the advanced-stage disease exhibited significantly higher frequencies of Tc17 cells compared with patients with the early-stage disease (Fig. 7D). It is known that Treg cells are more abundant in the peripheral blood of
cancer-bearing patients compared with healthy subjects. We have reconfirmed that the number of CD4+CD25+FOXP3+Treg cells in the peripheral blood of patients with HNSCC is significantly in-creased (Fig. 7E). Moreover, we have shown that the numbers of Tc17 cells and CD4+CD25+FOXP3+Treg cells increase simulta-neously during disease progression (Fig. 7F). Consistent with our findings, Treg cells maintain Tc17 cell survival (Fig. 6) but do not limit Tc17 cell development (Fig. 2). It may be argued that the variation of elicited Tc17 population occurred in independent studies. Such variation did not interfere the conclusion that the presence of CD4+CD252, but not the CD4+CD25+cells, limits Tc17 skewing.
In a previous study, we detected very high levels of TGF-b and IL-6 in the malignant effusions of cancer patients (22). In addition, it has been demonstrated that tumor-activated monocytes promote the expansion of Tc17 cells (43). In this study, we show that high levels of TGF-b and IL-6 mRNA are detected in tumor tissues from patients with HNSCC (Fig. 7A, 7B). These results indicate that the tumor microenvironment favors Tc17 cell development. In conclusion, a possible tumor escape mechanism is depicted in Fig. 8. The CTLs in tumor-bearing hosts are biased to an IL-17–se-creting phenotype. In this scenario, the continued differentiation and apoptosis of Tc17 cells is deleterious to the development of CTLs in the presence of IL-2 produced by non-Treg cells during their initial activation. Thus, CTL clones are dampened by de-pletion. Contrarily, Treg cells deplete IL-2, which favors rather than inhibits Tc17 cell differentiation. However, Tc17 cells are disarmament CTLs (18–21) that may be endowed with a proin-flammatory property but an attenuated cytotoxicity. The outcome will encourage tumor growth. Our findings open new avenues for understanding the regulation of Tc17 cell development and tumor escape mechanisms.
Acknowledgments
We thank Shian-Yan I and Ya-Ken Chen for assistance in collecting the clin-ical samples and the written informed consents. We also thank Dr. Michel Klein for critically reviewing the manuscript and for providing suggestions.
Disclosures
The authors have no financial conflicts of interest.
References
1. Husmann, L. A., and M. J. Bevan. 1988. Cooperation between helper T cells and cytotoxic T lymphocyte precursors. Ann. N. Y. Acad. Sci. 532: 158–169. 2. Keene, J. A., and J. Forman. 1982. Helper activity is required for the in vivo
generation of cytotoxic T lymphocytes. J. Exp. Med. 155: 768–782. 3. Bevan, M. J. 2004. Helping the CD8+T-cell response. Nat. Rev. Immunol. 4:
595–602.
4. Castellino, F., and R. N. Germain. 2006. Cooperation between CD4+and CD8+
T cells: when, where, and how. Annu. Rev. Immunol. 24: 519–540.
5. Williams, M. A., and M. J. Bevan. 2007. Effector and memory CTL differen-tiation. Annu. Rev. Immunol. 25: 171–192.
6. Sakaguchi, S., N. Sakaguchi, M. Asano, M. Itoh, and M. Toda. 1995. Immu-nologic self-tolerance maintained by activated T cells expressing IL-2 receptor a-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 155: 1151–1164.
7. Fontenot, J. D., M. A. Gavin, and A. Y. Rudensky. 2003. Foxp3 programs the development and function of CD4+CD25+regulatory T cells. Nat. Immunol. 4:
330–336.
8. Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T cell de-velopment by the transcription factor Foxp3. Science 299: 1057–1061. 9. Suvas, S., U. Kumaraguru, C. D. Pack, S. Lee, and B. T. Rouse. 2003. CD4+
CD25+ T cells regulate virus-specific primary and memory CD8+ T cell
responses. J. Exp. Med. 198: 889–901.
10. Onizuka, S., I. Tawara, J. Shimizu, S. Sakaguchi, T. Fujita, and E. Nakayama. 1999. Tumor rejection by in vivo administration of anti-CD25 (interleukin-2 receptor a) monoclonal antibody. Cancer Res. 59: 3128–3133.
11. Shimizu, J., S. Yamazaki, and S. Sakaguchi. 1999. Induction of tumor immunity by removing CD25+CD4+T cells: a common basis between tumor immunity and
autoimmunity. J. Immunol. 163: 5211–5218.
1678 CTLs PARALYZED UNDER IL-17–POLARIZING CONDITIONS
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/
12. Chen, M. L., M. J. Pittet, L. Gorelik, R. A. Flavell, R. Weissleder, H. von Boehmer, and K. Khazaie. 2005. Regulatory T cells suppress tumor-specific CD8 T cell cytotoxicity through TGF-b signals in vivo. Proc. Natl. Acad. Sci. USA 102: 419–424.
13. Mempel, T. R., M. J. Pittet, K. Khazaie, W. Weninger, R. Weissleder, H. von Boehmer, and U. H. von Andrian. 2006. Regulatory T cells reversibly suppress cytotoxic T cell function independent of effector differentiation. Immunity 25: 129–141.
14. Chappert, P., M. Leboeuf, P. Rameau, M. Lalfer, S. Desbois, R. S. Liblau, O. Danos, J. M. Davoust, and D. A. Gross. 2010. Antigen-specific Treg impair CD8+T-cell priming by blocking early T-cell expansion. Eur. J. Immunol. 40: 339–350.
15. Heit, A., F. Gebhardt, K. Lahl, M. Neuenhahn, F. Schmitz, F. Anderl, H. Wagner, T. Sparwasser, D. H. Busch, and K. Kastenmu¨ller. 2008. Circumvention of regulatory CD4+T cell activity during cross-priming strongly enhances T cell-mediated immunity. Eur. J. Immunol. 38: 1585–1597.
16. McNally, A., G. R. Hill, T. Sparwasser, R. Thomas, and R. J. Steptoe. 2011. CD4+CD25+regulatory T cells control CD8+T-cell effector differentiation by
modulating IL-2 homeostasis. Proc. Natl. Acad. Sci. USA 108: 7529–7534. 17. Liu, S.-J., J.-P. Tsai, C.-R. Shen, Y.-P. Sher, C.-L. Hsieh, Y.-C. Yeh, A.-H. Chou,
S.-R. Chang, K.-N. Hsiao, F.-W. Yu, and H.-W. Chen. 2007. Induction of a dis-tinct CD8 Tnc17 subset by transforming growth factor-b and interleukin-6. J. Leukoc. Biol. 82: 354–360.
18. Huber, M., S. Heink, H. Grothe, A. Guralnik, K. Reinhard, K. Elflein, T. Hu¨nig, H. W. Mittru¨cker, A. Bru¨stle, T. Kamradt, and M. Lohoff. 2009. A Th17-like developmental process leads to CD8+Tc17 cells with reduced cytotoxic activity.
Eur. J. Immunol. 39: 1716–1725.
19. Yen, H. R., T. J. Harris, S. Wada, J. F. Grosso, D. Getnet, M. V. Goldberg, K. L. Liang, T. C. Bruno, K. J. Pyle, S. L. Chan, et al. 2009. Tc17 CD8 T cells: functional plasticity and subset diversity. J. Immunol. 183: 7161–7168. 20. Hamada, H., Mde. L. Garcia-Hernandez, J. B. Reome, S. K. Misra, T. M. Strutt,
K. K. McKinstry, A. M. Cooper, S. L. Swain, and R. W. Dutton. 2009. Tc17, a unique subset of CD8 T cells that can protect against lethal influenza challenge. J. Immunol. 182: 3469–3481.
21. Curtis, M. M., S. S. Way, and C. B. Wilson. 2009. IL-23 promotes the production of IL-17 by antigen-specific CD8 T cells in the absence of IL-12 and type-I interferons. J. Immunol. 183: 381–387.
22. Tsai, J.-P., H.-W. Chen, M.-L. Cheng, H.-K. Liu, Y.-P. Lee, C. L. Hsieh, K.-T. Luh, C.-W. Wu, L.-H. Hsu, T.-Y. Chao, et al. 2005. Analysis of host versus tumor interaction in cancer patients: opposing role of transforming growth factor-b1 and interleukin-6 in the development of in situ tumor immu-nity. Immunobiology 210: 661–671.
23. Bettelli, E., Y. Carrier, W. Gao, T. Korn, T. B. Strom, M. Oukka, H. L. Weiner, and V. K. Kuchroo. 2006. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441: 235–238. 24. Mangan, P. R., L. E. Harrington, D. B. O’Quinn, W. S. Helms, D. C. Bullard,
C. O. Elson, R. D. Hatton, S. M. Wahl, T. R. Schoeb, and C. T. Weaver. 2006. Transforming growth factor-b induces development of the T(H)17 lineage. Nature 441: 231–234.
25. Veldhoen, M., R. J. Hocking, C. J. Atkins, R. M. Locksley, and B. Stockinger. 2006. TGFb in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17‑producing T cells. Immunity 24: 179–189.
26. Sheibanie, A. F., T. Khayrullina, F. F. Safadi, and D. Ganea. 2007. Prostaglandin E2 exacerbates collagen-induced arthritis in mice through the inflammatory
interleukin-23/interleukin-17 axis. Arthritis Rheum. 56: 2608–2619.
27. Sheibanie, A. F., I. Tadmori, H. Jing, E. Vassiliou, and D. Ganea. 2004. Pros-taglandin E2induces IL-23 production in bone marrow-derived dendritic cells.
FASEB J. 18: 1318–1320.
28. Kryczek, I., S. Wei, L. Zou, S. Altuwaijri, W. Szeliga, J. Kolls, A. Chang, and W. Zou. 2007. Cutting edge: Th17 and regulatory T cell dynamics and the regulation by IL-2 in the tumor microenvironment. J. Immunol. 178: 6730–6733. 29. Murugaiyan, G., and B. Saha. 2009. Protumor vs antitumor functions of IL-17. J.
Immunol. 183: 4169–4175.
30. Maniati, E., R. Soper, and T. Hagemann. 2010. Up for mischief? IL-17/Th17 in the tumour microenvironment. Oncogene 29: 5653–5662.
31. Wilke, C. M., I. Kryczek, S. Wei, E. Zhao, K. Wu, G. Wang, and W. Zou. 2011. Th17 cells in cancer: help or hindrance? Carcinogenesis 32: 643–649. 32. Ji, Y., and W. Zhang. 2010. Th17 cells: positive or negative role in tumor?
Cancer Immunol. Immunother. 59: 979–987.
33. Laurence, A., C. M. Tato, T. S. Davidson, Y. Kanno, Z. Chen, Z. Yao, R. B. Blank, F. Meylan, R. Siegel, L. Hennighausen, et al. 2007. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity 26: 371– 381.
34. Amadi-Obi, A., C. R. Yu, X. Liu, R. M. Mahdi, G. L. Clarke, R. B. Nussenblatt, I. Gery, Y. S. Lee, and C. E. Egwuagu. 2007. TH17 cells contribute to uveitis and scleritis and are expanded by IL-2 and inhibited by IL-27/STAT1. Nat. Med. 13: 711–718.
35. Neufert, C., C. Becker, S. Wirtz, M. C. Fantini, B. Weigmann, P. R. Galle, and M. F. Neurath. 2007. IL-27 controls the development of inducible regulatory T cells and Th17 cells via differential effects on STAT1. Eur. J. Immunol. 37: 1809–1816.
36. Brucklacher-Waldert, V., K. Steinbach, M. Lioznov, M. Kolster, C. Ho¨lscher, and E. Tolosa. 2009. Phenotypical characterization of human Th17 cells unambig-uously identified by surface IL-17A expression. J. Immunol. 183: 5494–5501. 37. Read, S., S. Mauze, C. Asseman, A. Bean, R. Coffman, and F. Powrie. 1998.
CD38+CD45RBlowCD4+T cells: a population of T cells with immune regulatory
activities in vitro. Eur. J. Immunol. 28: 3435–3447.
38. Thornton, A. M., and E. M. Shevach. 1998. CD4+CD25+ immunoregulatory
T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J. Exp. Med. 188: 287–296.
39. Ortega, C., S. Ferna´ndez-A, J. M. Carrillo, P. Romero, I. J. Molina, J. C. Moreno, and M. Santamarı´a. 2009. IL-17‑producing CD8+T lymphocytes from psoriasis skin plaques are cytotoxic effector cells that secrete Th17-related cytokines. J. Leukoc. Biol. 86: 435–443.
40. Krammer, P. H., R. Arnold, and I. N. Lavrik. 2007. Life and death in peripheral T cells. Nat. Rev. Immunol. 7: 532–542.
41. Barthlott, T., H. Moncrieffe, M. Veldhoen, C. J. Atkins, J. Christensen, A. O’Garra, and B. Stockinger. 2005. CD25+CD4+T cells compete with naive
CD4+T cells for IL-2 and exploit it for the induction of IL-10 production. Int.
Immunol. 17: 279–288.
42. Pandiyan, P., L. Zheng, S. Ishihara, J. Reed, and M. J. Lenardo. 2007. CD4+
CD25+Foxp3+regulatory T cells induce cytokine deprivation-mediated apoptosis
of effector CD4+T cells. Nat. Immunol. 8: 1353–1362.
43. Kuang, D. M., C. Peng, Q. Zhao, Y. Wu, L. Y. Zhu, J. Wang, X. Y. Yin, L. Li, and L. Zheng. 2010. Tumor-activated monocytes promote expansion of IL-17 ‑pro-ducing CD8+T cells in hepatocellular carcinoma patients. J. Immunol. 185:
1544–1549.
44. Garcia-Hernandez, Mde. L., H. Hamada, J. B. Reome, S. K. Misra, M. P. Tighe, and R. W. Dutton. 2010. Adoptive transfer of tumor-specific Tc17 effector T cells controls the growth of B16 melanoma in mice. J. Immunol. 184: 4215–4227. 45. Chen, X., J. Wan, J. Liu, W. Xie, X. Diao, J. Xu, B. Zhu, and Z. Chen. 2010.
Increased IL-17‑producing cells correlate with poor survival and lymphangio-genesis in NSCLC patients. Lung Cancer 69: 348–354.
46. Wu, C., S. Wang, F. Wang, Q. Chen, S. Peng, Y. Zhang, J. Qian, J. Jin, and H. Xu. 2009. Increased frequencies of T helper type 17 cells in the peripheral blood of patients with acute myeloid leukaemia. Clin. Exp. Immunol. 158: 199–204.
at National Health Research Institutes on August 5, 2012
http://jimmunol.org/