Denbinobin suppresses breast cancer metastasis through the inhibition
of Src-mediated signaling pathways
☆
Pei-Hsuan Chen
a, Chieh-Yu Peng
b,1, Hui-Chen Pai
b, Che-Ming Teng
b, Chien-Chih Chen
c, Chia-Ron Yang
a,⁎
aSchool of Pharmacy, College of Medicine, National Taiwan University, Taipei, Taiwan bInstitute of Pharmacology, College of Medicine, National Taiwan University, Taipei, Taiwan
c
Department of Biotechnology, Hungkuang University, Taichung, Taiwan
Received 10 March 2010; received in revised form 14 May 2010; accepted 4 June 2010
Abstract
Denbinobin (5-hydroxy-3,7-dimethoxy- 1,4-phenanthraquinone), a biologically active chemical isolated from Ephemerantha lonchophylla, has been demonstrated to display anti-cancer activity. Breast cancer is the leading cause of female mortality, and the high mortality is mainly attributable to metastasis. Src kinase activity is elevated in many human cancers, including breast cancer, and is often associated with aggressive disease. In the present study, we examined the anti-metastatic effects of denbinobin through decreasing Src kinase activity in human and mouse breast cancer cells. Denbinobin caused significant block of Src kinase activity in both human and mouse breast cancer cells. Moreover, phosphorylation of the signaling molecules focal adhesion kinase, Crk-associated substrate and paxillin downstream of Src was also inhibited by denbinobin. Furthermore, denbinobin inhibited the in vitro migration, invasion and in vivo metastasis of breast cancers in a mouse metastatic model. The denbinobin-treated group showed a significant reduction in tumor metastasis, orthrotopic tumor volume, and spleen enlargement compared to the control group. In addition, transfection of breast cancer cells with a plasmid coding for a constitutively active Src prevented the denbinobin-mediated phosphorylation of Src and downstream molecules and cell migration. Our findings provide evidences that denbinobin inhibits Src-mediated signaling pathways involved in controlling breast cancer migration and metastasis, suggesting that it has therapeutic potential in breast cancer treatment.
© 2010 Elsevier Inc. All rights reserved.
Keywords: Denbinobin; Natural products; Breast cancer; Src; Migration; Metastasis
1. Introduction
Invasion and metastasis are the main clinical phenomena that
distinguish malignant from benign tumors and are the leading causes
of death in cancer patients. Metastasis is a complicated process
involving the degradation of the basement membrane, invasion of the
stroma, adhesion, angiogenesis, cell proliferation and migration
[1]
.
Many factors in tumor metastasis need further investigation, as a
variety of modulators may be involved
[2]
, and current clinical
methods cannot accurately identify which patients will develop
metastasis
[3]
. In addition, there are few treatments that can
significantly suppress tumor metastasis, so the development of
more effective therapeutic agents is a high priority.
Src family kinases comprise a subclass of membrane-associated
non-receptor tyrosine kinases involved in a variety of cellular signal
transduction pathways. Src family kinases include Src, Yes, Fyn, Lyn,
Hck, Blk, Brk, Fgr, Frk, Srm and Yrk
[4]
. Although most Src family
members are expressed primarily in cells of hematopoietic origin, Src
displays a more ubiquitous pattern of expression, with high levels in
some epithelial tissue
[4]
. Under normal physiological condition,
90-95% of Src is in an inactive conformation
[5]
. However, activated Src is
frequently reported in human cancers, including colon
adenocarci-noma
[6]
, breast cancer
[7]
and pancreatic cancer
[8]
. In the case of
breast cancer, increased Src activity is believed to play an important
role in development and progression
[4,5]
. When acted upon by
upstream signals, such as growth factors or cytoplasmic proteins, Src
undergoes a conformational change resulting in activation of its
kinase activity
[9]
. Activated Src coordinates multiple signaling
pathways involved in tumor progression, such as proliferation,
survival, motility, angiogenesis, and invasion
[10]
. In addition, its
ability to promote tumor cell invasion can lead to the development of
tumor metastasis
[11]
. A recent study using Src
−/−mice
demonstrat-ed that abolishing Src expression/activity decreases tumor cell
extravasation and subsequently decreases experimental metastasis
[12]
. Thus, Src is a potential therapeutic target of human cancer,
including breast cancer.
Increased Src activity can be caused by increased transcription or
by deregulation due to overexpression of upstream growth factor
receptors, such as epidermal growth factor receptor or
platelet-derived growth factor receptor, or by cytoplasmic proteins, such as
Journal of Nutritional Biochemistry xx (2010) xxx
–xxx
☆ Grant support: National Science Council of Taiwan
(NSC97-2320-B-002-019-MY3).
⁎ Corresponding author. Tel: +886 2 23952060; fax: +886 2 23919098. E-mail address:[email protected](C.-R. Yang).
1 Co-first author: contributed equally to this work.
0955-2863/$ - see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.jnutbio.2010.06.004
ephrins and integrin
[13,14]
. Src kinases phosphorylate tyrosyl
residues of critical cellular substrates, resulting in the activation of
oncogenic signal transduction pathways
[15]
. Focal adhesion kinase
(FAK), one such substrate of Src, plays an important role in integrin
signaling and is highly expressed in many tumor cells
[16]
. A study
using an inducible dominant-negative Src demonstrated that Src
suppression significantly reduces the migration, attachment, and
spreading of MCF-7 cells through changes in FAK activation and the
interaction of CRK-associated substrate (p130Cas) with FAK
[17]
.
The Src-FAK signaling complex recruits and phosphorylates a number
of signaling proteins and is involved in adhesion regulation and the
motile and invasive phenotype
[5]
.
Denbinobin (5-hydroxy-3,7-dimethoxy- 1,4-phenanthraquinone)
is a biologically active chemical isolated from Ephemerantha
loncho-phylla. Previous studies have shown that it acts as an antioxidant and
anti-tumorigenesis agent
[18
–23]
. In this study, we evaluated its
effects on Src kinase inhibition and breast cancer cell migration in
vitro and on tumor metastasis in vivo.
2. Materials and methods 2.1. Materials
Denbinobin was extracted and purified by one of our colleagues (Dr. Chien-Chih Chen) to a purity of greater than 98%, as shown by high-performance liquid chromatography and nuclear magnetic resonance[19]. Rabbit polyclonal antibodies against human phospho-FAK (Tyr576/577), phospho-FAK (Tyr925), phospho-p130Cas (Tyr410), and phospho-paxillin (Tyr118) and monoclonal antibodies against human Src were purchased from Cell Signaling Technology (Danvers, MA, USA). Mouse monoclonal anti-human phospho-Src (Tyr416) antibody was purchased from Millipore (Billerica, MA, USA). Rabbit polyclonal anti-human FAK, p130Cas, or Glyceraldehyde 3-phosphate dehydrogenase antibodies and protein A/G-PLUS agarose were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Horse radish peroxidase (HRP)-or flu(HRP)-orescein isothiocyanate (FITC)-conjugated goat anti-mouse (HRP)-or anti-rabbit IgG antibodies were obtained from Jackson ImmunoResearch (Cambridgeshire, UK). Constitutively active c-Src (CA-Src) plasmid, which contains a tyrosine-to-phenylal-anine substitution (Y529F), was obtained from Upstate (Lake Placid, NY, USA), and the pGL4.74[hRluc/TK] vector was obtained from Promega (Madison, WI, USA). Lipofectamine 2000 reagent was purchased from Gibco Laboratories Inc. (Palo Alto, CA, USA). The blind well chemotaxis chamber was obtained from Neuro Probe (Gaithersburg, MD, USA) and the Nuclepore Track-etch polycarbonate membrane from Whatman (Maidstone, Kent, UK). Recombinant human EGF was purchased from PeproTech Asia (Rehovot, Israel). The protein tyrosine kinase assay kit and all other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
2.2. Cell culture
Mouse breast tumor 4T1-Luc cell lines cotransfected with the firefly luminance-containing vector and selected in G418 were kindly provided by Dr. Min-Liang Kuo (Institute of Pathology, College of Medicine, National Taiwan University) and human breast adenocarcinoma MDA-MB-231 cells were obtained from the American Type Culture Collection (Manassas, VA, USA). Both were cultured in the media recom-mended by the supplier (high glucose DMEM medium for 4T1-Luc cells and RPMI-1640 medium for MDA-MB-231 cells) supplemented with 10% (v/v) fetal bovine serum (FBS; Invitrogen Life Technologies, Carlsbad, CA, USA), 100 U/ml of penicillin, and 100 μg/ml of streptomycin (Biological Industries, Kibbutz Beit Haemek, Israel) at 37°C in a humidified atmosphere of 5% CO2in air.
2.3. Cell viability assays Cells (1×104
) in 100μl of medium in 96-well plates were incubated with vehicle or test compound for 48 h, then 25μl of 1 mg/ml of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) was added, and the plates incubated at 37°C for 2 h. The cells were then pelleted and lysed in 100μl of dimethyl sulfoxide and the absorbance at 550 nm measured on a microplate reader.
2.4. Immunoblot analysis Cells (1×106
) were lysed in a lysis buffer and whole-cell extracts (60μg) were electrophoresed on 10% sodium dodecyl sulfate-polyacrylamide gel electrohporesis and blotted onto nitrocellular membranes. Immunoblot detection was performed with the corresponding antibodies using an ECL detection kit and exposure to photographic film.
2.5. Src kinase assays in vitro
Cells were lysed in 50 mM HEPES, pH 7.4, containing 0.1% Triton X-100, 10% glycerol, 1 mM dithiothreitol, 1 mM sodium vanadate, 1 mM benzamidine, 10μg/ml of aprotinin, 10μg/ml of leupeptin, and 2 μg/ml of pepstatin A and the lysates were clarified by centrifugation at 17,000 g for 30 min at 4°C, then active Src was precipitated using 1μg of anti-c-Src antibody and protein A/G-agarose beads at 4°C overnight. The precipitated beads were washed three times with 1 ml of ice-cold cell lysis buffer and used in the kinase reaction. The Src kinase assay was performed using the supplier's protocol (Sigma-Aldrich). The absorbance was measured at 492 nm.
2.6. Transient transfection with a constitutively activated c-Src mutant
1×106cells were seeded in 6-well plates in 1 ml of serum-free medium one day
before transfection. Following the manufacturer's protocol, 10μl of Lipofectamine 2000 (Invitrogen) was mixed for 5 min with 50μl of Opti-MEMI reduced serum medium, then 10μg of CA-Src (Y529F) plasmid DNA and 10μg of pEGFP-N1 plasmid in 50 μl of Opti-MEMI reduced serum medium were added. The mixture was incubated for 20 min at room temperature, then was added to the cells, and the mixture incubated for 24 h at 37°C in a humidified atmosphere of 5% CO2in air. Transfection efficiency, determined
by fluorescence microscopy, wasN60% in all experiments. 2.7. Cell migration and invasion assays
The migration assays were measured using a blind well chemotaxis chamber. Cells (1×105
in 100μl of culture medium containing 0.5% FBS) or 100 μl of culture medium containing 10% FBS was added to the top or bottom wells of the chamber, respectively. The chambers were incubated for 24 h at 37°C in a humidified atmosphere of 5% CO2in air, then non-migrated cells on the top of the filters were
wiped off with cotton swabs and the migrated cells attached to the bottom of the filters were fixed and stained with 1% crystal violet, counted in microscope and the absorbance at 550 nm measured. In invasion assays, the filter was coated with Matrigel (BD Biosciences, Bedford, MA) at a concentration of 125 μg/cm2
, then followed protocols were performed as above described.
2.8. Metastatic assays in vivo 4T1-Luc cells (2×105
in 0.1 ml of medium) were injected into the left lower mammary fat pad of 5-week-old female BALB/c mice. Tumor growth was measured weekly and the volume (mm3
) was determined by measuring the largest diameter (l) and smallest diameter (s), using the equation volume=0.5 ls2. The animals were also
weighed weekly. After the orthotopic tumor volume reached 50 mm3(14 days after
tumor cell injection), the mice were either left untreated or were injected intraperitoneally with either paclitaxel (20 mg/kg) every 4 days or denbinobin (10 mg/kg) 3 times a week. An IVIS 200 biophotonic imager (Xenogen) was used to monitor metastasis of the breast carcinoma cells. Briefly, the animals were anesthetized by isoflurane inhalation and injected intraperitoneally with 100μl of 150 mg/ml of D-luciferin (Xenogen) in phosphate-buffered saline (PBS), and bioluminescence imaging with a CCD camera (IVIS, Xenogen) was initiated 10 min after injection. At the experimental endpoint of day 40 after tumor injection, when lung metastasis was observed in the control group by bioluminescence imagery, the animals were sacrificed and imaged within 15 min after injection. All animal studies were performed using protocols approved by the Institutional Animal Care and Use Committee of the College of Medicine, National Taiwan University.
2.9. Immunocytochemistry and microscopy
Cells or removed orthotopic tumors were fixed in 4% paraformaldehyde in PBS. After fixation, the orthotopic tumors were embedded in paraffin and cut into 5-μm sections. Nonspecific binding sites in the fixed cultures or tumor slices were blocked by overnight incubation with 5% bovine serum albumin (BSA) in PBS overnight, then the samples were incubated with primary antibodies (1:100) in 0.5% BSA for 60 min at room temperature. After 3×10-min washes in PBS, the cells or tumor sections were stained for another 60 min with FITC-conjugated goat anti-mouse or anti-rabbit IgG Fig. 1. Denbinobin inhibits the kinase activity of Src and its downstream signaling pathways in vitro. In (A) and (B), 1×106
4T1-Luc cells (1×106
) (A) or MDA-MB-231 cells (B) were incubated with EGF (100 ng/ml) for 0–60 min, then the cells were harvested and whole cell extracts prepared for Western blot analysis for the indicated proteins. In (C) and (D), 4T1-Luc cells (1×106
) (C) or MDA-MB-231 cells (1×106
) (D) were incubated with 10μg empty vector (EV) or constitutively active Src plasmid (CA-Src) for 24 h, denbinobin (1,3,10μM) for 30 min, and 100 ng/ml EGF for 1h as indicated, then whole cell extracts were prepared for Western blot analysis for the indicated proteins. Viability of 4T1-Luc cells (E) or MDA-MB-231 cells (F) treated with 0.3-10μM denbinobin for 6 or 12 h compared to that of the control group estimated using the MTT assay. The data are the mean±S.E.M. for four separate experiments.
antibodies (1:100 dilution in PBS) then were viewed and photographed under a Leica TCS SP5 confocal laser-scanning microscope using appropriate fluorescence filters. 2.10. Data analysis
The data are expressed as the mean±S.E.M. and were analyzed statistically using one-way analysis of variance (ANOVA). When ANOVA showed significant differences between groups, Tukey post hoc test was used to determine the specific pairs of groups showing statistically significant differences. A P value of less than .05 was considered statistically significant.
3. Results
3.1. Denbinobin inhibits the kinase activity of Src and its downstream
signaling pathways in vitro
Src-mediated downstream signaling pathways involving FAK,
p130Cas, and paxillin are reported to play an essential role in tumor
cell migration and metastasis
[24
–30]
. EGF treatment induced Src
phosphorylation at Tyr416 within 5 min and this was maintained for
at least 1 h in both 4T1-Luc (
Fig. 1
A) and MDA-MB-231 (
Fig. 1
B)
breast cancer cells. Furthermore, following Src activation, FAK was
phosphorylated at Src-dependent phosphorylation sites (Tyr576 and
Tyr925) and p130Cas was phosphorylated at Tyr410 (
Fig. 1
A).
Denbinobin treatment for 30 min not only significantly reduced Src
Tyr416 phosphorylation but also reduced phosphorylation of FAK
Tyr576 and Tyr925, p130Cas Tyr410, and paxillin Tyr118 in a
concentration-dependent manner in both cell types (
Fig. 1
C, D).
This inhibition were not due to decreased protein levels, since total
Src and FAK levels were unchanged and none of the treatments had
any significant effect on cell viability at 6 or 12 h, assessed using the
MTT assay (
Fig. 1
E, F). In order to directly examine the role of
denbinobin in Src activation, both types of breast cancer cells were
transfected for 24 h with 10
μg of plasmid CA-Src coding for a
constitutively activate Src and phosphorylation of Src, FAK, p130Cas,
and paxillin was found to be significantly increased, reversing the
inhibition by denbinobin (
Fig. 1
C, D). These data clearly demonstrate
that denbinobin inhibits the phosphorylation of Src and associated
downstream molecules.
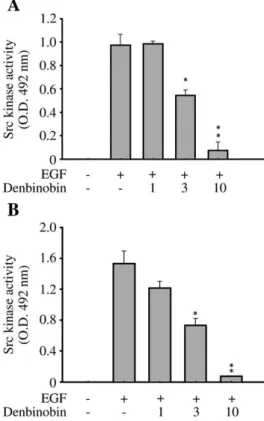
We next asked whether denbinobin influenced Src kinase. To
address this question, we treated both cell lines with denbinobin
and evaluated the denbinobin-mediated Src kinase inhibitory effect
using the Src kinase assay as described in the Materials and
Methods section. As shown in
Fig. 2
, 10
μM of denbinobin resulted
in marked inhibition of Src kinase to less than 20% of control levels
in both cell lines.
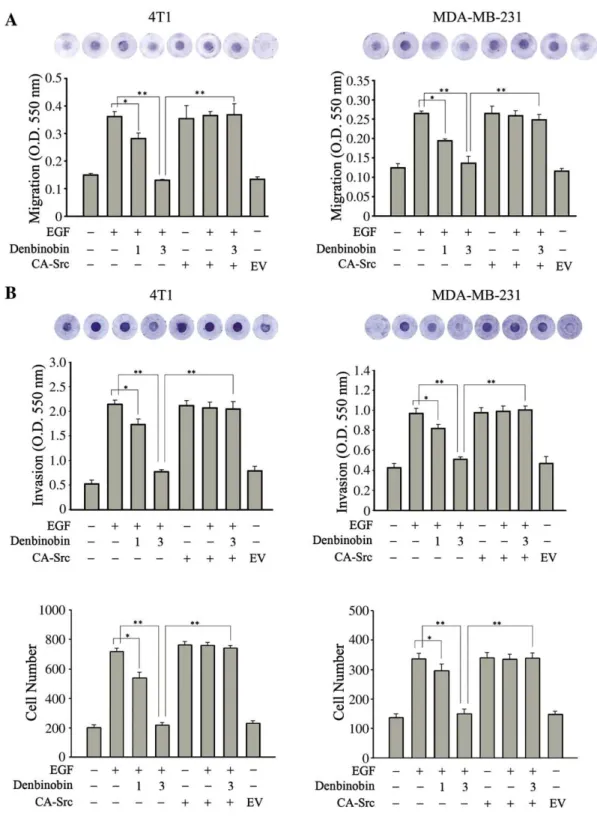
3.2. Denbinobin inhibits the migration and invasion of breast cancer
cells in vitro
Since Src kinase plays an important role in cell migration, we next
examined the effect of denbinobin on cell migration and invasion in a
chemotaxis chamber assay. MDA-MB-231 cells and 4T1-Luc cells are
highly invasive breast cancer cell lines. EGF treatment increased the
migration and invasion of both cell types, and these effects were
significantly inhibited by 3
μM denbinobin to less than 35% of the
levels in the EGF control group, respectively (
Fig. 3
A, B). Furthermore,
after transfection with CA-Src, the constitutively active Src increased
the migratory and invasion motility of cancer cells with or without
EGF treatment and prevented inhibition by denbinobin (3
μM) (
Fig.
3
A, B). These results confirmed our previous argument that
denbinobin exerts its anti-migratory effect via Src inactivation in
breast cancer cells.
In order to further characterize the molecules involved in the
inhibitory effect of denbinobin, we labeled cells on coverslips with
FITC-conjugated antibodies against phosphorylated Src, FAK, or
paxillin and examined the results on a confocal laser-scanning
microscope. As shown in
Fig. 4
, p-Src was found to be localized in
the cytoplasm in cells with or without treatment with 3
μM
denbinobin for 1 h. However, the denbinobin-treated cells were
rounder in shape and the intensity of the p-Src labeling was
significantly reduced (
Fig. 4
A, B). Likewise, labeling for FAK
phosphorylated at Tyr576/577 and paxillin phosphorylated at
Tyr118 was decreased by the same treatment (
Fig. 4
A, B), supporting
our results obtained by immunoblot analysis (
Fig. 1
) and Src kinase
assay (
Fig. 2
). These findings clearly demonstrate that denbinobin
inhibits breast cancer cell migration by inhibiting Src-FAK signaling.
3.3. Denbinobin inhibits breast cancer cell metastasis in vivo
We next used a metastatic animal model to evaluate whether
denbinobin inhibited breast cancer cell metastasis. When introduced
orthotopically, the 4T1-Luc mammary carcinoma cells can
metasta-size to several organs, including the lung, liver, and spleen. We used
an IVIS biophotonic imager to monitor the luminance of 4T1-Luc
metastasis and an end point of 40 days, when all control mice showed
lung metastasis. At 40 days after injecting BALB/c mice with 2×10
5mouse breast cancer cells, strong luminance was observed in the lung
(
Fig. 5
A). Luminance could be detected in the removed organs (lung,
liver and spleen) and spleen enlargement was also observed,
indicating that the orthotopic 4T1-Luc cells had metastasized to the
organs. Paclitaxel treatment reduced the luminance in the lung, and
less luminance was seen in the removed lung and none in the liver
(
Fig. 5
A). In the denbinobin (10 mg/kg)-treated group, no detectable
luminance was seen in the lung in the living animals (
Fig. 5
A).
Fig. 2. In vitro inhibition of Src kinase by denbinobin. 1064T1-Luc cells (A) orMDA-MB-231 cells (B) were incubated with 0–10 μM denbinobin for 30 min in the presence of EGF (100 ng/ml) for another 1 h, then equal amounts of total cell lysates were immunoprecipitated with 1μg of anti-Src antibody, followed by kinase assay using a tyrosine kinase kit. The absorbance was measured at 492 nm. The data represent the mean±S.E.M. for four replicates.⁎Pb.05 and ⁎⁎Pb.01 compared to the control group, respectively. The experiment was performed four times with similar results. 4 P.-H. Chen et al. / Journal of Nutritional Biochemistry xx (2010) xxx–xxx
Furthermore, in order to investigate the signaling molecules involved
in the metastatic model, immunofluorescence analysis was
per-formed on paraffin-embedded tumor tissues. Immunofluorescence
images of the orthotopic tumors again revealed that denbinobin
treatment significantly inhibited phosphorylation of Src, FAK, and
paxillin (
Fig. 5
B). These results suggest that denbinobin inhibits
breast cancer metastasis by suppressing the activity of Src-activated
downstream molecules.
Together, these results demonstrate that denbinobin
significant-ly inhibits the kinase activity of Src and that its inhibitory effect on
Src-mediated signals causes suppression of breast cancer migration
and metastasis.
Fig. 3. Inhibitory effects of denbinobin on breast cancer cell migration and invasion. (A) 2×105
4T1-Luc cells or 1×105
MDA-MB231 cells were incubated with 1, 3μM denbinobin for 30 min in the presence of EGF (100 ng/ml) for another 1 h, or transfected with 10μg of empty vector (EV) or CA-Src plasmid for 24 h, then cells harvested and placed in the top compartment of chemotaxis chamber with indicated treatment. Culture medium containing 10% FBS was added to the bottom compartment of the chamber. After 24 h, cell migration was measured as described in the Materials and Methods section. (B) Cells were seeded onto the upper chamber consisting of 8μm pore-size filters coated with Matrigel, then treated without or with denbinobin (1, 3μM) for 24 h in the absence or presence of EGF (100 ng/ml). Cells that invaded the filter were measured or counted in microscope. The data represent the mean±S.E.M. for three replicates.⁎Pb.05 and ⁎⁎Pb.01 comparing the indicated groups. The results shown are representative of those obtained in three independent experiments.
4. Discussion
Elevated Src expression and Src kinase activity have been seen in
many solid tumors, including breast cancer
[5,7]
. Src kinases are
transducers of signals and are activated by various cell-surface
receptors and interact with numerous substrates, mediating a wide
range of biological events, such as growth
[25]
, proliferation
[31]
,
angiogenesis
[32]
, invasion
[33]
, metastasis
[12,34]
, chemoresistance
[35]
and bone turnover
[36]
. The abolition of Src signals using Src
−/−mice or Src inhibitors significantly inhibits in vitro or in vivo tumor
progression, suggesting Src as a therapeutic target for cancer therapy
[4,5,27]
. Using several small-molecule Src inhibitors, inhibition of cell
migration and invasion is consistently seen
[17,34,37]
. Recent studies
have suggested that cancer cells may regulate their own adhesion to
matrix proteins by intracellular signals that alter the binding affinity
for matrix receptors
[38
–40]
.
In the present study, we demonstrated that denbinobin
markedly inhibited Src kinase and blocked the activity of Src-FAK
signaling pathways. FAK is phosphorylated by Src on a number of
tyrosine residues and, like Src, is associated with malignant
progression of breast cancer
[28,37,41]
. Our data showed that
decreased phosphorylation of FAK on Tyr925 was seen following
denbinobin treatment and that this correlated with a reduction in
motility. These results agree with previous finding indicating that
Fig. 4. Denbinobin inhibits the phosphorylation of Src and downstream proteins in breast cancer cells. 1×1054T1-Luc cells (A) or MDA-MB-231 cells (B) were cultured on coverslips for 1 h in the presence or absence of 3μM denbinobin (Den), then were fixed and stained overnight with the indicated antibodies at 4°C. The proteins are labeled with FITC (green, left panels), the nuclei are stained with DAPI (blue, center panels), and the merged images are shown on the right. The results shown are representative of those obtained in four independent experiments. Scale bar=50μm.
the Src kinase-dependent phosphorylation of Tyr925 in FAK is
important in controlling the extension and retraction of cell
protrusions or adhesion turnover
[42]
. Another substrate of Src,
p130Cas, is also involved in cell spreading, focal adhesion
formation, and invasion, and its phosphorylation was also decreased
following denbinobin treatment. A recent report suggested that FAK
promotes assembly of a Src-p130Cas-CRK-DOCK180 complex, which
activates Rac1 and subsequently promotes matrix
metalloprotein-ase-mediated matrix degradation
[28]
. Another study demonstrated
that Src kinase can directly phosphorylate paxillin or promote
paxillin phosphorylation indirectly by activation of FAK/cell
adhe-sion kinase-
β
[43,44]
. Paxillin functions as an adaptor protein that
facilitates the assembly of multi-protein complexes to regulate the
active state of Rho family proteins, e.g., Rac, and, thus, regulate the
dynamics of the actin cytoskeleton
[44]
. Our Western blot (
Fig. 1
)
and immunofluorescence (
Fig. 4
) results showed that denbinobin
treatment decreased cell migration at concentrations resulting in
detectable inhibition of phosphorylation of Src, FAK, p130Cas, and
paxillin, suggesting that denbinobin causes its migration inhibitory
effect by suppressing Src-FAK signals.
Fig. 5. Denbinobin suppresses breast cancer metastasis in vivo. (A) 4T1-Luc cells (2×105in 0.1 ml of medium) were injected into the left lower mammary fat pad of five-week-old
female BALB/c mice (five animals per group). After the orthotopic tumor volume reached 50 mm3(14 days after tumor cell injection), the mice were left untreated or were injected
intraperitoneally with paclitaxel (20 mg/kg) every 4 days or denbinobin (10 mg/kg) 3 times a week. An IVIS 200 biophotonic imager (Xenogen) was used to monitor metastasis of the breast carcinoma. At 40 days after implantation, the whole animals were imaged as described in the Materials and Methods section. The experiment was performed three times with similar results. (B) After 40 days implantation, the orthotopic tumors were removed, embedded in paraffin, and cut into 5-μm sections, which were treated with the indicated antibodies for 24 h at 4°C. The specific proteins are labeled by FITC (green) and the nuclei are stained with DAPI (blue). All the stained slides were viewed and photographed under an SP5 confocal laser-scanning microscope. The results shown are representative of those obtained in four independent experiments. Scale bar=25μm.
The 4T1 metastatic model is a suitable model for evaluating the
metastatic process in vivo
[45]
. The xenograft models, in which
human tumor cells are introduced into immunocompromised mice,
have been used extensively for the study of tumor growth and
metastasis
[45,46]
. However, while some human xenograft models
can approximate primary tumor growth in mice, replication of tumor
metastasis is rather problematic. For instance, human tumor cells
generally metastasize poorly in mice, and even when metastasis does
occur, unexpected and uncharacteristic conditions are often observed
[45,46]
. On the other hand, the 4T1 model, a syngeneic mouse model,
has been shown to be more appropriate for the evaluation of in vivo
metastatic process
[45]
. Such models are particularly useful as that
the metastatic tissues, the tumor microenvironment and the host are
from the same species and can therefore faithfully represent the
typical metastatic characteristics similar to those observed in cancer
patients
[45,47]
. This is important especially when considering the
significance of the microenvironment and tumor-host interactions to
tumor cell development. The 4T1 model also poses an important
advantage of allowing analyses to be carried out on animals with
normal immune function
[45]
. Since the immune system plays an
important role in the development and progression of cancer, models
that can be applied on immunocompetent mice are essential for
analysis of cancer progression and evaluation of therapeutics for
cancer treatment
[45]
.
Denbinobin, known as Shi-Hu in Chinese medicine, has been used
to treat the common cold and fever for centuries in the practice of
traditional medicine in China. It has been reported that denbinobin
exhibits anti-oxidant effect and cytotoxicity against human lung
carcinoma
[19,22]
, colorectal cancer
[23]
, and myelogenous leukemia
cell lines
[21]
. In the present study, we observed that denbinobin
suppressed breast cancer metastasis significantly. This novel
obser-vation allows us to hypothesize that more potent drugs for
suppressing breast cancer metastasis could be developed from
compounds sharing similar structural features with denbinobin
with proper chemical modifications.
In summary, our study demonstrates that denbinobin significantly
inhibits Src kinase activity and the Src-mediated phosphorylation of
downstream molecules in two types of breast cancer cells. This
inhibitory effect results in a decrease in breast cancer cell migration in
vitro and metastasis in vivo. These results indicate that denbinobin
has potential application in the treatment of breast cancer metastasis.
References
[1] Yoshida BA, Sokoloff MM, Welch DR, Rinker-Schaeler CW. Metastasis-suppressor genes: a review and perspective. J Natl Cancer Inst 2000;92:1717–30. [2] Woodhouse EC, Chuaqui RF, Liotta LA. General mechanisms of metastasis. Cancer
1997;80:1529–37.
[3] Shih JY, Yang SC, Hong TM, Yuan A, Chen JJ, Yu CJ, et al. Collapsin response mediator protein-1 and the invasion and metastasis of cancer cells. J Natl Cancer Inst 2001;93:1392–400.
[4] Hiscox S, Nicholson RI. Src inhibitors in breast cancer therapy. Expert Opin Ther Targets 2008;12:757–67.
[5] Finn RS. Targeting Src in breast cancer. Ann Oncol 2008;19:1379–86.
[6] Zhu S, Bjorge JD, Cheng HC, Fujita DJ, Decreased CHK. protein levels are associated with Src activation in colon cancer cells. Oncogene 2008;27:2027–34. [7] Myoui A, Nishimura R, Williams PJ, Hiraga T, Tamura D, Michigami T, et al. C-SRC
tyrosine kinase activity is associated with tumor colonization in bone and lung in an animal model of human breast cancer metastasis. Cancer Res 2003;63: 5028–33.
[8] Ichihara N, Kubota Y, Kitanaka A, Tanaka T, Taminato T. Inhibition of Src reduces gemcitabine-induced cytotoxicity in human pancreatic cancer cell lines. Cancer Lett 2008;260:155–62.
[9] Ishizawar R, Parsons SJ. c-Src and cooperating partners in human cancer. Cancer Cell 2004;6:209–14.
[10] Frame MC. Newest findings on the oldest oncogene; how activated Src does it. J Cell Sci 2004;117:989–98.
[11] Yeatman TJ. A renaissance for Src. Nat Rev Cancer 2004;4:470–80.
[12] Weis S, Cui J, Barnes L, Cheresh D. Endothelial barrier disruption by VEGF-mediated Src activity potentiates tumor cell extravasation and metastasis. J Cell Biol 2004;167:223–9.
[13] Chou MT, Wang J, Fujita DJ. Src kinase becomes preferentially associated with the VEGFR, KDR/Flk-1, following VEGF stimulation of vascular endothelial cells. BMC Biochem 2002;3:32–42.
[14] Palmer A, Zimmer M, Erdmann KS, Eulenburg V, Porthin A, Heumann R, et al. EphrinB phosphorylation and reverse signaling: regulation by Src kinases and PTP-BL phosphatase. Mol Cell 2002;9:725–37.
[15] Bromann PA, Korkaya H, Courtneidge SA. The interplay between Src family kinases and receptor tyrosine kinases. Oncogene 2004;23:7957–68.
[16] Playford MP, Schaller MD. The interplay between Src and integrins in normal and tumor biology. Oncogene 2004;23:7928–46.
[17] Gonzalez L, Agullo-Ortuno MT, Garcia-Martinez JM, Calcabrini A, Gamallo C, Palacios J, et al. Role of c-Src in human MCF7 breast cancer cell tumorigenesis. J Biol Chem 2006;281:20851–64.
[18] Lee YH, Park JD, Baek NI, Kim SI, Ahn BZ. In vitro and in vivo antitumoral phenanthrenes from the aerial parts of Dendrobium nobile. Planta Med 1995;61: 178–80.
[19] Chen HY, Shiao MS, Huang YL, Shen CC, Lin YL, Kuo YH, et al. Antioxidant principles from Ephemerantha lonchophylla. J Nat Prod 1999;62:1225–7. [20] Lin TH, Chang SJ, Chen CC, Wang JP, Tsao LT. Two phenanthraquinones from
Dendrobium moniliforme. J Nat Prod 2001;64:1084–6.
[21] Huang YC, Guh JH, Teng CM. Denbinobin-mediated anticancer effect in human K562 leukemia cells: role in tubulin polymerization and Bcr-Abl activity. J Biomed Sci 2005;12:113–21.
[22] Kuo CT, Chen BC, Yu CC, Weng CM, Hsu MJ, Chen CC, et al. Apoptosis signal-regulating kinase 1 mediates denbinobin-induced apoptosis in human lung adenocarcinoma cells. J Biomed Sci 2009;16:43.
[23] Chen TH, Pan SL, Guh JH, Chen CC, Huang YT, Pai HC, et al. Denbinobin induces apoptosis by apoptosis-inducing factor releasing and DNA damage in human colorectal cancer HCT-116 cells. Naunyn Schmiedebergs Arch Pharmacol 2008;378:447–57.
[24] Thomas SM, Brugge JS. Cellular functions regulated by Src family kinases. Annu Rev Cell Dev Biol 1997;13:513–609.
[25] Park EJ, Min HY, Chung HJ, Hong JY, Kang YJ, Hung TM, et al. Down-regulation of c-Src/EGFR-mediated signaling activation is involved in the honokiol-induced cell cycle arrest and apoptosis in MDA-MB-231 human breast cancer cells. Cancer Lett 2009;277:133–40.
[26] Zhou Q, Shaw PG, Davidson NE. Inhibition of histone deacetylase suppresses EGF signaling pathways by destabilizing EGFR mRNA in ER-negative human breast cancer cells. Breast Cancer Res Treat 2009;117:443–51.
[27] Trevino JG, Summy JM, Gallick GE. Src inhibitors as potential therapeutic agents for human cancers. Mini Rev Med Chem 2006;6:681–7.
[28] McLean GW, Carragher NO, Avizienyte E, Evans J, Brunton VG, Frame MC. The role of focal-adhesion kinase in cancer— a new therapeutic opportunity. Nat Rev Cancer 2005;5:505–15.
[29] Sharma A, Mayer BJ. Phosphorylation of p130Cas initiates Rac activation and membrane ruffling. BMC Cell Biol 2008;9:50.
[30] Brown MC, Turner CE. Paxillin: adapting to change. Physiol Rev 2004;84:1315–39. [31] Fox EM, Bernaciak TM, Wen J, Weaver AM, Shupnik MA, Silva CM. Signal transducer and activator of transcription 5b, c-Src, and epidermal growth factor receptor signaling play integral roles in estrogen-stimulated proliferation of estrogen receptor-positive breast cancer cells. Mol Endocrinol 2008;22: 1781–96.
[32] Choi S, Lee SA, Kwak TK, Kim HJ, Lee MJ, Ye SK, et al. Cooperation between integrin alpha5 and tetraspan TM4SF5 regulates VEGF-mediated angiogenic activity. Blood 2009;113:1845–55.
[33] Leroy C, Fialin C, Sirvent A, Simon V, Urbach S, Poncet J, et al. Quantitative phosphoproteomics reveals a cluster of tyrosine kinases that mediates SRC invasive activity in advanced colon carcinoma cells. Cancer Res 2009;69: 2279–86.
[34] Jallal H, Valentino ML, Chen G, Boschelli F, Ali S, Rabbani SAA. Src/Abl kinase inhibitor, SKI-606, blocks breast cancer invasion, growth, and metastasis in vitro and in vivo. Cancer Res 2007;67:1580–8.
[35] Santen RJ, Fan P, Zhang Z, Bao Y, Song RX, Yue W. Estrogen signals via an extra-nuclear pathway involving IGF-1R and EGFR in tamoxifen-sensitive and -resistant breast cancer cells. Steroids 2009;74:586–94.
[36] Soysa NS, Alles N, Shimokawa H, Jimi E, Aoki K, Ohya K. Inhibition of the classical NF-kappaB pathway prevents osteoclast bone-resorbing activity. J Bone Miner Metab 2009;27:131–9.
[37] Vultur A, Buettner R, Kowolik C, Liang W, Smith D, Boschelli F, et al. SKI-606 (bosutinib), a novel Src kinase inhibitor, suppresses migration and invasion of human breast cancer cells. Mol Cancer Ther 2008;7:1185–94.
[38] von Sengbusch A, Gassmann P, Fisch KM, Enns A, Nicolson GL, Haier J. Focal adhesion kinase regulates metastatic adhesion of carcinoma cells within liver sinusoids. Am J Pathol 2005;166:585–96.
[39] Van der Voort van Zyp J, Thamilselvan V, Walsh M, Polin L, Basson MD. Extracellular pressure stimulates colon cancer cell adhesion in vitro and to surgical wounds by Src (sarcoma protein) activation. Am J Surg 2004;188: 467–73.
[40] Basson MD. An intracellular signal pathway that regulates cancer cell adhesion in response to extracellular forces. Cancer Res 2008;68:2–4.
[41] Watermann DO, Gabriel B, Jager M, Orlowska-Volk M, Hasenburg A, zur Hausen A, et al. Specific induction of pp125 focal adhesion kinase in human breast cancer. Br J Cancer 2005;93:694–8.
[42] Brunton VG, Avizienyte E, Fincham VJ, Serrels B, Metcalf 3rd CA, Sawyer TK, et al. Identification of Src-specific phosphorylation site on focal adhesion kinase: 8 P.-H. Chen et al. / Journal of Nutritional Biochemistry xx (2010) xxx–xxx
dissection of the role of Src SH2 and catalytic functions and their consequences for tumor cell behavior. Cancer Res 2005;65:1335–42.
[43] Sachdev S, Bu Y, Gelman IH. Paxillin-Y118 phosphorylation contributes to the control of Src-induced anchorage-independent growth by FAK and adhesion. BMC Cancer 2009;9:12.
[44] Schaller MD. Paxillin: a focal adhesion-associated adaptor protein. Oncogene 2001;20:6459–72.
[45] Tao K, Fang M, Alroy J, Sahagian GG. Imagable 4T1 model for the study of late stage breast cancer. BMC Cancer 2008;8:228.
[46] Bibby MC. Orthotopic models of cancer for preclinical drug evaluation: advantages and disadvantages. Eur J Cancer 2004;40:852–7.
[47] Vernon AE, Bakewell SJ, Chodosh LA. Deciphering the molecular basis of breast cancer metastasis with mouse models. Rev Endocr Metab Disord 2007;8: 199–213.