The Crawling Speeds of the Three Sympatric Laticaudine Sea Kraits
on Different Substrates
S. Wang and Ming-Chung Tu*
Department of Life Science, National Taiwan Normal University Taipei, Taiwan
(Received: 29 April 2013, accepted: 7 May 2013)
ABSTRACT
Three sympatric laticaudine sea kraits are abundant at Orchid Island during the night time. Among habitats of three kinds of substrates (shingle, coral, fine gravel), they are especially abundant at the coral reef areas. An investigation on the crawling speeds of the sea kraits on the three substrates was conducted in our study. On the basis of crawling speed may be affected by the substrates, we predicted that sea kraits may have different crawling speeds on the various substrates. Based on the assumptions that a better locomotor performance at the most frequently utilized habitat should be favored by animals, a better crawling ability of sea kraits should be found on the coral surface. Although our results showed a different sprint speeds on the three substrates, the excellent crawling ability on the fine gravel surface was beyond our prediction.
Key words: sea krait, Laticauda, crawling speed, substrate, habitat
Introduction
Better locomotor performance can benefit animals in various ways. Garter snakes (Thamnophis sirtalis fitchi) which have greater values of both speed and distance capability (the maximum distance a snake can crawl during the trial) are more likely to survive (Jayne and Bennett, 1990). High correlation between locomotor performance and survival was also found on lizards (Husak, 2006). In the collared lizard (Crotaphytus
collaris), a strong correlation between male sprint
speed and territory size indicates that faster males may have higher possibility to mate (Peterson and Husak, 2006). Besides, a higher sprint speed also helps males in reproduction (Husak et al., 2006) and survival (Peterson and Husak, 2006).
Predation risk is an important selective force able to influence morphology, physiology, behavior, and life-history traits in the natural populations (Steiner, 2007; Hossie et al., 2010), and resource acquisition may affect animals’ energy allocation between growth and reproduction (Bonnet et al., 1998; Kubicka, 2009). Resource acquisition and predator avoidance are not only important to animals’ survival but also the crucial factors in the
process of evolution.
There are three laticaudine sea kraits distributed in the waters around Taiwan, and they are especially abundant in certain areas of our study site at Orchid Island. Three kinds of microhabitats characterized by various substrates were found there, however, the sea kraits were often found sympatrically at the coral reef areas during the night time (Liu et al., 2012). Although the sea kraits spend lots of time foraging in the ocean, it is also important to crawl well for these amphibious locomotors due to their needs to return to the land for mating, oviposition, digestion and skin sloughing (Heatwole 1999).
Locomotor performance is supposed to have strong influence on animal’s survival for its likely role in affecting predator avoidance or acquiring prey, and locomotor systems generally function well in the circumstances where they are usually employed (Aubret and Shine, 2008; Losos, 1990). For terrestrial lateral undulatory locomotors like snakes, reactive forces are formed when their bodies push against irregularities of the substrate. Once the forces are great enough to overcome the sliding frictional resistance, the animals are moving (Gans, 1974). Friction is responsible for the
generation of propulsive force in the terrestrial locomotion. In addition, previous work in snake crawling has revealed that the substrate may affect snakes’ locomotor mode (Byrnes and Jayne, 2010) and locomotor performance (Kelly et al., 2003). Therefore we expect that a differential locomotor performance on different substrates may be found in the sea kraits. A better crawling performance on the coral surface should be found in our study for their frequently retreating to the coral reef during the night time.
Materials and Methods Study site and study species
Orchid Island (Lanyu, 22°03’ N, 121°32’ E) is a 45 km2 island situated 60 km off the south east of Taiwan where the amphibious laticaudine sea kraits are abundant. Laticauda colubriana, L. laticaudata and L. semifasciata distribute sympatrically in coral reef areas there. Our study focuses on the crawling speed differences of the three substrates in the 3 sea kraits.
Animal collection and housing
Sea krait surveys were conducted during 2008 and 2009. Females were neglected in our study for avoiding the potential effect of their gravidity. Males were collected by hand once sighted moving or retreating near the sea shore areas. All the snakes were maintained in damp cloth bags and were transported to National Taiwan Normal University, Taipei city. Measurements of snout–vent length (SVL), tail length, and body mass were taken from the snakes we caught. Each snake was housed individually in a plastic container (length × width × height = 27 × 24 × 16 cm) filled with 3-cm-deep water. All the snakes were fastened and placed in a room which was kept at a temperature of 25°C where a 12h of light photoperiod was provided during the testing period. After the completion of the testing, all the sea kraits were released at the site where they had been collected.
Crawling speeds
A straight acrylic racetrack (3.0 m long, 0.5 m wide and 0.5 m deep) was used in all the locomotor performance tests. The bottom of the racetrack was painted with black lines in every 25 cm. 10 sets of cameras (30 fps, VG4C-XP-V, Chateau Technical Corp.) with digital timer were suspended by the
racetrack in every 25 cm. The first camera set was placed at the 50 cm of the racetrack. The time that the snakes crossed the lines could be recorded since the camera sets are just hung above the black lines. Three substrates were glued on the board (3.0 m long, 0.5 m wide) as crawling surfaces for terrestrial locomotor performance tests. 3 substrates (shingle, coral and fine gravel) were provided in the crawling test. All the snakes were tested in random orders. Each snake was released at one end of the racetrack and encouraged to move down it by light taps on the tail. We stopped the test when we found a snake had stopped moving or began moving in the opposite direction. Then we repeated the test after at least one hour. For each snake, sprint speed was the fastest speed calculated in any section of the racetrack, whereas average speed was the mean speed measured after the completion of the trial. Sprint speed and average speed are adopted as the locomotor performance indices for the following analyses. Data on the average crawling speeds tested on the various substrates are logarithmic transformed for the variance heterogeneity. The residuals from a regression of the log-transformed data against SVL were analyzed in order to avoid any potential influence of body size on the performance. A subsequent two-factor ANOVA with repeated-measures on substrates (coral/ shingle/gravel) was conducted for the data analysis. Data of sprint speeds were also transformed (residuals of speeds regressed against SVL) in order to avoid any potential influence of body length. Results
50 individuals (10 L. colubrina, 22 L.
laticaudata and 18 L. semifasciata) were tested in
the trials measuring their crawling speeds on the three different substrates. Both sprint speed and average speed were adopted as performance indices in our study.
The species effect and the substrate effect are both statistically insignificant on the average crawling speed (Fig. 1).
A significant substrate effect (F2, 94=23.87; P <0.0001) and an insignificant species effect (F2, 94=0.99; P =0.38) on the sprint speed are found in the data analysis, however, the highly significant interaction term (F4, 94=5.95; P =0.0003) reflects that these sea kraits may not have consistent sprint speeds on different substrates. A subsequent
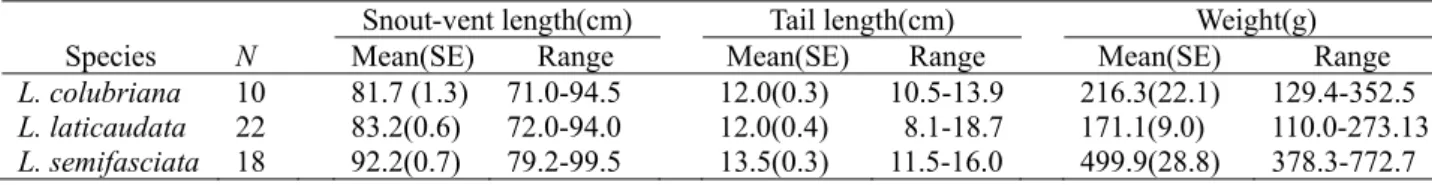
Table 1. Sample sizes and body sizes of sea kraits measured for the crawling ability test.
Snout-vent length(cm) Tail length(cm) Weight(g)
Species N Mean(SE) Range Mean(SE) Range Mean(SE) Range
L. colubriana 10 81.7 (1.3) 71.0-94.5 12.0(0.3) 10.5-13.9 216.3(22.1) 129.4-352.5
L. laticaudata 22 83.2(0.6) 72.0-94.0 12.0(0.4) 8.1-18.7 171.1(9.0) 110.0-273.13
L. semifasciata 18 92.2(0.7) 79.2-99.5 13.5(0.3) 11.5-16.0 499.9(28.8) 378.3-772.7
Tukey’s test shows that there is locomotor performance difference existing among groups (P < 0.05, Tukey’s test). Laticauda laticaudata shows no sprint speed difference on the 3 substrates (Fig. 2). There is no difference in sprint speed found on the coral and shingle surface for L. colubrina and L.
semifasciata, but a significantly faster sprint speed
is achieved on the gravel surface for them. Discussion
Ecomorphology presumes a match between an organism’s environment and its locomotor abilities, and previous studies also reveal that animals function most effectively at the microhabitats they inhabited (Losos et al., 1993; Vanhooydonck and Van Damme, 2003; Vanhooydonck and Van Damme, 2001).
Based on the assumption that a better locomotor performance at the most frequently utilized microhabitat should be favored, we suspected that an excellent crawling ability on coral surfaces should be found in the sea kraits for their being used to inhabit the areas of coral reef. The substrate effect on sea kraits’ crawling speeds was insignificant when average crawling speeds was adopted as the locomotor performance index, but the substrates had a significant effect on their sprint crawling speeds. Although L. laticaudata showed no sprint crawling performance difference on the three substrates, other sea kraits had significant higher sprint speeds on the fine gravel surface. The result of their better sprinting performance on the fine gravel was beyond our prediction.
Scribner and Weatherhead (1995) have demonstrated that the snake’s terrestrial locomotor speed is affected by the friction difference among the substrates. Among the three substrates, the rough coral surface may help sea kraits moving forward efficiently for enough reaction force generation. But research also reveals that the crawling speed varies with the substrate area that the snake’s body is in contact with (Finkler and Claussen, 1999). A higher crawling speed on the
L. colubrina L. laticaudata L. semifasciata
Average crawling spe ed (m/sec) 0.00 0.05 0.10 0.15 0.20 0.25 0.30 Coral Shingle Fine Gravel
Figure 1. Average crawling speeds of the 3 laticaudine
sea kraits tested on different substrates.
L. colubrina L. laticaudata L. semifasciata
Sprint speed (m/sec)
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 Coral Shingle Fine gravel A B B A A A b b a
Figure 2. Sprinting speeds of the three laticaudine sea
kraits tested on different substrates. Bars labeled with the same letter do not differ significantly (Tukey's test, alpha=0.05).
fine gravel can be achieved if the fine gravel surface can provide more push points and larger contacting area as well. However, our data on the substrate character is inadequate to explain sea kraits’ better performance on the fine gravel surface.
Locomotor performance is supposed to have a strong influence on an animal’s survival for its likely role in affecting predator avoidance or acquisition of prey, and Anolis lizards are reported to avoid habitats in which their performance
capacities are submaximal (Irschick and Losos, 1999). Although sea kraits were found to have a better crawling performance on fine gravel surfaces, the fine gravel beaches do not provide any shelter for their not being a fossorial species (Bonnet et al., 2009). Apart from predator avoidance, snake’s habitat exploitation can also be affected by the prey availability (Bonnet, 2012; Heard et al., 2004; Madsen and Shine, 1996).
Moreover, the sea snakes’ habitat utilization can also be influenced by the vulnerability to predation (Kerford et al., 2008), available fresh water sources (Lillywhite et al., 2008), precipitation (Lillywhite and Tu, 2011), salinity (Brischoux et al., 2012) and environmental temperature (Bonnet et al., 2009; Lelièvre et al., 2011). Snakes may have other ecological needs which must be met for them in order to inhabit a suitable microhabitat. These factors may explain our findings of the sea kraits suboptimal performance in their preferred coral reef microhabitat.
Acknowledgments
We are greatly indebted to K. Lai for his help on designing the racetrack. We express our appreciation to and also to C.-J. Chang, C. Huang for their assistance on field work. This research was funded by the National Science Council of Taiwan (NSC97-2621-B-003-004-MY3).
References
Aubret F and Shine R. 2008. The origin of evolutionary innovations: locomotor consequences of tail shape in aquatic snakes. Funct. Ecol. 22: 317-322.
Bonnet X. 2012. Long-term field study of sea kraits in New Caledonia: fundamental issues and conservation. Integ. Comp. Biol. 52: 281-295. Bonnet X, Brischoux F, Pearson D and Rivalan P.
2009. Beach-rock as a keystone habitat for sea kraits. Environ. Conserv. 36: 62-70.
Bonnet X, Bradshaw D and Shine R. 1998. Capital versus income breeding: an ectothermic perspective. Oikos 83: 333-342.
Brischoux F, Tingley R, Shine R and Lillywhite HB. 2012. Salinity influences the distribution of marine snakes: implications for evolutionary transitions to marine life. Ecography 35: 994-1003.
Byrnes G and Jayne BC. 2010. Substrate diameter and compliance affect the gripping strategies and locomotor mode of climbing boa constrictors. J. Exp. Biol. 213:4249-4256. Finkler MS and Claussen DL. 1999. Influence of
temperature, body size, and inter-individual variation on forced and voluntary swimming and crawling speeds in Nerodia sipedon and
Regina septemvittata. J. Herpetol. 33:62-71.
Gans C. 1974. Biomechanics an approach to vertebrate biology. University of Michigan Press, Ann Arbor, Michigan.
Heard GW, Black D and Robertson P. 2004. Habitat use by the inland carpet python (Morelia spilota metcalfei: Pythonidae): seasonal relationships with habitat structure and prey distribution in a rural landscape. Aust. Ecol. 29: 446-460.
Heatwole H. 1999. Sea snakes. University of New South Wales Press, Sydney.
Higham TE. 2007. The integration of locomotion and prey capture in vertebrates: Morphology, behavior, and performance. Integ. Comp. Biol. 47: 82-95.
Hossie TJ, Ferland-Raymond B, Burness G and Murray DL. 2010. Morphological and Behavioural Responses of Frog Tadpoles to Perceived Predation Risk: A Possible Role for Corticosterone Mediation? Ecoscience 17: 100-108.
Husak JF. 2006. Does speed help you survive? A test with Collared Lizards of different ages. Funct. Ecol. 2006: 174-179.
Husak JF, Fox SF, Lovern MB and Bussche RA. 2006. Faster lizards sire more offspring: sexual selection on whole-animaimal performance. Evolution 60: 2122-2130.
Irschick DJ and Losos JB. 1999. Do lizards avoid habitats in which performance is submaximal? The relationship between sprinting capabilities and structural habitat use in Caribbean anoles. Am. Nat. 154: 293–305.
Jayne BC and Bennett AF. 1990. Selection on Locomotor Performance Capacity in a Natural Population of Garter Snakes. Evolution 44: 1204-1229.
Kelley KC, Arnold SJ and Gladstone J. 1997. The effects of substrate and vertebral number on locomotion in the garter snake Thamnophis
Kerford M, Wirsing AJ, Heithaus MR and Dill LM. 2008. Danger on the rise: habitat use by bar-bellied sea snakes in Shark Bay, Western Australia. Mar. Ecol. Prog. Series 358: 289-294.
Kubicka L. 2009. First grow, then breed and finally get fat: hierarchical allocation to life-history traits in a lizard with invariant clutch size. Funct. Ecol. 23: 595-601.
Lelièvre H, Blouin-Demers G, Pinaud D, Lisse H, Bonnet X and Lourdais O. 2011. Contrasted thermal preferences translate into divergences in habitat use and realized performance in two sympatric snakes. J. Zool., Lond. 284: 265-275.
Lillywhite HB and Tu M-C. 2011. Abundance of Sea Kraits Correlates with Precipitation. PLoS ONE 6(12): e28556.
Lillywhite HB, Babonis LS, Sheehy CM and Tu M-C. 2008. Sea snakes (Laticauda spp.) require fresh drinking water: implications for the distribution and persistence of populations. Physiol. Biochem. Zool. 81: 785-796.
Liu Y-L, Chen Y-H, Lillywhite HB and Tu M-C. 2012. Habitat selection by sea kraits (Laticauda spp.) at coastal sites of Orchid Island, Taiwan. Integ. Comp. Biol. 52: 274-280.
Losos JB, Walton BM and Bennett AF. 1993.
Trade-offs between sprinting and clinging ability in Kenyan chameleons. Funct. Ecol. 7: 281–286.
Losos BJ. 1990. The Evolution of Form and Function: Morphology and Locomotor Performance in West Indian Anolis Lizards. Evolution 44: 1189-1203.
Madsen T and Shine R. 1996. Seasonal migration of predators and prey-a study of pythons and rats in tropical Aus. Ecol. 77: 149-156.
Peterson CC and Husak JF. 2006. Locomotor Performance and Sexual Selection: Individual Variation in Sprint Speed of Collared Lizards (Crotaphytus collaris) Copeia, 2006: 216-224. Scribner, S. J. and P. J. Weatherhead. 1995.
Locomotion and antipredator behaviour in three species of semi-aquatic snakes. Can. J. Zool. 73: 321-329.
Steiner UK. 2007. Investment in defense and cost of predator-induced defense along a resource gradient. Oecologia, 152: 201-210.
Vanhooydonck B and Van Damme R. 2003. Relationships between locomotor performance, microhabitat use and antipredator behaviour in lacertid lizards. Funct. Ecol. 17:160–169. Vanhooydonck B and Van Damme R. 2001.
Evolutionary tradeoffs in locomotor capacities in lacertid lizards: are splendid sprinters clumsy climbers? J. Evol. Biol. 14: 46–54.