Appl Microbiol Biotechnol (2006) 72: 1254–1262 DOI 10.1007/s00253-006-0404-8
A P P L I E D M I C R O B I A L A N D C E L L P H Y S I O L O G Y
Chun-Lin Lee . Jyh-Jye Wang . Shing-Lin Kuo . Tzu-Ming Pan
Monascus fermentation of dioscorea for increasing
the production of cholesterol-lowering agent
—monacolin
K and antiinflammation agent
—monascin
Received: 28 December 2005 / Revised: 26 February 2006 / Accepted: 5 March 2006 / Published online: 28 March 2006
# Springer-Verlag 2006
Abstract Monacolin K, an inhibitor for cholesterol syn-thesis, is the secondary metabolite of Monascus species. The formation of the secondary metabolites of the Monascus species is affected by cultivation environment and method. This research uses sweet potato (Ipomoeaba-tatas), potato (Solanum tuberosum), casava (Manihotescu-lenta), and dioscorea (Dioscoreabatatas) as the substrates and discusses the best substrate to produce monacolin K. The results show that Monascuspurpureus NTU 301, with dioscorea as the substrate, can produce monacolin K at 2,584 mg kg−1, which is 5.37 times to that resulted when rice is used as the substrate. In addition, more amount of yellow pigment can be found in Monascus-fermented dioscorea than in Monascus-fermented rice. The certain composition of yellow pigment is identified as monascin, which has been shown as an antiinflammation agent exhibiting potent inhibitory effects on 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-induced inflammation in mice in previous studies. Therefore, dioscorea is concluded to be the best substrate for Monascus species to produce the cholesterol-lowering agent—monacolin K and antiinflam-mation agent—monascin.
Introduction
The Monascus species is a Chinese traditional fermentation fungus used on food for over thousands of years in China, and its special effect and application on food has recorded in ancient Chinese record. Red mold rice is obtained
usually by cultivation of Monascus species on the rice grains. The traditional manufacture of red mold rice include the following steps: rice is washed, soaked in water for 24 h, then it is drained, steamed, sterilized, fermented, and dried (Dizon and Sanchez1984).
Monascus species have been proven to produce many functional secondary metabolites. These pigments (yellow pigment: ankaflavin and monascin; orange pigment: monascorubrin and rubropunctanin; red pigment: monas-corubramine and rubropuctamine) were investigated and applied to the food colorant in early study (Martinkova et al. 1999; Wong and Koehler 1981). In current study, Monascus-fermented product was gradually regarded as the functional food because the monacolin K (antihyper-cholesterolemic agents), γ-aminobutyric acid (GABA) (hypotensive agent), and dimerumic acid (antioxidant) were found (Su et al.2003; Aniya et al.1999).
Endo (1979) discovered that a more active methylated form of compactin known as monacolin K would be formed in the broths of Monascus ruber. Monacolin K, known as a statin compound, has been regarded as a cholesterol-lowering agent because it was proven to be a potent competitive inhibitor of 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMG-CoA reductase) (Albert et al. 1980; Serizawa et al. 1983). In the current study, Monascus-fermented rice has been demonstrated to per-form significant hypolipidemic effects in hyperlipidemia hamster model (Lee et al.2005). Therefore, it was proven to have special effect when used as a cholesterol-lowering drug. The relationship between cholesterol and atheroscle-rosis has long been of concerned, and an excessive concentration of cholesterol may lead to coronary artery disease or cause death (Carlson1982).
The previous study on the secondary metabolites of Monascus species, found that the biomass growth and the types and production of metabolites would be directly or indirectly affected by the cultivation environment and cultivation methods. As far as the cultivation methods were concerned, Lin (1973) states that solid state cultivation results in higher pigment yield than cultivation in shaken culture and conclude that this phenomenon is due to the C.-L. Lee . T.-M. Pan (*)
Institute of Microbiology and Biochemistry, National Taiwan University,
1, Sec. 4, Roosevelt Road, Tapei 10617, Taiwan e-mail: [email protected] Tel.: +886-2-33664519 Fax: +886-2-23627044 J.-J. Wang . S.-L. Kuo
Department of Biotechnology, Tajen University, Ping Tung, Taiwan
fact that pigments are released into grains under solid state culture, and the pigments are accumulated in the mycelium under submerged cultivation. Wang et al. (1999) have shown that under solid state cultivation, production conditions of lovastatin by Monascus ruber are different from those of other pigment formation. The report also points out the accumulated depth of substrate and the water content will affect lovastatin production, and the lovastatin production quantity resulted from solid state cultivation is 20 times more than that of liquid state cultivation. Furthermore, our previous research also showed that different fermentation methods would affect monacolin K and GABA production (Su et al.2003; Wang et al.2003). Carbon and nitrogen sources are nutritional sources required for microbial growth. Generally, glucose is considered as the best carbon source for pigment formation (Broder and Koehler 1980; Lin and Demain 1991), but various strains also lead to different results. Santerre et al. (1995) compare the difference of pigment formations when glucose and ethanol are used as carbon sources, and found out that ethanol has better pigment formation ability when it is used as carbon source. Moreover, others like starch, maltose, sucrose, and galactose (Lin 1973; Panitz et al. 1991; Yoshimura et al.1975) are also good carbon sources. Carel and Shepherd (1977) study the effect of Monascus species growth by different nitrogen sources and find out that adding yeast extract to broth medium is helpful for the biomass production, but it has adverse effect on pigment formation. In addition, ammonium chloride, sodium nitrate, peptone, and monosodium glutamate have different effects on metabolite formation (Blanc et al. 1995b; Lin and Demain1991; Su et al.2003).
As far as substrate is concerned, Lin and Lizuka (1982) compare the effect of different substrates on pigment production and find that steamed bread (manton) was a good substrate. Besides, rice, bread, oat (Rashbaum and Barrington 1983), corn, or wheat grain (Hesseltine1965; Lin and Lizuka1982), all can be used as the substrate for Monascus species to produce large amount of pigment. Using fermentation method to result in huge amount of monacolin K is always one of the key studies for our research team. Currently, there are no studies on the influence of substrates on the monacolin K production reported in the literature. Therefore, the objective of this work is to seek for the substrate that could result in higher quantity of monacolin K production.
Materials and methods
Chemicals
γ-Amino-n-butyric acid (GABA), monacolin K, citrinin, isooctane are purchased from Sigma Chemical (St. Louis, MO, USA). LC grade acetonitrile is purchased from Merck (Darmstadat, Germany). Tryptone, yeast extract, peptone, malt extract, PDA broth, and bacto-agar are purchased from Difco (Detroit, MI, USA). Reagent grade ethyl acetate is purchased from ALPS Chem. (Taiwan).
Microorganism and seed cultures
The microorganisms used in this study include M. purpureus BCRC 31499, 31504, 31530, 31540, 32966, NTU 601, NTU 301, Monascus sp. BCRC 32807, 32808, and 32809. These strains are either purchased from the Bioresources Collection and Research Center (BCRC, Taiwan) or isolated from red mold rice. The culture strains are maintained on potato dextrose agar (PDA) slanted at 10 °C and transferred monthly. In addition, M. purpureus NTU 601 is the parental strain for the generation of mutants M. purpureus NTU 301. The preparation of seed culture is described in our previous study (Su et al.2003).
Red mold substrate preparation
The traditional red mold substrates preparation method has been proposed in our previous studies (Su et al.2003). All substrates (long-grain rice, Ipomoeabatatas; sweet potato, Solanumtuberosum; potato, Manihotesculenta; cassava, Manihotesculenta; and dioscorea, Dioscoreabatatas) are purchased from local supermarket and are used as the substrates for red mold metabolite production under solid state cultivation. Five hundred grams rice and the other substrates are, respectively, soaked in distilled water for 8 and 1 h. After that, excess water is removed with a sieve. The substrate is autoclaved for 20 min at 121 °C in a “koji-dish” (the koji-dish is made of wood with the dimension of 30×20×5 cm). After being cooled, the substrate is inoculated with a 5% (v/w) spore suspension (107 spores ml−1) and 0.3% (v/w) ethanol. The inoculated substrate is cultivated at 30 °C for 10 days. At the end of cultivation, the crushed and dried product with the mold was used for the experiments (Su et al. 2003). Besides, the optimum condition for the formation of red mold product is also investigated by using the response surface methodology (RSM) reported in our previous study (Wang et al.2003). The RSM applied in this research is stated as below: 500 g substrate is inoculated with a 5% (v/w) spore suspension (107spores ml−1) and 0.3% (v/w) ethanol. The inoculated substrate is cultivated at 30 °C for 10 days. In addition, during the culturing stage, 145 ml of water is added once every 12 h at a total of three times and the addition of water starts on the fifth day of cultivation.
Determination of the monacolin K concentration
Red mold product (1 g) is extracted with 5 ml ethyl acetate at 70 °C for 1.5 h. The suspension is then filtered through filter paper. The filtrate is evaporated to dryness under vacuum. After lactonization, 1 ml acetonitrile is added to the resulting mixture, followed by filtrating with 0.45μm pore size filter and analyzed by high performance liquid chromatography (HPLC, Model L-6200, Hitachi, Japan) (Wang et al. 2003). Chromatographic separation is conducted on a Beckman Ultrasphere ODS column (150×4.6 mm I.D.). Acetonitrile-phosphoric acid (65:35,
v/v) solution of 0.5% is used as the mobile phase. The eluent is pumped at a flow rate of 0.7 ml min−1. UV detection was set at 238 nm.
Preparation of mold extract and detection of citrinin
Red mold product (1 g) is extracted with 10 ml methanol at 50 °C for 1.5 h. The filtered extract is twice defatted with isooctane (Sigma). After adding an equal volume of water and acidifying to pH 4.5 with H2SO4 (50:50, v/v), the extract is partitioned with CHCl3. The lower phase is evaporated to dryness and is then dissolved in methanol, followed by filtering with 0.45 μm pore size filter and analyzed by HPLC (Blanc et al. 1995a). Citrinin is determined by HPLC on a C18 column (LiChroCART 250-4, Merck, Darmstadat, Germany) using the mobile phase with the composition of water to acetonitrile to triflouacetate (450:550:0.5). The flow rate is set at 1.0 ml min−1and the detector used is fluorescence detector (FL-1, Rainin, Wobum, MA, USA). The excitation and emission wavelength was set at 330 and 500 nm, respectively (Kycko et al.1998).
Pigment estimation
Pigment concentrations are estimated using spectropho-tometer set at 400 nm for yellow pigment and 500 nm for red pigment. The results are expressed as optical density units per gram of dried medium multiplied by dilution factor (Lin and Lizuka1982).
Identification of monacolin K by NMR
Purified monacolin K of red mold dioscorea was separated and collected by HPLC. Nuclear magnetic resonance spectroscopy (NMR) experiments were performed on a Bruker AMX 500 WB spectrometer (Bruker BioSpin, Rheinstetten, Germany) equipped with a z gradient probe head capable of producing gradients of strength 50 G/cm, T=300 K. DOSY-NMR experiments were run using the BPPLED pulse sequence (Gibbs and Johnson1991; Pelta et al.1998): total gradient duration, 1.4 ms; diffusion delay (Δ), 100 ms; eddy current delay (Te), 50 ms; gradients intensities varied geometrically from 0.6 to 50 G/cm; 16 1D spectra acquired (4,096 data points, 128 scans/exper-iment); after Fourier transformation and baseline correc-tion, the diffusion dimension was processed using the Bruker Xwinnmr package (version 2.6) which permits calculation of dependable and comparable integrals (Silvia et al.2002).
Identification of monacolin K and yellow pigment by LC/MS
Liquid chromatography and mass spectrometry (LC/MS) are carried out using a system equipped with LC system (Finnigan MAT Spectra system, San Jose, CA, USA) including quaternary pump, on-line degasser, column heater compart, autosampler, and a LCQ mass spectrometer (Finnigan MAT LCQ, San Jose, CA, USA) that consists of an ESI (electrospray ionization) interface and an ion trap mass analyzer. The software for the control of the equipment, and the acquired and treatment of data is Xcalibur Core Data System Software Revision 1.0 (Xcalibur Software, Herndon, VA, USA).
The separation by LC-MS is carried out by a reversed-phase C18 HPLC column (25 cm×4.6 mm I.D., 5 μm, Discovery, Bellefontia, PA, USA) with an isocratic elution of acetonitrile-water (55:45, v/v) at a flow rate of 0.5 ml min−1. All the analyses were performed using ESI interface with the following settings: positive ionization mode; temperature of the capillary, 25 °C; spray voltage, 4.5 kV; capillary voltage, 6 V; sheath gas (N2) flow, 30 AU; and auxiliary gas (N2) flow, 10 AU. The electrospray interface and mass spectrometric parameters were optimized to obtain maximum sensitivity unit resolution (Lee et al. 2006).
Results
M. purpureus NTU 301 induced from M. purpureus NTU 601 by using N-methyl-N′-nitro-N-nitrosoguanidine (NTG) method can produce monacolin K (481 mg kg−1) and GABA (5,210 mg kg−1) (Wang et al.2004). Sweet potato (Ipomoeabatatas), potato (Solanumtuberosum), casava (Manihotesculenta), and dioscorea (Dioscoreabatatas) are tubers that are rich in carbohydrates. We use these materials as the fermentation substrates of M. purpureus NTU 301. The content of monacolin K and citrinin obtained by using different substrates in RSM methods (Wang et al.2003) is reported in Table1. From the table, we find that dioscorea is the best substrate for producing monacolin K, and the amount of the monacolin K produced from M. purpureus NTU 301 is 2,584±127 mg kg−1which is 13.18 times more than that when sweet potato is used as the substrate, and is 5.37, 5.22, and 4.95 times more when rice, potato, and cassava is used as substrate, respectively. Even though the amount of monacolin K can be dramatically increased by using dioscorea as substrate, the other metabolites of Monascus species—citrinin, can also be increased from 0.23±0.01 to 0.35±0.01 mg kg−1. The increased ratio was about 1.52 times. Because monacolin K and citrinin are the metabolites of Monascus species, their productions are interrelated. Furthermore, we can see that from Table1, sweet potato used as the substrate would lead to decrease in monacolin K production of M. purpureus NTU 301 and M. purpureus NTU 601 as compared to rice. Dioscorea and sweet potato perform the opposite effect on the stimulation of monacolin K
production; therefore, this result clearly indicates that various substrates would crucially result in different monacolin K production even though they were rich in starch.
To understand whether M. purpureus NTU 301 used dioscorea as substrate could successfully stimulate the formation of monacolin K was one specific case or the common result obtained for all the other Monascus species; we choose other Monascus species to carry out fermenta-tion and try to find out the effect of using dioscorea as the substrate on monacolin K productions. As shown in Table 2, dioscorea used as the substrate of various Monascus strains can perform significant increase in monacolin K production, especially in Monascus sp. BCRC 32808, which can use dioscorea as substrate to form a remarkable increase at 8.36 times in monacolin K production as compared with using rice as substrate. Thus, we prove that dioscorea is a better substrate to produce monacolin K for all Monascus strains chosen by this study. During the fermentation with dioscorea or with rice, Monascus strains form different amount of monacolin K and pigment. Therefore, dioscorea and rice are used, respectively, as the substrate to find out the change of monacolin K and pigment production during the entire fermentation stage. The time course of monacolin K production is shown in Fig. 1. Monacolin K would be gradually formed from the second day and stagnated on the sixth day if rice is used as the substrate. However, monacolin K will be continually accumulated until the tenth day if dioscorea is used in place of rice. In addition,
the rapid formation phase of monacolin K is from the fifth day to the tenth day. After whole fermentation by M. purpureus NTU 301, monacolin K in fermented dioscorea reaches 2,584 mg kg−1, which is at least five times more than that of fermented rice.
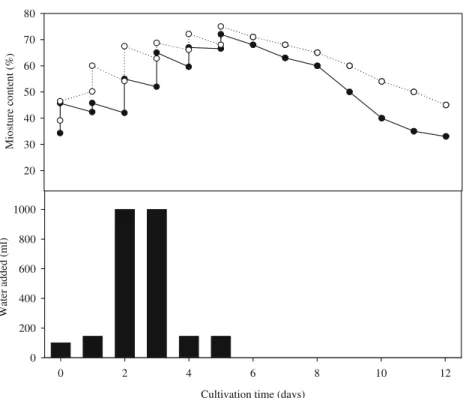
Moisture content is important for the growth of Monascus species, and a substrate which is excessively dry or wet is not suitable for the culture of Monascus species and production of secondary metabolite. Therefore, moisture content of fermented substrate should be con-trolled and monitored during fermentation stage. In this study, various amount of water is appropriately supple-mented to the substrate before fifth day and stop supplementing from sixth day to twelfth day to avoid making the substrate excessively wet. The changes of substrate moisture content during fermentation are shown in Fig. 2. Moisture content of fermented dioscorea is substantially higher than fermented rice before the third day and after the eighth day. The initial fermentation stage is involved with the growth of Monascus species, and the later fermentation stage is important to form secondary metabolite (Wang et al. 2004). Rice substrate cannot effectively protect moisture against evaporation especially during the later fermentation stage, which may be the reason why monacolin K cannot be continually produced after the eighth day. On the contrary, dioscorea leading to higher monacolin K production may result from the well moisture maintainability during later fermentation stage.
As far as pigment production is concerned, when dioscorea is used as the substrate of M. purpureus NTU Table 1 Production of monacolin K and citrinin on diverse substrates by Monascus strains
Substrate Strains Monacolin K (mg kg−1) Citrinin (mg kg−1) Red pigment (A500 g−1) Yellow pigment (A400 g−1)
Cassava M. purpureus NTU 601 265±13 0.31±0.01 45±5.3 32±1.4
Cassava M. purpureus NTU 301 522±34 0.41±0.01 50±3.8 34±3.8
Dioscorea M. purpureus NTU 601 1876±58 0.63±0.02 27±3.1 68±3.4
Dioscorea M. purpureus NTU 301 2584±127 0.35±0.01 30±5.0 74±4.7
Sweet potato M. purpureus NTU 601 128±24 0.45±0.01 43±2.1 40±5.2
Sweet potato M. purpureus NTU 301 196±21 0.23±0.01 51±3.2 39±3.9
Potato M. purpureus NTU 601 113±17 0.35±0.01 45±8.3 36±5.0
Potato M. purpureus NTU 301 495±37 0.21±0.01 48±6.9 32±4.8
Rice M. purpureus NTU 601 530±32 0.46±0.02 44±5.5 30±4.3
Rice M. purpureus NTU 301 481±33 0.23±0.01 50±4.2 41±4.8
Table 2 Production of monacolin K on solid culture by Monascus strainsa
aThe inoculated substrate was cultivated at 30 °C for 10 days. None of any nutrient sources was added
b
Control unfermented substrate
Strains Rice Dioscorea Increased time
Monacolin K (mg kg−1) Monacolin K (mg kg−1) M. purpureus BCRC 31499 119±21 134±21 1.13 M. purpureus BCRC 31504 25±8.7 49±5.1 1.96 M. purpureus BCRC 31530 54±6.4 224±27 4.15 M. purpureus BCRC 31540 43±5.5 134±21 3.12 M. purpureus BCRC 32966 26±6.5 192±28 7.38 Monascus sp. BCRC 32807 63±7.7 87±9.5 1.38 Monascus sp. BCRC 32808 14±2.0 117±18 8.36 Monascus sp. BCRC 32809 26±5.9 131±16 5.04 Controlb 0 0 –
301, certain amount of pigment will be produced. As shown in Fig. 1, Monascus-fermented rice and dioscorea have the different colorant on the surface. During the initial fermentation phase, more amount of yellow pigment is displayed on the surface of Monascus-fermented product (Fig. 3), but red pigment gradually takes the place of yellow pigment at later fermentation phase. However, we can find that the final Monascus-fermented dioscorea have more amount of yellow pigment than Monascus-fermented rice. Using rice as substrate always leads to Monascus being formed more red pigment during whole fermentation phase. Similar result is observed on other substrate fermented by Monascus species. Figure 3 shows that dioscorea can stimulate Monascus to form more yellow pigments as compared with rice, but there is a contrary result if red pigment is concerned. Yellow pigment of Monascus species including monascin and ankaflavin has been shown as an anticancer agent inducing cell death on Hep G2 cells (Su et al.2005) and antiinflammation agent
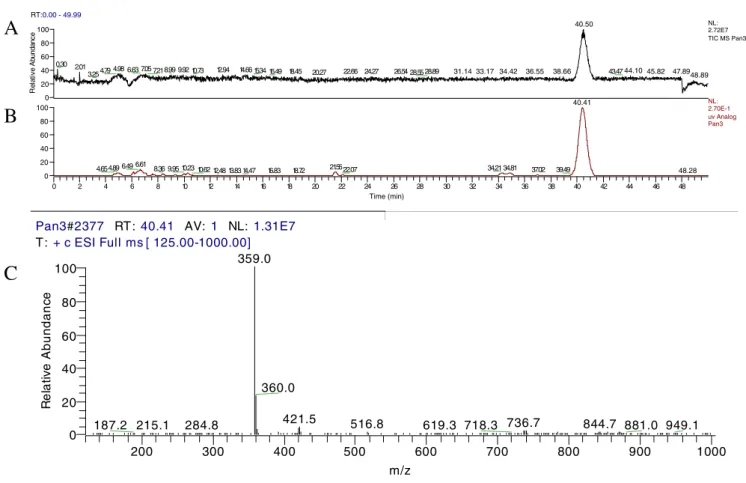
exhibiting potent inhibitory effects on 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-induced inflammation in mice (Akihisa et al. 2005). This study further uses liquid chromatography/mass spectra (LC/MS) to identify the certain composition of yellow pigment in Monascus-fermented dioscorea. As shown in Fig. 4, monascin (M+ 1=359.0), shown as mass spectrum, corresponded to the retention time at 36.80 min in LC chromatogram of 233 nm, which can demonstrate that monascin is the main yellow pigment of Monascus-fermented dioscorea.
Monacolin K has the elemental composition C24H36O5 (parent ion M+ 404.2563, calc. 404.2563). Mass spectral fragments at mass-to-change ratio m/e 386 (C24H34O4), 302 (C19H26O3), and 284 (C19H24O2) resulted from the loss of water, a C5H10O2 moiety, or both. Important fragment ions further appeared at m/e 269 (C18H21O2), 224 (C17H20), 197–199 (C15H18+1), 174 (C13H18), 169 (C13H13), 159 (C12H15), and 157 (C12H13) (Albert et al.
1980). To confirm the increased compound of red mold dioscorea, monacolin K, HPLC, MS, and NMR are used to identify the structure of purified monacolin K from the extract of red mold dioscorea. Certainly, through HPLC, NMR, and MS analysis, it has proven that red mold dioscorea does contain monacolin K and the production is higher than red mold rice (data not shown).
Discussion
Monacolin K, shown as a cholesterol-lowering agent, leads to Monascus product being promoted to the functional food. For this reason, current studies focus the goal on the monacolin K production of Monascus by using different culture condition. However, there is no universal and excellent method for stimulating monacolin K formation. Formerly, Monascus was used to being fermented under steamed rice to obtain more red pigment in the East Asia. There are few Monascus studies about using other kinds of substrate to produce monacolin K besides steamed rice. As far as we know, the composition of solid substrate is very important for the production of secondary metabolite and the growth of Monascus. In this study, we can find that a different substrate leads to Monascus species being produced with different levels of monacolin K and citrinin (Table 1). This research successfully used dioscorea as a substrate and obtained more monacolin K after Monascus species fermentation. The increased level was 5.37 times more than that for rice production. To our knowledge, there was no research concerning monacolin K production studies by using dioscorea as the fermentation substrate in the previous literature.
Many Monascus studies omitted detecting the citrinin concentration of Monascus product as monacolin K production is increased by a culture condition (Wang et al.1999,1998). Citrinin, grouped into a kind of polyketide derivatives of Monascus, is formed via the similar biosyn-thesis pathway of monacolin K (Wang et al.2003,2004). Therefore, citrinin production is usually increased as monacolin K production is increased by the culture
0 2 4 6 8 10 12 14 Y ellow pig m ent ( A 400 /g) 0 20 40 60 80 R ed p igm ent (A50 0/g) 0 20 40 60 80 Mon acolin K concen tratio n ( m g/kg) 0 500 1000 1500 2000 2500 3000
Fermentation time (day)
Fig. 1 Time course of production on monacolin K, red pigment and yellow pigment by M. Purpureus NTU301 (• rice grain; ○ dioscorea)
condition (Wang et al. 2003). In this study, Monascus fermentation using dioscorea substrate can result in a significant increase in monacolin K production and less increase in citrinin production. An increase of monacolin K production can be found in many Monascus-fermented dioscorea when compared to Monascus-fermented rice. It is important to note that the increase of monacolin K production is higher than that of citrinin production as far as the metabolite of the same Monascus strain is concerned. Monacolin K would be formed between middle and later phase because it was a secondary metabolite of Monascus species. This study demonstrated that monacolin K formation of Monascus using rice as substrate is carried out from the 2nd day to the 6th day (Fig.1), and the potato, sweet potato, and cassava perform similar tendency as well (data not shown). Dioscorea is the optimal substrate to extend the time of monacolin K formation of Monascus due to the probable reason that the specific composition or
condition of dioscorea is suitable for the growth of Monascus at later fermentation phase. In contrast, the reason why rice or other substrates with rich carbohydrate cannot lead to an extended duration for monacolin K formation at the later fermentation stage is due to an unseemly condition provided by the substrate for mon-acolin K formation or the growth of Monascus. In addition, moisture content of substrate is demonstrated as an important factor for the solid culture of Monascus (Wang et al.2004). Substrate with too much or too less moisture content is unsuitable for the growth of Monascus. The moisture is easier to evaporate in steamed rice than in steamed dioscorea during the fermentation process and dioscorea consists of more amylose than rice, so it is probably difficult to evaporate the moisture of steamed dioscorea and convert dioscorea starch into an aging structure (Fig.2). That is a probable reason for extending
Fig. 3 Photographs of final fermented rice (a) and fermented dioscorea (b) by M. purpureus NTU301
Cultivation time (days)
0 2 4 6 8 10 12 Water added (ml) 0 200 400 600 800 1000 M io st u re co n ten t ( % ) 20 30 40 50 60 70 80 Fig. 2 Comparison of moisture
content between Monascus-fermented dioscorea and rice (• rice grain; ○ dioscorea)
growth and monacolin K formation time of Monascus with dioscorea.
Monascus-fermented dioscorea is also proven to possess higher production of yellow pigment, besides monacolin K, in contrast with other Monascus-fermented product. From the discussion shown above, we can conclude that dioscorea is better than other substrates when stimulating the formation of monacolin K and yellow pigment of Monascus spp, and dioscorea may possess special ingredients or characters that will help the formation of yellow pigments and monacolin K. Furthermore, both monacolin K and yellow pigment are observably increased via Monascus fermentation with dioscorea, which may indicate that the two compounds have a reciprocal effect on biosynthesis. It should be further investigated whether monacolin K increased as yellow pigment increased or not. In general, yellow pigment produced by Monascus includes monascin and ankaflavin, which have been shown as an anticancer agent in addition to being used as a colorant (Su et al. 2005). In this study, the yellow pigment of Monascus-fermented dioscorea is demon-strated as monascin by identification of LC/MS. Monascin has been widely used as natural food colorant in East Asia for a long time ago particularly for Chinese food and cosmetics (Jongrungruangchok et al. 2004). Martinkova et al. (1999) confirmed that monascin and its derivatives are not toxic. A previous
study reported that monascin possessed mild antibiotic activity against Bacillussubtilis and Candidapseudotro-picalis, and also showed immunosuppressive activity on mouse T-splenocytes (Jongrungruangchok et al. 2004; Martinkova et al. 1999). In a current study, Akihisa et al. (2005) demonstrate that monascin can exhibit potent inhibitory effects on TPA-induced inflammation in mice, the induction of Epstein–Barr virus early antigen (EBV-EA) by TPA in Raji cells, and the activation of (+/ −)-(E)-methyl-2[(E)-hydroxy-imino]-5-nitro-6-methoxy-3-hexemide (NOR 1) known as a nitric oxide (NO) donor. This fact will be a great benefit to the future development of Monascus func-tional food.
Dioscorea species is a member of the monocotyledonous family Dioscoreaceae. Dioscorea may possess some health benefits. More than 70% of starch existed in the dried dioscorea. Amylose is the major type of starch in dioscorea, and its content in the dioscorea is more than that in the rice grain (Wanasundera and Ravindran 1994). In general, amylose molecules is consist of single mostly-unbranched chains with 500–20,000 α-(1-4)-D-glucose units dependent on source was easy to decompose and use by microorgan-ism (Hoover 2001). This character may result in a comfortable environment for Monascus species to grow. In addition to being the substrate of Monascus, dioscorea has been used as the substrate for fermentation of various microorganisms in current studies (Aderiye and Ogunjobi RT:0.00 - 49.99 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42 44 46 48 Time (min) 0 20 40 60 80 100 0 20 40 60 80 100 R e la ti v e A b unda nc e 40.50 0.30 2.01 4.986.637.05 8.999.92 12.94 14.66 4.79 7.21 10.73 15.3416.49 18.45 20.27 22.66 24.27 26.5428.5528.89 31.1433.17 34.42 36.55 38.66 43.4744.1045.82 47.89 3.25 48.89 40.41 6.61 6.49 21.55 4.89 10.23 34.2134.81 4.65 8.369.95 10.6212.4813.8314.47 16.83 18.72 22.07 37.02 39.49 48.28 NL: 2.72E7 TIC MS Pan3 NL: 2.70E-1 uv Analog Pan3
Pan3#2377 RT:40.41 AV:1 NL: 1.31E7
T: + c ESI Full ms [ 125.00-1000.00] 200 300 400 500 600 700 800 900 1000 m/z 0 20 40 60 80 100 R e la ti v e A bun danc e 359.0 360.0 421.5 516.8 736.7 844.7 881.0 284.8 215.1 619.3 718.3 949.1 187.2
A
B
C
Fig. 4 Chromatograms of Monascus-fermented dioscorea and mass spectra of selected peaks a HPLC profile at 233 nm, b total ion current (TIC), and c mass spectra of monascin (M+1=359.0) at 40.41 min
1998; Okigbo 2005). Dioscorea is also shown as a beneficial substrate for the growth of microorganisms. Diosgenin, allanton, choline, and essential amino acids are found in dioscorea besides prolific starch. In further work, it requires to be studied whether any of these components can stimulate the formation of secondary metabolites of Monascus species. Araghiniknam et al. (1996) has indicated that dioscorea has antioxidant ability reducing serum lipid peroxidation and prophylaxis of cardiovascular diseases increasing the high-density lipoprotein (HDL) in the serum. In this study, we also found that Monascus-fermented dioscorea can perform a similar or better activity on antioxidation as compared to unfermented dioscorea (data not shown). Aniya et al. (1999) has found an antioxidant-dimerumic acid from Monascus product, which is proven to reduce NO, DPPH, and free radicals.
Thus, dioscorea is a pretty good health food. If it were used as the substrate, it would have additive effect to the health. It should be noted that using dioscorea as fermen-tation substrate is proven to significantly increase not only the cholesterol-lowering agent monacolin K but also an antiinflammation agent monascin, which will be a great benefit to result in Monascus product being a multi-functional food. The result can be used as an important reference for later study concerning the monacolin K and monascin production through fermentation.
References
Aderiye BI, Ogunjobi AA (1998) Fermentation of yam: microbi-ology and sensory evaluation of cooked fermented yam tissues. Plant Foods Hum Nutr 52:49–54
Akihisa T, Tokuda H, Yasukawa K, Ukiya M, Kiyota A, Sakamoto N, Suzuki T, Tanabe N, Nishino H (2005) Azaphilones, furanoisophthalides, and amino acids from the extracts of Monascuspilosus-fermented rice (red-mold rice) and their chemopreventive effects. J Agric Food Chem 53:562–565 Albert AW, Chen J, Springer J (1980) Mevinolin: a highly potent
competitive inhibitor of hydroxymethylglutaryl-coenzyme A reductase and a cholesterol lowering agent. Proc Natl Acad Sci USA 77:3957–3961
Aniya Y, Ohtani II, Higa T, Miyagi C, Gibo H, Shimabukuro M, Nakanish H, Taira J (1999) Dimerumic acid as an antioxidant of the mold, Monascusanka. Free Radic Biol Med 286:999–1004 Araghiniknam M, Chung S, Nelson-White T, Eskelson C, Watson RR (1996) Antioxidant activity of dioscorea and dehydroepi-nandrosterone (DHEA) in older humans. Life Sci 59:147–157 Blanc PJ, Laussac JP, Le Bars J, Le Bars P, Loret MO, Pareilleux A, Prome D, Prome JC, Santerre AL, Goma G (1995a) Characterization of monascidin A from Monascus as citrinin. Int J Food Microbiol 27:201–213
Blanc PJ, Loret MO, Goma G (1995b) Production of citrinin by various species of Monascus. Biotechnol Lett 17:291–294 Broder CU, Koehler PE (1980) Pigments production by
Monascus-purpureus with regard to quality and quantity. J Food Sci 45:567–569
Carel M, Shepherd D (1977) The effect of different nitrogen source on pigment production and sporulation of Monascus sp. in submerged shaken culture. Can J Microbiol 23:1360–1372 Carlson LA (1982) Serum lipids and atherosclerotic disease. In:
Carlson LA, Pernow B (eds) Metabolic risk factors in ischemia cardiovascular disease. Raven, New York, pp 1–16
Dizon EI, Sanchez PC (1984) Potential of Monascus pigment as coloring agent for banana sauce and tocino. Philipp Agric 67:157–166
Endo A (1979) Monacolin K, a new hypocholesterolemic agent produced by Monascus species. J Antibiot 32:852–854 Gibbs SJ, Johnson CS (1991) A PGF NMR experiment for accurate
diffusion and flow studies in the presence of eddy currents. J Magn Reson 93:395–402
Hesseltine CW (1965) Amillenium of fungi, food and fermentation. Mycologia 57:149–197
Hoover R (2001) Composition, molecular structure, and physico-chemical properties of tuber and root starches: a review. Carbohydr Polym 45:253–267
Jongrungruangchok S, Kittakoop P, Yongsmith B, Bavovada R, Tanasupawat S, Lartpornmatulee N, Thebtaranonth Y (2004) Azaphilone pigments from a yellow mutant of the fungus Monascuskaoliang. Phytochemistry 65:2569–2575
Kycko S, Shiho SS, Yoko K, Tamio M (1998) Analytical method for citrinin in Monascus colour. Jpn J Food Chem 5:64–68 Lee CL, Wang JJ, Pan TM (2006) A synchronous analysis method
for detecting citrinin and monacolin K lactone and the acid form of red mold rice. J AOAC Int (in press)
Lee CL, Wang JJ, Tsai TY, Pan TM (2005) In vivo hypolipidemic effects and safety of low dosage Monascus powder in a hamster model of hyperlipidemia. Appl Microbiol Biotechnol (on line) Lin CF (1973) Isolation and culture conditions of Monascus sp. for the production of pigments in a submerged shaken culture. J Ferment Technol 51:407–414
Lin CF, Demain AL (1991) Effect of nutrition of Monascus sp. on formation of red pigment. Appl Microbiol Biotechnol 36:70–75 Lin CF, Lizuka H (1982) Production of extracellular pigment by a mutant of Monascuskaoliang sp nov. Appl Environ Microbiol 43:671–676
Martinkova L, Patakova-Juzlova P, Krent V, Kucerova Z, Havlicek V, Olsovsky P, Hovorka O, Rihova B, Vesely D, Vesela D, Ulrichova J, Prikrylova V (1999) Biological activities of oligoketide pigments of Monascuspurpureus. Food Addit Contam 16:15–24
Okigbo RN (2005) Biological control of postharvest fungal rot of yam (Dioscorea spp.) with Bacillussubtilis. Mycopathologia 159:307–314
Panitz C, Frost P, Kunz B (1991) Pigment- und Biomassebildung von Monascuspurpureus in synthetischen. Medien Bioeng 7:72–75
Pelta MD, Barjat H, Morris GA, Davis AL, Hammond SJ (1998) Pulse sequences for high-resolution diffusion-ordered spectros-copy (HRDOSY). Magn Reson Chem 36:706–714
Rashbaum SA, Barrington EMY (1983) Natural red coloring prepared from an oat substrate. US Patent 4418081
Santerre AL, Queinnec I, Blanc PJ (1995) A fedbath strategy for optimal red pigment production by Monascusruber. Bioprocess Eng 13:245–250
Serizawa N, Nakagawa K, Hamano K, Tsujita Y, Terahara A, Kuwano H (1983) Microbial hydroxylation of ML-236B (compactin and monacolin K (MB-530). J Antibiot 36:604–607 Silvia B, Livia B, Giangiacomo B, Maria B, Manuela R, Matilde M (2002) Production of lovastatin examined by an integrated approach based on chemometrics and DOSY-NMR. Biotechnol Bioeng 80:579–583
Su NW, Lin YL, Lee MH, Ho CY (2005) Ankaflavin from Monascus-fermented red rice exhibits selective cytotoxic effect and induces cell death on Hep G2 cells. J Agric Food Chem 53:1949–1954
Su YC, Wang JJ, Lin TT, Pan TM (2003) Production of the secondary metabolites γ-aminobutyric acid and monacolin K by Monascus. J Ind Microbiol Biotechnol 30:40–46
Wanasundera JPD, Ravindran G (1994) Nutritional assessment of yam (Dioscoreaalata) tubers. Plant Foods Hum Nutr 46:33–39 Wang JJ, Lee CL, Pan TM (2003) Improvement of monacolin K, γ-aminobutyric acid and citrinin production ratio as a function of environmental conditions of Monascuspurpureus NTU 601. J Ind Microbiol Biotechnol 30:669–676
Wang JJ, Lee CL, Pan TM (2004) Modified mutant method for screening low citrinin-producing strains of Monascuspurpureus on rice culture. J Agric Food Chem 52:6977–6982
Wang L, Mo H, Shi H (1999) Production of lovastatin with solid state fermentation of Monascusruber. Zhongguo Kangshengsu Zazhi 24:96–98
Wang UL, Houng JY, Chang HS, Chien HCR, Hsu WH (1998) Selection of drug-resistant mutants Monascuspilosus for enhanced monacolin K production. J Chin Agric Chem Soc 36:192–200
Wong HC, Koehler PE (1981) Production and isolation of an antibiotic from Monascuspurpureus and its relationship to pigment production. J Food Sci 46:589–592
Yoshimura M, Yamanaka S, Mitsugi K, Hirose Y (1975) Production of Monascus pigment in a submerged culture. Agric Biol Chem 39:1789–1795