RSC Advances
This is an Accepted Manuscript, which has been through the
Royal Society of Chemistry peer review process and has been
accepted for publication.
Accepted Manuscripts are published online shortly after
acceptance, before technical editing, formatting and proof reading.
Using this free service, authors can make their results available
to the community, in citable form, before we publish the edited
article. This Accepted Manuscript will be replaced by the edited,
formatted and paginated article as soon as this is available.
You can find more information about Accepted Manuscripts in the
Information for Authors
.
Please note that technical editing may introduce minor changes
to the text and/or graphics, which may alter content. The journal’s
standard
Terms & Conditions
and the
Ethical guidelines
still
apply. In no event shall the Royal Society of Chemistry be held
responsible for any errors or omissions in this Accepted Manuscript
or any consequences arising from the use of any information it
contains.
This article can be cited before page numbers have been issued, to do this please use: H. Lin, Z. Y. Hong, Y. Li and I. Liau, RSC Adv., 2016, DOI: 10.1039/C6RA13073A.
Journal Name
ARTICLE
Received 00th January 20xx, Accepted 00th January 20xx DOI: 10.1039/x0xx00000x www.rsc.org/Fluorescent tracer of dopamine enables selective labelling and
interrogation of dopaminergic amacrine cells in the retina of living
zebrafish
†
Hui-Jen Lin,
aZhen-Yi Hong
,
aYaw-Kuen Li
*
aand Ian Liau
*
abThe retinal dopaminergic amacrine cell plays multiple roles in vision; it optimizes the encoding of visual stimuli, serves as an output of the circadian rhythm, and influences trophic processes. Specific identification of such cells in vivo remains challenging. A fluorescent tracer that mimics natural dopamine would help elucidate the distribution and the functional roles of dopaminergic neurons. Here we report a fluorescent tracer of dopamine for the selective labeling and study of retinal dopaminergic amacrine cells in vivo. This tracer has low toxicity and effective selectivity, and enables fluorescent determination of the evoked release of dopamine and the femtosecond laser ablation of single dopaminergic amacrine cells in living zebrafish. Our approach is readily adaptable by other research groups, and is expected to accelerate biomedical research in domains related to the pathophysiology of retinal dopaminergic amacrine cells.
Introduction
Amacrine cells are interneurons that interact with retinal ganglion cells or bipolar cells. Distinctively, the cell bodies of amacrine cells locate in the inner nuclear layer (INL) of the retina while their dendrites project into the inner plexiform layer (IPL).1-3 Amacrine cells are commonly classified according to not only the morphology of their dendrites but also the neurotransmitters that they express. Among amacrine cells of varied types, the dopaminergic amacrine cell, which is characterized by their expression, release and uptake of dopamine (DA) neurotransmitters, has been shown to possess multiple roles in vision; it optimizes the encoding of visual stimuli at varied levels of illumination, serves as an output of the retinal circadian rhythm, mediates visual processes such as adapting light and dark retinal circuits, and influences trophic processes, such as the survival of photoreceptors and the degree of refractive errors in myopia.3-6 Despite its pathophysiological importance in vision, our understanding towards the function and activity of the dopaminergic amacrine cell remain elusive. In particularly, important mechanistic questions such as how the secretion of retinal DA is regulated, and how illumination controls the activity of the dopaminergic amacrine neurons are largely unanswered.6
Scheme 1 Selective labelling of dopaminergic amacrine cells in the retina of living zebrafish. OPL: outer plexiform layer; INL: inner nuclear layer; IPL: inner plexiform layer; GCL: ganglion cell layer; DAT: dopamine transporter.
The identification of amacrine cells of particular types in vivo is a key step to interrogate their function and activity, and to elucidate their pathophysiological role in vision; however, a technique for this purpose is lacking with regard to dopaminergic neurons.7 Immunohistochemical stains targeting transporters or enzymes that are specific to the transmission and synthesis of particular neurotransmitters are commonly employed to identify amacrine cells in retina.8 In particular, the antibody to tyrosine hydroxylase (TH) has been developed and utilized to label amacrine cells in mice and zebrafish.9-11 The TH enzyme is responsible for catalyzing the conversion of L-tyrosine to L-3,4-dihydroxyphenylalanine (L-DOPA), and can be found in amacrine cells that express catecholamines of varied types; the antibody to TH is hence not specific to dopaminergic
RSC
Advances
Accepted
Manuscript
ARTICLE
Journal Name
amacrine cells. An antibody to dopamine transporter (DAT) has been developed and utilized to label dopaminergic cells in mice,12,13 but its specificity to dopaminergic amacrine cells has not yet been demonstrated. Furthermore, these approaches are inapplicable to living tissues or organisms thereby making them impractical to research that requires the determination of neural activity. On the other hand, transgenic lines of zebrafish that express fluorescent proteins in tyrosine hydroxylase (TH) or dopamine transporter (DAT) expressing amacrine cells have been developed.10,14,15 However, their application to selective labelling remains yet fully demonstrated. In particular, the fluorescent proteins that are co-expressed with either TH or DAT are found not exclusively in dopaminergic cells.8,10,15 Besides, the expression level of fluorescent proteins might be subject to interference due to various conditions; in particular, the green fluorescent protein (GFP) has been reported to express ectopically in some non-dopaminergic amacrine cells.16 Moreover, an interrogation of dopaminergic amacrine cells in the retina of living animals has not yet been demonstrated to the best of our knowledge.
DA neurotransmitters are expressed, released and accumulated by dopaminergic neurons. Combined with sensitive fluorescence imaging, a fluorescently tagged DA that retains the property of natural DA would be a promising strategy to facilitate research on the neurotransmission of dopaminergic neurons at the level of single cells. Towards this goal, we have developed a fluorescent tracer of DA by direct conjugating a fluorescent dye to DA neurotransmitter; we showed that such ‘fluorescent dopamine’ retains numerous essential features of natural DA:17 it accumulates abundantly in dopaminergic cells but not cells of other -ergic types; its uptake is mediated mainly by DAT; it is evoked to release by potassium ion at a high concentration, and it enables specific fluorescent labelling of dopaminergic PC12 cells in vitro and selective mapping of regions abundant in dopaminergic innervations in acute brain slices of rats.
In light of the preceding results, we further postulated that such ‘fluorescent dopamine’ can specifically label dopaminergic amacrine cells in the retina in vivo. To test this hypothesis, we employed zebrafish as a model. The zebrafish possesses numerous attractive features such as ease of maintenance, rapid development, availability of forward and reverse genetic methods for phenotypical and genetic analysis,18-21 and has emerged as a popular model organism to investigate neural science and the pathophysiology of vision.22,23 In particular, the zebrafish has been employed to study retinal diseases,24 such as age-related macular degeneration, retinitis pigmentosa and diabetic retinopathy.25 Its small and translucent larval body further provides an unprecedented opportunity to interrogate individual cells in vivo with optical means. For the purpose of demonstration, we reveal the fluorescent determination of DA, which is stimulated to become released from dopaminergic amacrine cells; guided with fluorescence imaging, we selectively ablated single dopaminergic amacrine cells with femtosecond laser pulses. We envisage that our work will help to elucidate the pathophysiology central to dopaminergic amacrine cells.
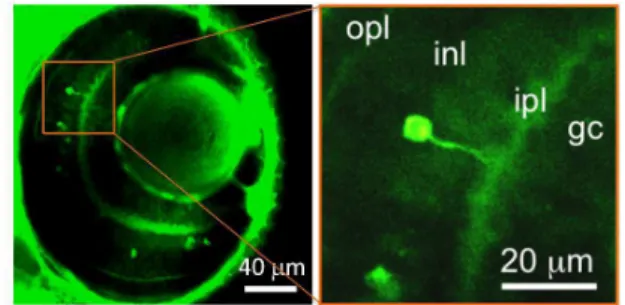
Fig. 1 Accumulation of FITC-DA in cells that possess features consistent with the characteristics of retinal amacrine cells. Representative (left) and magnified (right) confocal fluorescent images acquired from a z-section show that some cells are distinctly labeled with FITC-DA and these cells are located in the INL with the dendrite protruding into the IPL.
Results and discussion
FITC-DA exhibits low toxicity for larval zebrafish
An ideal fluorescent tracer for living organisms would impose little toxicity on the organism. To assess the toxicity of FITC-DA, we tested whether the treatment of FITC-DA might impair the growth of larval zebrafish. Larval zebrafish (wild type) at three and four days post fertilization (dpf) were incubated in a solution of FITC-DA (2 mM) for varied durations (1, 2, 3 and 4 h); their growth was observed for another four days. Our results show that these larvae continued to grow normally, similarly to the untreated control group, with nearly all larvae surviving (ESI Fig. S1†). Such a result demonstrates that incubation of FITC-DA imposed negligible toxicity on larval zebrafish and did not impair their growth.
FITC-DA accumulates in cells with features conforming to retinal amacrine cells
An initial evaluation of the fluorescent tracer of DA was conducted on incubating larval zebrafish (4 dpf) with FITC-DA (2 mM) for 60 min. A representative confocal image acquired near the eye of larval zebrafish showed that several cells exhibited strong fluorescence, which indicated that these cells were distinctively labelled with FITC-DA (left, Fig. 1). A magnified image of a cell revealed further marked features: the cell exhibited neuron-like morphology possessing a dendrite; moreover, the cell is located in the INL with its dendrite protruding into the IPL (right, Fig. 1). These findings are consistent with the characteristics of amacrine cells as described.1-3
FITC-DA selectively labels dopaminergic amacrine neurons in the retina of living zebrafish
We investigated the specificity of FITC-DA with immunofluorescence imaging according to the following protocol. A larva (4 dpf) was firsts soaked with FITC-DA (2 mM) for 60 min; confocal images near the eye of the larva were acquired at varied z-depths. The larva was then fixed, labelled sequentially with the primary antibody of TH and a fluorescently tagged secondary antibody, and imaged.
RSC
Advances
Accepted
Manuscript
Published on 20 July 2016. Downloaded by RMIT University Library on 22/07/2016 03:34:31.
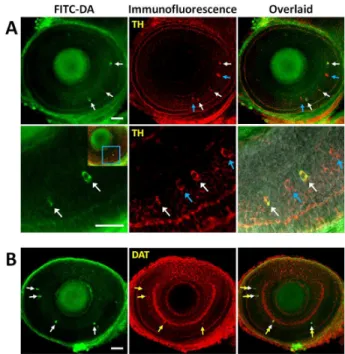
Comparison of the immunofluorescence image of TH with the fluorescence image of FITC-DA and inspection of the overlaid image showed that all FITC-DA positive cells colocalized with TH positive cells (white arrows), whereas some TH- positive cells were not labelled with FITC-DA (blue arrows) (Fig. 2A). As described, TH is responsible for the conversion of L-tyrosine to L-DOPA, and are expected to be present in cells that express catecholamines but not exclusive to dopaminergic neurons.8 Consistent to this notion, our result shows clearly that FITC-DA labelled catecholaminergic amacrine cells, and some but not all catecholaminergic cells were labelled with FITC-DA. We proceeded to verify the selective labelling of dopaminergic cells with FITC-DA according to the following protocol. A larva was first soaked with FITC-DA, and then labelled with immunofluorescence stains targeting DAT. The result showed that the fluorescence of FITC-DA colocalized with the immunofluorescence of DAT (white and yellow arrow, Figure 2B).
With these results taken together, we conclude that FITC-DA accumulated selectively in retinal dopaminergic amacrine cells of living larval zebrafish. At present, antibodies specific to transporters of neurotransmitters (such as DAT for dopaminergic neurons),26 or to enzymes responsible for the synthesis of neurotransmitters (such as TH for catecholaminergic neurons)8,27 are commonly employed as markers of specific neuronal populations in retina. The procedure of immunohistochemistry, however, inevitably impairs the integrity of retinas, thereby preventing tests of physiological activities on neuronal cells. In comparison, our approach is advantageous as it not only requires minimum sample preparation but also allows noninvasive and selective fluorescence labelling of dopaminergic amacrine cells.
The neurotransmitter moiety of the FITC-DA conjugate is crucial for the selective labelling of dopaminergic amacrine cells
The preceding result indicates that the conjugate of DA and FITC retained the essential property of DA, and was shown to accumulate selectively in dopaminergic amacrine cells in vivo. We suggest that such selective accumulation was mediated primarily by the neurotransmitter moiety of the conjugate rather than the FITC moiety. To verify our deduction, we synthesized fluorescent conjugates of two monoamines, benzylamine (BA) and cyclohexylamine (CA), and verified their property in a way similar to that with FITC-DA. These monoamines have structures similar to that of DA, but are not neurotransmitters; their fluorescent conjugates hence served as a negative control for the verification of our deduction. As expected, the uptake of the fluorescent conjugates of these two non-neurotransmitter monoamines (FITC-BA and FITC-CA) by retinal amacrine cells was insignificant (Fig. 3). Such a result hence supports our notion that the DA moiety of the FITC-DA conjugate plays an essential role in the selective accumulation of the conjugate in the dopaminergic amacrine cells.
Fig. 2 Confirmation of the selective labelling of dopaminergic amacrine neurons in the retina of larval zebrafish. (A) Upper panel: fluorescence, immunofluorescence images and their overlay show that all FITC-DA positive cells colocalized with some but not all TH positive cells (scale bar: 40 µm). Lower panel: magnified images (scale bar: 20 µm). The white and blue arrows indicate FITC-DA and TH positive cells, respectively. (B) Results show that all FITC-DA colocalized with DAT positive cells (scale bar: 40 µm). The white and yellow arrows indicate FITC-DA and DAT positive cells, respectively.
Fig. 3 Results validating that the dopaminergic moiety plays an essential in the selective uptake of FITC-DA by dopaminergic amacrine cells. The images show that neither FITC-BA (lefT) nor FITC-CA (right) accumulated in the
dopaminergic amacrine cell of larval zebrafish (4 dpf). Scale bars: 40 µm.
RSC
Advances
Accepted
Manuscript
ARTICLE
Journal Name
Fig. 4 Fluorescent determination of the secretion of dopamine from single retinal dopaminergic amacrine cells. (A) Upper panel: time-course images demonstrate a significant decrease of the fluorescent intensity of a dopaminergic amacrine cell (white arrow) after an injection of high potassium to the orbital of larval zebrafish. Low panel: results of a sham experiment prepared on an injection of PBS. Scale bars: 10 µm. (B) Temporal change of the fluorescence intensity shows that rapid decline of the fluorescent intensity induced by the injection of potassium ion (red: KCl; blue: PBS; black: untreated; n = 5).
FITC-DA enables fluorescence detection of the evoked release of DA from single retinal dopaminergic amacrine cells Having shown that FITC-DA was selectively accumulated dopaminergic cells in larval zebrafish, we expected that ‘fluorescent dopamine’ would enable fluorescence imaging and detection of the secretion of DA neurotransmission from single dopaminergic amacrine cells in vivo.
As a demonstration, we assessed the evoked release of DA at the level of single cells in living zebrafish with fluorescence detection. A larval zebrafish (4 dpf) was soaked with FITC-DA (2 mM) for 60 min, and injected with a solution of potassium ion with concentration significantly higher than the intracellular concentration of potassium in freshwater fish (140 mM vs. 90 mM). After injection, time-course images were acquired; the fluorescence intensity of FITC-DA was analyzed to determine the temporal change of the intracellular centration of DA.
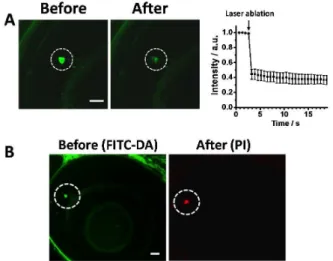
The time-course images and quantitative analysis of the images showed that injection of potassium caused a significant decrease in fluorescence of the cells labelled with FITC-DA soon after the injection, which indicates an secretion of DA evoked by potassium ion; in contrast, the untreated control and the sham experiment, which was prepared on injecting PBS ([K+] = 27 mM), resulted in insignificant change in the fluorescence (Fig. 4). Such a finding intricately demonstrates the unique ability of our approach to determine quantitatively the evoked release of DA neurotransmitters from single dopaminergic amacrine cells in living zebrafish.
Fig. 5 Femtosecond laser ablation of single dopaminergic amacrine cells facilitated with selective labeling of dopaminergic neurons. (A) Left: images of the target cell (while dotted circle) acquired before and approximately 0.65 s after the ablation. Right: the temporal change of the fluorescence intensity shows a dramatic decline of the fluorescence intensity immediately after ablation. n = 3. (B) Fluorescence images of FITC-DA (left) and PI (after) acquired before and after the ablation of a cell show that the target cell (whilte dotted circle) was ablated and become dead. Scale bars: 20 µm.
Femtosecond-laser ablation of single dopaminergic amacrine cell in vivo
The unique ability of our ‘fluorescent dopamine’ provides a new opportunity to interrogate single dopaminergic amacrine cells in vivo, in particular through the femtosecond-laser ablation of single cells. To demonstrate this ability, we first treated a larval zebrafish with FITC-DA using the protocol described above, and positioned a dopaminergic amacrine cell at the focus of the ablation laser under the guidance of fluorescence imaging. After a brief exposure to the laser illumination, the fluorescence of the target cell became dimmed with the intensity decreasing by approximately 40 % relative to the baseline (Fig. 5A). In another test, we employed propidium iodide (PI), a common indicator of dead cells,28 to verify the viability of cells after laser ablation. The fluorescence image acquired after ablation showed that only the target cell was selectively labelled with PI, whereas cells adjacent to the target cells showed negligible PI fluorescence (Fig. 5B). These results demonstrated that, guided with fluorescence imaging of FITC-DA, the target dopaminergic amacrine cell was selectively ablated with the adjacent cells remaining intact.
Conclusions
FITC-DA selectively accumulates in dopaminergic amacrine cells in the retina of living zebrafish. Our work provides a prospective tool for the selective labeling of dopaminergic neurons, and enables both quantitative imaging of the evoked release of dopamine and interrogation of single dopaminergic neurons in vivo. We are progressing with evaluation of FITC-DA
RSC
Advances
Accepted
Manuscript
Published on 20 July 2016. Downloaded by RMIT University Library on 22/07/2016 03:34:31.
as a novel fluorescent tracer of DA neurotransmission using zebrafish as a model.
Materials and methods
All experiments were performed in compliance with the relevant laws and institutional guidelines and approved by the Animal Investigation Committee of National Chiao Tung University.
Maintenance and handling of zebrafish
The AB strain zebrafish (Danio rerio) were provided by Taiwan Zebrafish Core Facility and maintained according to protocols previously reported.29 To inhibit the formation of melanophores, PTU (Sigma, Aldrich) was added to the egg medium beginning from 24 h post fertilization. For anesthetization, zebrafish larvae were immersed with a mixture of tricaine (300 mg/L) (Sigma, Aldrich) and isoflurane (100 ppm) (Alfa Aesar).30 Larval zebrafish at ages 3 and 4 dpf were used throughout this work.
Synthesis and characterization of fluorescent tracers
Details of the synthesis, purification and characterization of ‘fluorescent dopamine’ are reported elsewhere.17 In brief, the fluorescent tracer of DA (FITC-DA) was synthesized through conjugation of the isothiocyanate reactive group (-N=C=S) of FITC (Sigma) to the primary amine group of DA (Alfa Asesar). The FITC conjugates of BA (Sigma Aldrich) and CA (Sigma Aldrich) were synthesized in the same manner. The purity of the products was verified with electrospray-ionization mass spectra (ESI-MS), and 1H and 13C unclear magnetic resonance (NMR) spectra.
Labelling of zebrafish with fluorescent tracers
Zebrafish larvae were soaked in a solution containing the indicated fluorescent tracers (2 mM in DPBS buffer) for 60 min. The control was prepared on soaking larvae in a DPBS buffer containing no fluorescent analogues for the same duration. Whole mount immunohistochemical fluorescent staining To verify the specificity of FITC-DA, we first soaked larval zebrafish (4 dpf) with FITC-DA, washed three times with water, and fixed in paraformaldehyde (4%, Sigma Aldrich) overnight. Second, the larvae were washed at least four times in PBS with Triton X-100 (1%, Sigma), permeabilized with ice-cold acetone (J. T. Baker) for 8 min, and incubated in a blocking solution (Visual Protein Biotech Corp.) overnight. The larvae were then transferred into a blocking solution containing anti-TH antibody (1:300, mouse monoclonal, MAB318; Merck Millipore) or anti-dopamine transporter antibody (1:100, rabbit polyclonal, AB1766; Merck Millipore), at 4 °C for three days. After intensive washing with PBS, larval zebrafish were incubated with a blocking solution containing the secondary antibodies, goat anti-mouse IgG conjugated with Alexa-647 (Life Technologies) or goat anti-rabbit IgG conjugated with Alexa-647 (Life Technologies), at 4 °C for 24 h.
Evoked released of FITC-DA from dopaminergic amacrine cells in zebrafish
Larval zebrafish were first loaded with FITC-DA (2 mM) for 60 min. A PBS solution of potassium ions (140 mM) was then injected into the orbital of larvae with a micro-injector (Eppendorf FemtoJet). The sham experiment was performed on injection of a PBS solution containing potassium ion of 27 mM in concentration.
Femtosecond -laser ablation of single dopaminergic amacrine cells in zebrafish
The experimental setup (ESI Fig. S2†) for the femtosecond-laser ablation was modified from a confocal microscope (TCS SP5II, Leica). The beam of a femtosecond-pulsed laser (Tsunami, Spectra-Physics; pulse width 150 fs, typical power 340 mW measured before the objective lens) was directed to the microscope through the back port of the microscope, and focused with a water-immersion objective lens (HC PL Apo CS2 63XW, NA 1.2, Leica). The target cell was identified from confocal fluorescent images and positioned with a translation stage to the laser focus. The duration of irradiation, typically set at 0.01 s, was controlled with a mechanical shutter (Sutter Instrument Co.).
To test the viability of cells after ablation, the ablated larval was incubated in a solution containing PI (100 µg/mL; Sigma) for 20 min and imaged with a confocal microscope.
Fluorescent and bright-field imaging
Fluorescent and bright-field images were acquired simultaneously with a laser scanning confocal microscope (TCS SP5 II, Leica). All images shown in this work were produced on averaging results of three consecutive scans.
To produce fluorescent images of FITC-DA, light from a blue laser (488 nm; typical power below 1 mW) was employed as excitation; emission between 495 and 570 nm was collected. To colocalize the emission of FITC-DA and fluorescently labeled secondary antibodies, the blue laser and a red laser (633 nm; typical power less than 1 mW) were used as excitation; emissions between 495 and 600 nm (FITC-DA) and between 650 and 760 nm (Alexa Fluor647) were collected. To examine the viability of cells, we used light from the blue laser and an orange laser (561 nm; typical power below 1 mW) as excitation; emissions between 495 and 555 nm (FITC-DA) and between 620 and 720 nm (PI) were collected.
Quantitative analysis of images
Analysis of the image was performed with ImageJ (National Institutes of Health, USA). Individual cells were first identified as a region of interest (ROI). The intensity per cell was determined on integrating the intensity of the ROI.
Statistical analysis
All data are expressed as mean ± S.E.M. Comparison between the means of two groups was made using the two-tailed Student’s t test. The levels of statistical significance were set at *P < 0.05, **P < 0.01, and ***P < 0.001, respectively.
RSC
Advances
Accepted
Manuscript
ARTICLE
Journal Name
Acknowledgements
We thank Professor Yuan-Pern Lee (National Chiao Tung University) for generous support and Professor Tzai-Wen Chiu (National Chiao Tung University) for critical suggestion and comments. Ministry of Science and Technology and the MOE-ATU program of Taiwan provided support to I.L.
Notes and references
1. Arenzana, F. J., Arevalo, R., Sanchez-Gonzalez, R., Clemente, D., Aijon, J., Porteros, A. Tyrosine hydroxylase immunoreactivity in the developing visual pathway of the zebrafish. Anatomy and Embryology 2006, 211, 323-334. 2. Cederlund, M. L., Morrissey, M. E., Baden, T., Scholz, D.,
Vendrell, V., Lagnado, L., Connaughton, V. P., Kennedy, B. N. Zebrafish Tg(7.2mab21l2:EGFP)ucd2 transgenics reveal a unique population of retinal amacrine cells. Investigative Ophthalmology & Visual Science 2011, 52, 1613-1621. 3. Witkovsky, P. Dopamine and retinal function. Documenta
Ophthalmologica 2004, 108, 17-40.
4. Djamgoz, M. B. A., Wagner, H. J. Localization and function of dopamine in the adult vertebrate retina. Neurochemistry International 1992, 20, 139-191.
5. Zhang, D.-Q., Wong, K. Y., Sollars, P. J., Berson, D. M., Pickard, G. E., McMahon, D. G. Intraretinal signaling by ganglion cell photoreceptors to dopaminergic amacrine neurons. Proceedings of the National Academy of Sciences of the United States of America 2008, 105, 14181-14186. 6. Zhang, D.-Q., Zhou, T.-R., McMahon, D. G. Functional
heterogeneity of retinal dopaminergic neurons underlying their multiple roles in vision. Journal of Neuroscience 2007, 27, 692-699.
7. Versauxbotteri, C., Nguyenlegros, J., Vigny, A., Raoux, N. Morphology, density and distribution of tyrosine hydroxylase-like immunoreactive cells in the retina of mice. Brain Research 1984, 301, 192-197.
8. Holzschuh, J., Ryu, S., Aberger, F., Driever, W. Dopamine transporter expression distinguishes dopaminergic neurons from other catecholaminergic neurons in the developing zebrafish embryo. Mechanisms of Development 2001, 101, 237-243.
9. Debertin, G., Kantor, O., Kovacs-Oeller, T., Balogh, L., Szabo-Meleg, E., Orban, J., Nyitrai, M., Voelgyi, B. Tyrosine hydroxylase positive perisomatic rings are formed around various amacrine cell types in the mammalian retina. Journal of Neurochemistry 2015, 134, 416-428.
10. Meng, S., Ryu, S., Zhao, B., Zhang, D.-Q., Driever, W., McMahon, D. G. Targeting retinal dopaminergic neurons in tyrosine hydroxylase-driven green fluorescent protein transgenic zebrafish. Molecular Vision 2008, 14, 2475-2483. 11. Filippi, A., Duerr, K., Ryu, S., Willaredt, M., Holzschuh, J.,
Driever, W. Expression and function of nr4a2, lmx1b, and pitx3 in zebrafish dopaminergic and noradrenergic neuronal development. Bmc Developmental Biology 2007, 7.
12. Kurosaki, R., Muramatsu, Y., Watanabe, H., Michimata, M., Matsubara, M., Imai, Y., Araki, T. Role of dopamine
transporter against MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) neurotoxicity in mice. Metabolic Brain Disease 2003, 18, 139-146.
13. Li, Z. S., Pham, T. D., Tamir, H., Chen, J. J., Gershon, M. D. Enteric dopaminergic neurons: definition, developmental lineage, and effects of extrinsic denervation. Journal of Neuroscience 2004, 24, 1330-1339.
14. Zhang, D. Q., Stone, J. F., Zhou, T. R., Ohta, H., McMahon, D. G. Characterization of genetically labeled catecholamine neurons in the mouse retina. Neuroreport 2004, 15, 1761-1765.
15. Noble, S., Godoy, R., Affaticati, P., Ekker, M. Transgenic zebrafish expressing mCherry in the mitochondria of dopaminergic neurons. Zebrafish 2015, 12, 349-356. 16. Matsushita, N., Okada, H., Yasoshima, Y., Takahashi, K.,
Kiuchi, K., Kobayashi, K. Dynamics of tyrosine hydroxylase promoter activity during midbrain dopaminergic neuron development. Journal of Neurochemistry 2002, 82, 295-304. 17. Lin, H.-J., Lu, H.-H., Liu, K.-M., Chau, C.-M., Hsieh, Z., Li,
Y.-K., Liau, I. Toward live-cell imaging of dopamine neurotransmission with fluorescent neurotransmitter analogues. Chemical Communications 2015, 51, 14080-14083.
18. Goldsmith, J. R., Jobin, C. Think small: zebrafish as a model system of human pathology. Journal of Biomedicine and Biotechnology 2012.
19. Neelkantan, N., Mikhaylova, A., Stewart, A. M., Arnold, R., Gjeloshi, V., Kondaveeti, D., Poudel, M. K., Kalueff, A. V. Perspectives on zebrafish models of hallucinogenic drugs and related psychotropic compounds. Acs Chemical Neuroscience 2013, 4, 1137-1150.
20. Siebel, A. M., Vianna, M. R., Bonan, C. D. Pharmacological and toxicological effects of lithium in zebrafish. Acs Chemical Neuroscience 2014, 5, 468-476.
21. Chatterjee, P. K., Shakes, L. A., Wolf, H. M., Mujalled, M. A., Zhou, C., Hatcher, C., Norford, D. C. Identifying distal cis-acting gene-regulatory sequences by expressing BACs functionalized with loxP-Tn10 transposons in zebrafish. Rsc Advances 2013, 3, 8604-8617.
22. Fadool, J. M., Dowling, J. E. Zebrafish: a model system for the study of eye genetics. Progress in Retinal and Eye Research 2008, 27, 89-110.
23. Bibliowicz, J., Tittle, R. K., Gross, J. M. Toward a better understanding of human eye disease insights from the zebrafish, Danio rerio. in Animal Models of Human Disease, Vol. 100 (eds. Chang, K. T., Min, K. T.) 287-330 (2011). 24. Gestri, G., Link, B. A., Neuhauss, S. C. F. The visual system of
zebrafish and its use to model human ocular diseases. Developmental Neurobiology 2012, 72, 302-327.
25. Chhetri, J., Jacobson, G., Gueven, N. Zebrafish-on the move towards ophthalmological research. Eye 2014, 28, 367-380. 26. Ciliax, B. J., Heilman, C., Demchyshyn, L. L., Pristupa, Z. B.,
Ince, E., Hersch, S. M., Niznik, H. B., Levey, A. I. The dopamine transporter: immunochemical characterization and localization in brain. Journal of Neuroscience 1995, 15, 1714-1723.
RSC
Advances
Accepted
Manuscript
Published on 20 July 2016. Downloaded by RMIT University Library on 22/07/2016 03:34:31.
27. Schweitzer, J., Lohr, H., Filippi, A., Driever, W. Dopaminergic and noradrenergic circuit development in zebrafish. Developmental Neurobiology 2012, 72, 256-268.
28. Riccardi, C., Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nature Protocols 2006, 1, 1458-1461.
29. Huang, S.-H., Hsiao, C.-D., Lin, D.-S., Chow, C.-Y., Chang, C.-J., Liau, I. Imaging of zebrafish in vivo with second-harmonic generation reveals shortened sarcomeres associated with myopathy induced by statin. Plos One 2011, 6, e24764. 30. Huang, W.-C., Hsieh, Y.-S., Chen, I. H., Wang, C.-H., Chang,
H.-W., Yang, C.-C., Ku, T.-H., Yeh, S.-R., Chuang, Y.-J. Combined use of MS-222 (tricaine) and isoflurane extends anesthesia time and minimizes cardiac rhythm side effects in adult zebrafish. Zebrafish 2010, 7, 297-304.