SKF 83959對時距有關的操作式制約行為之影響效果及其神經機制 - 政大學術集成
140
0
0
全文
(2) 摘要 近年來與多巴胺功能相關的研究,除探討各型多巴胺受體引發細胞內訊息傳遞機 制,亦開始檢測其次級受體之間如何互動及其功能作用為何。近期研究在大腦中的紋狀體 發現了由第一型多巴胺受體(D1R)與第二型多巴胺受體(D2R)所共同組成的雜二聚體 (heterodimer),其活化會激發一連串有別於傳統多巴胺傳導的訊息傳遞路徑,其中包括 經磷脂酶 C 調控的磷酸肌醇之水解,以及後續的細胞內鈣離子釋放。目前,此 D1-D2 雜 二聚體在活體(in vivo)層級的功能不明,仍尚待釐清。因此,本研究以一個屬於 D1-D2 雜二聚體致效劑的 SKF 83959 藥物,檢測其對大鼠於兩項與時距有關的操作式制約行為. 治 政 特定蛋白質表現受此藥物行為作用之影響。本研究結果發現,隨著 SKF 83959 藥物處理 大 立 的劑量增加,大鼠於 FI 30-s 以及 DRL 10-s 作業上的反應率受到了顯著的降低,然而自發. 表現(FI 30-s 與 DRL 10-s)和自發性活動量之影響,並測量四個大腦多巴胺相關區域的. ‧ 國. 學. 性活動量並未受到藥物效果影響。此項結果顯示 SKF 83959 可能對獲取酬賞物相關的內 在動機歷程有影響,而不是因藥物引起運動失能之效。在生化測試蛋白質表現量的結果,. ‧. SKF 83959 對背側紋狀體以及伏隔核中的 pCaMKII、PKA、及 pCREB 引發了較明顯的蛋. y. Nat. 白質表現量變化,在前額葉與海馬迴則未有此效。本研究另外嘗試藉由藥理拮抗的實驗,. io. sit. 檢測多巴胺受器拮抗劑是否能反轉 SKF 83959 藥物對行為表現。實驗結果大致顯示單獨. n. al. er. 施打 SCH 23390 以及共同施打 SCH 23390 與 eticlopride 的前處理,無法反轉 SKF83959 對. i n U. v. 於操作式制約行為表現的影響;然而,低劑量的 eticlopride 對回復 SKF 83959 所引發的低. Ch. engchi. 反應率有部份藥理反轉的效果。綜合以上結果,SKF 83959 會有不等程度的影響本研究所 採之兩種時距有關的操作式制約行為,其可能涉及大腦紋狀體內 CaMKII/CREB 的生化作 用。未來研究可考慮直接操弄細胞內蛋白質的表現量,或者觀測 D1-D2 雜二聚體於活體內 的活動,以提供更多關於此 D1-D2 雜二聚體參與個體的行為功能與其相關神經機制。 關鍵字:多巴胺受體、D1-D2 雜二聚體、SKF 83959、操作式制約行為、FI 30 秒作 業、DRL 10 秒作業. I.
(3) Abstract As the functions and signaling mechanisms of dopamine (DA) receptor subtypes remain popular topics of research, recent studies have also begun to investigate the interactions between different subtypes of receptors. The formation of DA D1-D2 receptor complexes was discovered in the striatum, whose activation leads to a novel signaling pathway via phospholipase Cmediated phosphoinositide hydrolysis, followed by intracellular calcium release. As the in vivo functional role of this D1-D2 receptor heteromer remains to be elucidated, the present study. 治 政 大 10-s), locomotor activity, and performance of schedule-controlled behaviors (FI 30-s and DRL 立 investigated the effects of SKF 83959, a proposed D1-D2 heteromer-selective agonist, on the. the expression of related proteins in four terminals of the mesocorticolimbic DA system, which. ‧ 國. 學. included the prefrontal cortex (PFC), dorsal striatum (DS), nucleus accumbens (NAc), and dorsal. ‧. hippocampus. The administration of SKF 83959 was found to reduce the response rates of FI 30s and DRL 10-s in a dose-dependent manner, whereas the locomotor activity was not affected.. y. Nat. er. io. sit. This suggests that SKF 83959 may have affected the processes of intrinsic motivation to obtain the reinforcers, rather than motor control. In respective to protein expression, SKF 83959. n. al. Ch. i n U. v. induced prominent changes in the levels of pCaMKII, PKA, and pCREB in the DS and NAc. engchi. relative to the PFC and hippocampus. Experiments of pharmacological antagonism were conducted in attempts to reverse the behavioral effects of SKF 83959. The results showed that the pretreatments of SCH 23390 alone and SCH 23390 combined with eticlopride did not reverse the effects of SKF 83959 on operant behaviors. However, low dose eticlopride appeared to have a partial effect in restoring the decline in operant response rates by SKF 83959. Together, the current data showed that SKF 83959 altered the time-based operant behaviors tested to different degrees, possibly via its influence on CaMKII-CREB signaling in the NAc. Future studies that. II.
(4) manipulate the activation of intracellular proteins or quantify the levels of D1-D2 heteromer activation may provide more information regarding the in vivo activation mechanisms of D1-D2 heteromers. Keywords: dopamine receptor, D1-D2 heteromer, SKF 83959, operant behaviors, FI 30 sec schedule, DRL 10 sec schedule.. 立. 政 治 大. ‧. ‧ 國. 學. n. er. io. sit. y. Nat. al. Ch. engchi. III. i n U. v.
(5) Contents 摘要.................................................................................................................................................. I Abstract ........................................................................................................................................... II Contents ........................................................................................................................................ IV List of Tables ................................................................................................................................ VI List of Figures .............................................................................................................................. VII Introduction ......................................................................................................................................1. 政 治 大 DA Receptors and Signaling Pathways ................................................................................... 3 立. The Behavioral Functions of DA: A Historical Overview ...................................................... 1. DA Transmission Pathways..................................................................................................... 5. ‧ 國. 學. The Discovery from D1R and D2R Co-localization to D1-D2 Receptor Heteromers ............... 7. ‧. SKF 83959 as a Proposed Agonist for D1-D2 Receptor Heteromers ....................................... 9 DA and Schedule-Controlled Behaviors ............................................................................... 12. y. Nat. io. sit. The Operant Behavioral Pharmacology of DA-Related Drugs ............................................. 15. n. al. er. Objective of the Present Study .............................................................................................. 17. i n U. v. Methods..........................................................................................................................................20. Ch. engchi. Subjects .................................................................................................................................. 20 Apparatus ............................................................................................................................... 20 Training of FI and DRL Behaviors........................................................................................ 22 Drugs ..................................................................................................................................... 22 Western Blot .......................................................................................................................... 23 Procedures ............................................................................................................................. 24 Statistical Analysis ................................................................................................................ 27 Results ............................................................................................................................................28. IV.
(6) Experiment 1: The Dose Effects of SKF 83959 on FI 30-s and DRL 10-s Performance ..... 28 Experiment 2: The Effects of SKF 83959 on Operant Performance and Levels of Selected Protein Expression in Mesocorticolimbic DA Terminals...................................................... 29 Experiment 3: The Effects of SKF 83959 on Locomotior Activity ...................................... 35 Experiment 4: The Effects of SCH 23390 and Eticlopride Pretreatment on SKF 83959Induced Operant Performance ............................................................................................... 36 Discussion ......................................................................................................................................46 SKF 83959 on Operant Behavioral Performance: A Comparison with Other DA Agents ... 47 The Involvement of Brain Regions in the Operant Behavioral Changes Induced by SKF 83959 ..................................................................................................................................... 53. 政 治 大 The Involvement of Signaling Proteins in the Operant Behavioral Changes Induced by SKF 立 83959 ..................................................................................................................................... 58. ‧ 國. 學. DAR Antagonists and the Operant Behavioral Changes Induced by SKF 83959................. 64 Study Limitations .................................................................................................................. 67. ‧. Suggestions for Future Studies .............................................................................................. 69. y. Nat. sit. Conclusion ............................................................................................................................. 70. n. al. er. io. References ......................................................................................................................................71. i n U. v. Tables………………………………….………………………………………………………………….92. Ch. engchi. Figures……………………………………………………………………………………………………97 Appendix…………………………………………………………………………………………..……130. V.
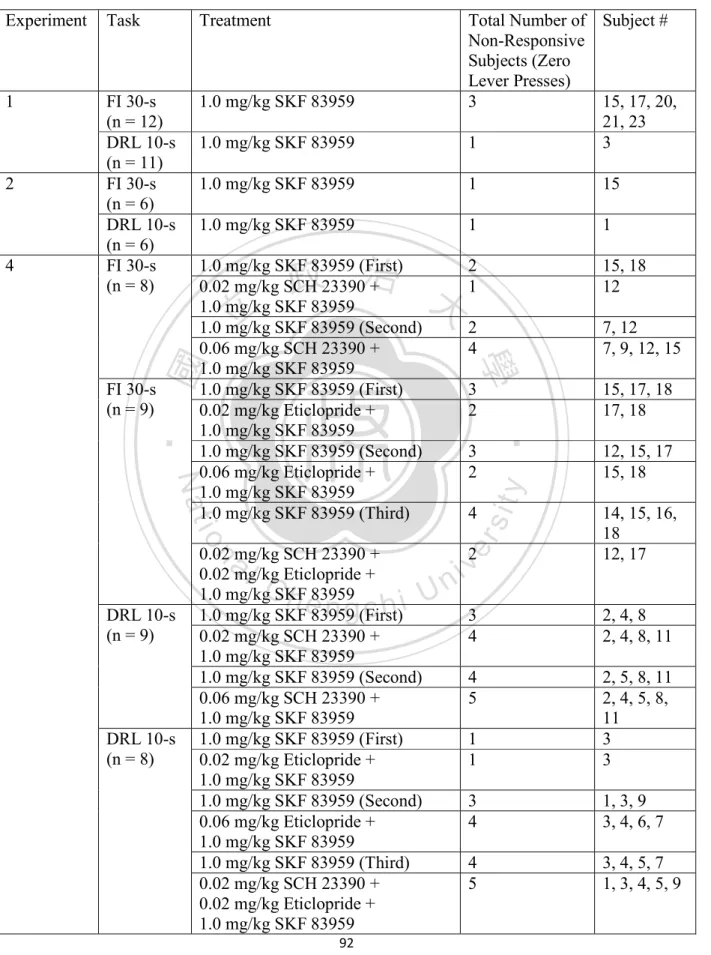
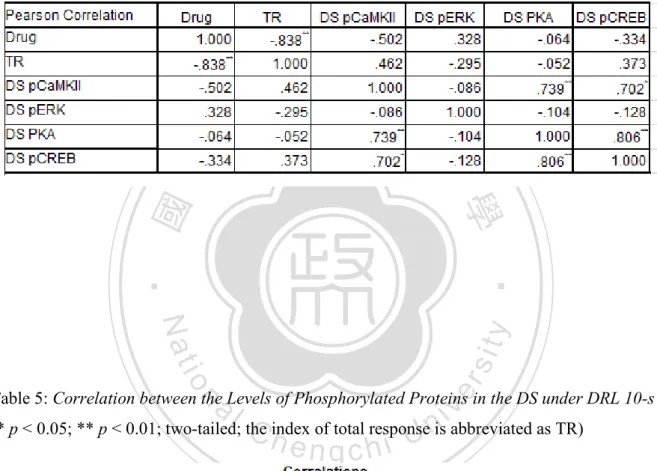
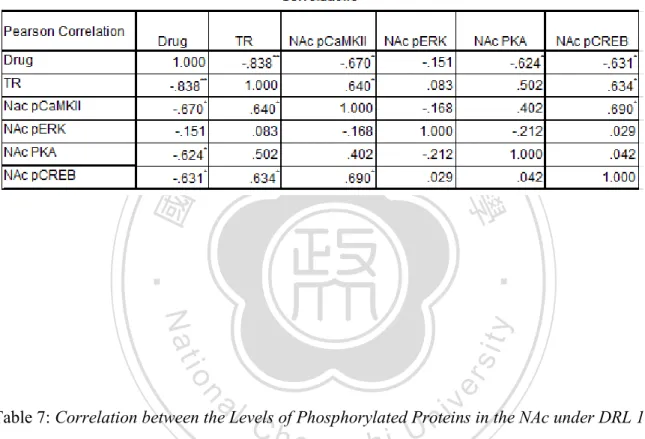
(7) List of Tables Table 1: An outline of non-responsive rats after SKF 83959 administration ............................... 92 Table 2: Correlation between levels of phosphorylated proteins in the PFC under FI 30-s ......... 93 Table 3: Correlation between levels of phosphorylated proteins in the PFC under DRL 10-s .... 93 Table 4: Correlation between levels of phosphorylated proteins in the DS under FI 30-s ........... 94 Table 5: Correlation between levels of phosphorylated proteins in the DS under DRL 10-s ...... 94 Table 6: Correlation between levels of phosphorylated proteins in the NAc under FI 30-s ........ 95 Table 7: Correlation between levels of phosphorylated proteins in the NAc under DRL 10-s .... 95. 政 治 大. Table 8: Correlation between levels of phosphorylated proteins in the hippocampus under FI 30-s ....................................................................................................................................................... 96. 立. ‧. ‧ 國. 學. Table 9: Correlation between levels of phosphorylated proteins in the hippocampus under DRL 10-s ................................................................................................................................................ 96. n. er. io. sit. y. Nat. al. Ch. engchi. VI. i n U. v.
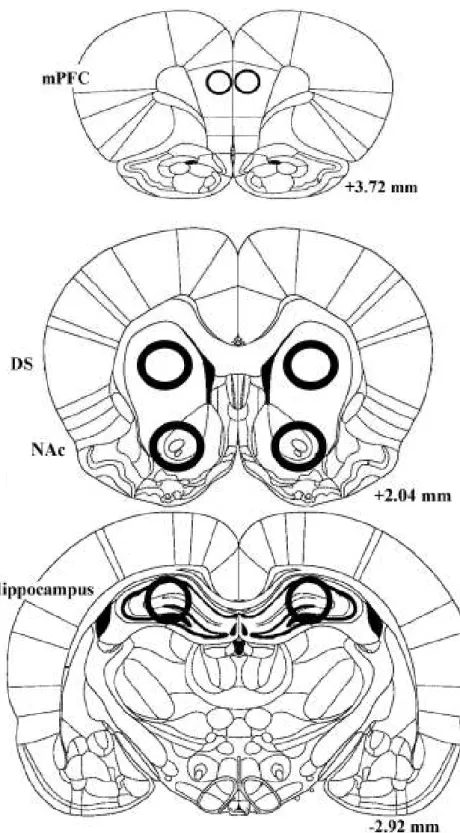
(8) List of Figures Figure 1. Schematic diagrams of the brain regions of tissue collection ....................................... 97 Figure 2. The dose effects of SKF 83959 on the IRT curve of FI 30-s performance ................. 98 Figure 3. The dose effects of SKF 83959 on FI 30-s response-based indexes and PRP ............. 99 Figure 4. The dose effects of SKF 83959 on the IRT curve of DRL 10-s performance............. 100 Figure 5. The dose effects of SKF 83959 on DRL 10-s response-based indexes……….….…..101 Figure 6. The dose effects of SKF 83959 on DRL 10-s peak rate, peak time, MRE………..…102 Figure 7. The effects of 1.0 mg/kg SKF 83959 on the IRT curve of FI 30-s performance. ....... 103. 政 治 大. Figure 8. The effects of 1.0 mg/kg SKF 83959 on FI 30-s response-based indexes and PRP ... 104. 立. Figure 9. The effects of 1.0 mg/kg SKF 83959 on the IRT curve of DRL 10-s performance.... 105. ‧ 國. 學. Figure 10. The effects 1.0 mg/kg SKF 83959 on DRL 10-s response-based indexes….............106 Figure 11. The effects of 1.0 mg/kg SKF 83959 on DRL 10-s peak rate, peak time, MRE. ..... 107. ‧. Figure 12. CaMKII expression in the specified brain regions after behavioral testing…….…..108. sit. y. Nat. Figure 13. Total ERK expression in the specified brain regions after behavioral testing……...109. io. er. Figure 14. ERK1 expression in the specified brain regions after behavioral testing………...…110 Figure 15. ERK2 expression in the specified brain regions after behavioral testing………...…111. al. n. v i n Figure 16. PKA expression in the specified behavioral testing………….....112 C h brain regions after engchi U Figure 17. CREB expression in the specified brain regions after behavioral testing………..…113 Figure 18. The dose effects of SKF 83959 on locomotor activity………………………...……114 Figure 19. The FI 30-s IRT curve after SCH 23390 pretreatment……………………………...115 Figure 20. The FI 30-s indexes after SCH 23390 pretreatment………………………………...116 Figure 21. The DRL 10-s IRT curve after SCH 23390 pretreatment…………………………..117 Figure 22. The DRL 10-s response-based indexes after SCH 23390 pretreatment…………….118 Figure 23. The DRL 10-s peak rate, peak time, and MRE after SCH 23390 pretreatment ........ 119 Figure 24. The FI 30-s IRT curve after eticlopride pretreatment………………………………120 VII.
(9) Figure 25. The FI 30-s indexes after eticlopride pretreatment ................................................... 121 Figure 26. The DRL 10-s IRT curve after eticlopride pretreatment……………………………122 Figure 27. The DRL 10-s response-based indexes after eticlopride pretreatment……………...123 Figure 28. The DRL 10-s peak rate, peak time, and MRE after eticlopride pretreatment……...124 Figure 29. The FI 30-s IRT curve after the co-administration of SCH 23390 and eticlopride…125 Figure 30. The FI 30-s indexes after the co-administration of SCH 23390 and eticlopride……126 Figure 31. The DRL 10-s IRT curve after the co-administration of SCH 23390 and eticlopride………………………………………………………………………………………………127. 政 治 大. Figure 32. The DRL 10-s reponse-based indexes after the co-administration of SCH 23390 and eticlopride………………………………………………………………………………………………128. 立. ‧. ‧ 國. 學. Figure 33. The DRL 10-s peak rate, peak time, and MRE after the co-administration of SCH 23390 and eticlopride……………………...……………………………………………………129. n. er. io. sit. y. Nat. al. Ch. engchi. VIII. i n U. v.
(10) Introduction The Behavioral Functions of DA: A Historical Overview The theory of operant conditioning by Skinner (1938), which described the reinforcement of actions with positive outcomes, has led to the search for reward-related regions in the brain, to better understand the mechanisms of operant learning. Olds and Milner (1954) implanted electrodes into rat brains and tested their operant responses for electrical stimulation, in a paradigm later known as the intracranial self-stimulation (ICSS). It was found that electrodes. 政 治 大. implanted in the septal area, located in the central forebrain beneath the corpus callosum,. 立. corresponded with increased responding for electrical stimulation during the acquisition phase. ‧ 國. 學. (Olds & Milner, 1954). Olds and Olds (1963) aimed to more precisely identify the brain loci involved in positive and negative reinforcement by implanting electrodes into different brain. ‧. regions across animal subjects and testing their responses on self-stimulation tasks; it was found. y. Nat. sit. that the implantation of electrodes in the medial forebrain bundle (MFB) was highly correlated. n. al. er. io. with increased responding for electrical stimulation, in other words, with positive reinforcement.. i n U. v. As ascending norepinephrine (NE) axons were observed within the MFB, and the. Ch. engchi. pharmacological manipulations of NE transmission with monoamine oxidase inhibitors (eg. damphetamine) and inhibitors (eg. reserpine) increased or decreased animal response rates on ICSS respectively, the catecholamine hypothesis, which suggested that NE neurons in the MFB are related to the reinforcing effects of the ICSS, was proposed to explain the correlation between NE transmission and ICSS performance (Fibiger, 1978). During the popularity of the catecholamine hypothesis, DA was considered as a precursor molecule in NE synthesis (Wise, 2008). In their studies, Carlsson, Lindqvist, and Magnusson (1957) administered 3,4dihydroxyphenylalanine (DOPA; a precursor in DA and NE syntheses) to mice, and found the 1.
(11) drug to counteract the motor deficits induced by reserpine. Carlsson, Lindqvist, Magnusson, and Waldeck (1958) further reported that such observations were related to the levels of DA instead of NE in the brain. Together, these findings led to the proposal that DA is not a mere reaction intermediate during NE synthesis; it may function as a neurotransmitter on its own (Carlsson et al., 1958). The monoamine oxidase inhibitors and catecholamine inhibitors mentioned earlier do not only affect NE transmission, but the DA system as well. Hence subsequent studies investigated. 治 政 大abilities to perform on selfnoradrenergic transmission was found to debilitate the general 立. the effects of NE- or DA-selective antagonism on operant responding. The antagonism of brain. administration tasks in animals rather than affecting their reward-mediated responses (Wise,. ‧ 國. 學. 2008). On the contrary, the blockade of DA transmission with pimozide decreased the operant. ‧. responding for food without affecting the general abilities to perform on the task (Wise, Spindler, De Wit, & Gerberg, 1978). As pimozide appeared to block the rewarding effects of food, the. y. Nat. er. io. sit. anhedonia hypothesis was proposed to suggest an important role of brain DA in the processing of pleasure and reward-mediated motivation (Wise et al., 1978; Wise, 1982). Since then, research. n. al. Ch. i n U. v. on reward-mediated brain regions has begun to focus on DA transmission in the brain. In recent. engchi. years, studies on the DA system have identified its involvement in functions beyond the processing of hedonic stimuli; the DA system is now believed to possess functional roles in a variety of activities, spanning from motivated behaviors, motor movement, effort-related decision making, to habit formation, reward prediction, and interval timing (Salamone & Correa, 2012; Schultz, Dayan, & Montague, 1997; Meck, Penney, & Pouthas, 2008). As selective agonists and antagonists for DA receptors (DARs) become available, it is possible to experimentally activate specific dopaminergic pathways and discover their impacts on animal. 2.
(12) behaviors. The emergence of studies on the various aspects of DA functions offers insights to the underlying mechanisms of motor movement, operant conditioning, cognitive processing, as well as the future understanding of pathological problems like drug addiction, Parkinson’s disease (PD), and schizophrenia. Particularly, the present proposed study will focus on the role of DA in reward-mediated operant behaviors.. DA Receptors and Signaling Pathways. 政 治 大 The transmission of DA is mediated via DARs on the cellular level. Research on the DA 立. ‧ 國. 學. system has identified 5 subtypes of G-protein-coupled DARs. The D1 and D5 receptors were categorized into the D1-like family based on their biochemical properties to stimulate the. ‧. downstream enzyme adenylyl cyclase (AC) via stimulatory α subunits of G proteins (Gαs/olf); the. sit. y. Nat. D2, D3, and D4 receptors were categorized as D2-like based on their properties to inhibit AC via. io. er. inhibitory α subunits of G proteins (Gαi/0) (Kebabian & Calne, 1979; Missale, Nash, Robinson,. al. v i n C h in the brain (Missale distinct, and they are differentially distributed e n g c h i U et al., 1998). n. Jaber, & Caron, 1998). The two classes of DARs are pharmacologically and biochemically. The differential modulation over AC by D1- and D2-like receptor activation leads to different downstream signaling cascades. The activation of AC via D1-like receptors in the striatum leads to the formation of the second messenger cyclic adenosine 3’5’monophosphate (cAMP), which continues on to activate protein kinase A (PKA), a cAMP-dependent protein kinase (Stoof & Kebabian, 1981). PKA will phosphorylate the DA- and cAMP-regulated phosphoprotein, 32 kDa (DARPP-32), which will go on to inhibit protein phosphatase-1 and indirectly increase the activation of extracellular signal regulated kinase (ERK), which is a. 3.
(13) protein in the glutamatergic N-methyl-D-aspartate receptor (NMDAR) signaling cascade (Greengard, Allen, & Nairn, 1999; Valjent et al., 2005). The signaling of PKA and ERK will lead to the phosphorylation of the cAMP response element binding protein (CREB), which is a transcription factor in the nucleus that serves as an integration site for signals from DAR and NMDAR activation and regulates gene expression for synaptic plasticity (Gonzalez & Montminy, 1989; Zanassi et al., 2001; Neve, Seamans, & Trantham-Davidson, 2004; Nishi, Kuroiwa, & Shuto, 2011).. 治 政 大of downstream proteins in the activation is expected to result in the signaling and modulation 立 In contrast to D1-like receptor activation, the inhibition of AC by D2-like receptor. opposite direction (Neve et al., 2004). Nishi, Snyder, and Greengard (1997) have showed that. ‧ 國. 學. quinpirole treatment (a D2 receptor (D2R)-selective agonist) reduced the phosphorylation of. ‧. DARPP-32 in mice striatal slices via a calcium-dependent mechanism. Furthermore, Yan, Feng, Fienberg, and Greengard (1999) have demonstrated that quinpirole treatment induced the. y. Nat. er. io. sit. phosphorylation of mitogen-activated protein kinases (MAPK)/ERK and CREB in brain slices via a protein kinase C (PKC)-dependent pathway involving intracellular calcium, calcium and. n. al. Ch. i n U. v. calmodulin-dependent protein kinase (CaMK), and DARPP-32. These findings on the signal. engchi. transduction of D1- and D2-like receptors help to determine the relationship between the receptor functions and their respective signaling pathways. As both classes of the DARs were found to exert modulation over MAPK/ERK and CREB phosphorylation, this signaling mechanism may be related to the interaction between D1 receptors (D1Rs) and D2Rs (Yan et al., 1999). Functionally, the cAMP/PKA signaling pathway in the striatum has been suggested to be involved in the neural plasticity underlying reward-related learning; it may also be related to long-term potentiation in the hippocampus (Beninger & Miller, 1998). On the other hand, ERK. 4.
(14) signaling is implicated in the synaptic plasticity related to learning and memory; the levels of striatal ERK activation are under the coordinated influence of the NMDARs and D1Rs (Zanassi et al., 2001; Shiflett & Balleine, 2011). In addition to the traditionally-identified AC pathway, Undie and Friedman (1990) discovered that the respective treatment of DA and the non-selective DA agonist apomorphine to rat brain slices could induce the accumulation of inositol phosphates in the amygdala, hippocampus, striatum, and frontal cortex; the drug-induced increases in inositol phosphate. 治 政 大suggesting that the underlying selective for the other receptors (such as sulpiride and atropine), 立. levels was blocked by the D1R-selective antagonist SCH 23390 and not by the antagonists. mechanism may be specifically related to D1Rs. In examination of how much of this DA-induced. ‧ 國. 學. phosphoinositide (PI) hydrolysis was related to the AC-coupled D1Rs, Undie, Weinstock, Sarau,. ‧. and Friedman (1994) tested the abilities of several D1R agonists of benzazepine derivative in stimulating PI hydrolysis and AC activation using rat striatal slices. The abilities of the tested. y. Nat. er. io. sit. compounds in activating PI hydrolysis and AC were not correlated, suggesting that the observed PI hydrolysis may be mediated by a D1-like receptor that is pharmacologically unique from the. n. al. Ch. i n U. v. AC-coupled D1-like receptors (Undie et al., 2004). These findings have inspired subsequent. engchi. studies to explore possible differences in the receptor composition, intracellular signaling, and behavioral functions between the AC- and PI-linked D1-like receptors.. DA Transmission Pathways DA transmission in the brain is mainly classified into the nigrostriatal pathway, the mesolimbic pathway, and the mesocortical pathway according to the routes of neural projection.. 5.
(15) First, the nigrostriatal pathway transmits DA from the substantia nigra (SNr) to the striatum, which has been shown by studies on PD to be implicated in functions of motor control and time perception. Post-mortem studies on patients with PD have reported a common degeneration of nigrostriatal DA in their brains (Kish, Shannak, & Hornykiewicz, 1988); animal lesion studies that introduced the neurotoxin 6-hydroxydopamine (6-OHDA) into the SNr or MFB have also demonstrated impaired motor movement in the animals (Deumens, Blokland, & Prickaerts, 2002). In relation to timing abilities, PD patients tended to underestimate the duration of time. 治 政 大of the DA precursor levodopaimpairment in time perception was reduced with the treatment 立 intervals on time estimation tasks and overestimate them on time reproduction tasks; this. carbidopa (Pastor, Artieda, Jahanshahi, & Obeso, 1992).. ‧ 國. 學. Second, the mesolimbic pathway transmits DA from the ventral tegmental area (VTA) to. ‧. the nucleus accumbens (NAc) located in the ventral striatum and the limbic nuclei; whereas the mesocortical pathway projects DA from the VTA to the prefrontal cortex (PFC). These two. y. Nat. er. io. sit. pathways may be altogether referred to as the mesocorticolimbic DA pathway in discussion of the DA projection from the VTA to the limbic nuclei and PFC. The mesocorticolimbic system is. n. al. Ch. i n U. v. comprised of interconnected neurotransmission; in addition to receiving dopaminergic input. engchi. from the VTA, the NAc also receives glutamatergic input from the medial PFC (mPFC) and limbic nuclei (Pierce & Kumaresan, 2006). In response to these neural inputs, medium spiny neurons (MSNs) of the NAc project gamma amino-butyric acid (GABA) output to the ventral pallidum, VTA, and SNr, which in turn send GABAergic signals to the medial dorsal thalamus, from where glutamatergic signals to the mPFC closes this circuit (Pierce & Kumaresan, 2006). A closer examination of the mesocorticolimbic DA pathway indicated that the striatal output can be distinguished as the direct striatonigral and the indirect striatopallidal pathways. 6.
(16) (Parent, Bouchard, & Smith, 1984). While both pathways transmit GABAergic signals, the striatonigral pathway projects from the striatal MSNs to the medial globus pallidus, then on to the SNr, and the striatopallidal pathway projects from the striatum to synaptic relays in the lateral globus pallidus, on to the subthalamic nucleus, finally to the SNr (Albin, Young, & Penney, 1989). The striatonigral pathway is characterized by neurons selectively expressing substance P and dynorphin, whereas the striatopallidal pathway is characterized by neurons expressing enkephalin (Bertran-Gonzalez, Herve, Girault, & Valjent, 2010). Early in situ hybridization and. 治 政 大 with the probes for D R, half of the striatal neurons, the majority of which were co-labeled 立. retrograde labeling experiments demonstrated that the striatonigral neurons constituted nearly 1. dynorphin, and substance P, rarely with the probes for D2R or enkephalin (Gerfen et al., 1990).. ‧ 國. 學. The remaining neurons were co-labeled with D2R and enkephalin probes, hence the distribution. ‧. of D1R and D2R in the striatum was proposed to be segregated despite the presence of a fraction. y. Nat. of striatal neurons that may express both D1R and D2R (Gerfen et al., 1990). The differential. er. io. sit. expression of substance P/dynorphin and enkephalin in striatal neurons has thereafter been utilized as protein markers to identify between striatonigral and striatopallidal neurons (Bertran-. al. n. Gonzalez et al., 2010).. Ch. engchi. i n U. v. The Discovery from D1R and D2R Co-localization to D1-D2 Receptor Heteromers While immunocytochemistry experiments using electron microscopy have reported that most of the D1R- and D2R-expressing MSNs in the dorsal striatum (DS) appeared segregated in distinct populations, co-localized D1R and D2R immunoreactivity was also observed in small numbers (Hersch et al., 1995). From in situ hybridization experiments, Lester, Fink, Aronin, and. 7.
(17) DiFiglia (1993) reported a co-localization of D1R and D2R mRNAs in about 26% of the striatal cells. Using fluoroprobes labeling, Wong, Shetreat, Clarke, and Rayport (1999) also observed a co-localization of D1- and D2-like receptors in over 20% of the cultured cells from the rat striatum and NAc. In light of the supportive evidence for D1-D2 receptor co-localization in the striatum, the possibility of a synergistic link between D1Rs and D2Rs has been proposed. Keefe and Gerfen (1995) found the co-administration of D1R- and D2R-selective agonists (SKF 38393 and. 治 政 大Subsequently, Lee et al. (2004) compared to the administration of either receptor agonist alone. 立. quinpirole) to enhance the levels of immediate early gene expression in DA-depleted rat striatum. observed a novel phospholipase C (PLC)-mediated calcium increase in cultured cells co-. ‧ 國. 學. expressing D1R and D2R upon the concurrent administration of D1R- and D2R-selective agonists. ‧. (SKF 81297 and quinpirole). The rise in intracellular calcium release was not observed when the. y. Nat. agonists were co-administered to cells expressing only D1Rs or D2Rs, neither when either agonist. er. io. sit. was administered alone to D1R-D2R co-expressing cells (Lee et al., 2004). Moreover, the dual agonist-induced calcium release could be blocked by either the D1R-selective antagonist SCH. n. al. Ch. i n U. v. 23390 or the D2R-selective antagonist eticlopride (Lee et al., 2004).. engchi. The findings on D1-D2 receptor co-localization and synergy led to the hypothesis that the D1R and D2R may interact to form a heteromeric signaling unit, whose activation possibly leads to the activation of PLC-mediated calcium release rather than the classic D1R-coupled AC pathway (Lee et al., 2004). This hypothesis of D1-D2 receptor heteromerization has received support from studies that used the confocal fluorescence resonance energy transfer (FRET) technique, which demonstrated co-localized D1R and D2R being in close proximity (within 5-7 nm of relative distance) in rat NAc in 10-20% of the NAc MSNs (Hasbi et al., 2009; Perreault et. 8.
(18) al., 2010; Hasbi et al., 2011; Chun, Free, Doyle, Huang, Rankin & Sibley, 2013). The existence of D1-D2 receptor heteromers was more directly demonstrated by co-immunoprecipitation experiments on D1-D2 receptor co-expressing cultured cells and cells from rat striatal tissues, which showed D1R and D2R to be parts of a common protein complex (Lee et al., 2004). Recently, O’Dowd, Ji, Nguyen, and George (2012) have identified the interaction of glutamic acid residues in the carboxyl tail of D1Rs with arginine residues in the intracellular loop 3 of D2Rs as a critical step in the formation of D1-D2 receptor heteromers, adding more details to the. 治 政 大therapeutic targets for drug Currently, D -D heteromers are perceived as potential 立. understanding of D1-D2 receptor heteromerization. 1. 2. addiction and schizophrenia, as they were found to exhibit enhanced activity and sensitivity after. ‧ 國. 學. repeated amphetamine treatment in rat striatum as well as in patients with schizophrenia. ‧. (Perreault, Hasbi, O’Dowd, & George, 2013; Perreault et al., 2010). Thus the availability of. y. Nat. compounds that act directly on the D1-D2 heteromers would be useful for possible experimental. n. al. er. io. sit. manipulations in future studies.. Ch. engchi. i n U. v. SKF 83959 as a Proposed Agonist for D1-D2 Receptor Heteromers SKF 83959 (R,S-3-methyl-6-chloro-7,8-dihydroxy-1-[3’-methylphenyl]-2,3,4,5tetrahydro-1H-benzazepine) is a compound of benzazepine derivative previously recognized as a low-efficacy D1R partial agonist (Neumeyer, Kula, Bergman, & Baldessarini, 2003). It was found to induce animal behaviors characteristic of D1R activation, such as grooming, vertical jaw movements, incisor chattering, and contralateral rotations in rats treated with unilateral 6-OHDA lesions (Downes & Waddington, 1993; Deveney & Waddington, 1995; Panchalingam & Undie,. 9.
(19) 2001; Makihara et al., 2007; Gnanalingham, Hunter, Jenner, & Marsden, 1995). However, it exhibited antagonist-like effects biochemically on DARs in failing to stimulate AC activity and actually inhibiting DA-induced AC activity (Arnt, Hyttel, & Sanchez, 1992). These observations led to the speculation that the atypical mechanism of SKF 83959 may be related to a DA signaling pathway that is independent of AC. In support of this idea, Panchalingam and Undie (2001) reported an induction of PLC-mediated PI hydrolysis in rat and monkey brain slices upon SKF 83959 treatments; hence SKF 83959 was suggested to act on the previously reported PI-. 治 政 大 83959 was applied to D -D Rashid et al. (2007) detected a modest calcium signal when SKF 立. linked D1-like receptor in the brain (Undie & Friedman, 1990; Undie et al., 2004). Subsequently, 1. 2. receptor co-expressing human embryonic kidney (HEK) cells in vitro; the calcium signal was. ‧ 國. 學. actually enhanced when SKF 83959 was co-administered with the D2R agonist quinpirole.. ‧. Radioactively-labeled guanosine triphosphate (GTPγS) assays showed the administration of. sit. y. Nat. SKF 83959 to significantly increase the incorporation of labeled GTPs into Gq/11 proteins,. io. er. suggesting that the PI-linked D1-like receptor may transmit neural signals via Gq proteins in contrast to Gs or Gi (Rashid et al., 2007). It is worth noting that the activation of Gq/11 by SKF. al. n. v i n C h of either SCH 23390 83959 was prevented by the administration or raclopride, which further engchi U. demonstrated the critical roles of concurrent D1R and D2R stimulation in the activation of the Gq/11-linked signaling cascade (Rashid et al., 2007). In summary of the above findings, the PIlinked D1-like signaling unit was hypothesized to be composed of the heterodimerized D1-D2 receptors, which can be activated by the novel compound SKF 83959, leading to the stimulation of Gq/11 proteins, PLC-mediated PI hydrolysis, and finally the release of intracellular calcium followed by the activation of the calcium/calmodulin-dependent protein kinase II (CaMKII) (George & O’Dowd, 2007; Hasbi, O’Dowd, & George, 2010).. 10.
(20) However, a contradictory finding has been reported by Chun et al. (2013), in which they found SKF 83959 to antagonize calcium mobilization in D1-D2 receptor co-expressing HEK cells and possess high binding affinities to several G-protein coupled receptors other than the D1R or D2R. Chun et al. raised doubts regarding the biochemical mechanism and receptor specificity of SKF 83959 as a D1-D2 receptor heteromer-specific agonist, and suggested that the D1-D2 receptor-mediated calcium signaling may not be entirely heteromer-specific and may involve other downstream signaling pathways.. 治 政 careful interpretation, the compound has also been utilized in大 behavioral studies to compare the 立 While the biochemical mechanism of SKF 83959 still requires further investigation and. behavior functions of AC- and PI-coupled D1-like receptor activation. For example, SKF 83959. ‧ 國. 學. was reported to differ from the AC-coupled SKF 83822 in stimulating the onset of maternal. ‧. behaviors in rats (Stolzenberg, Zhang, Luskin, Ranker, Bress, & Numan, 2010). In addition, Cools, Lubbers, Van Oosten, and Andringa (2002) found the intra-accumbal injection of SKF. y. Nat. er. io. sit. 83959 at high doses to antagonize the locomotor activity induced by SKF 81297, an AC-coupled D1R agonist. According to Cools et al. (2002), the administration of SKF 83959 alone did not. n. al. Ch. i n U. v. have significant effects on locomotor activity. Using the rat model of 6-OHDA lesion, Zhang,. engchi. Ma, Wang, Chen, and Zhen (2007) reported that SKF 83959 induced contralateral rotations with less severe symptoms of dyskinesia. Moreover, the chronic administration of SKF 83959 was found to decrease the dyskinesia induced by L-DOPA (Zhang et al., 2007). In monkeys, high doses of SKF 83959 have been reported to induce catalepsy (Platt, Rowlett, & Spealman, 2000). Apart from the studies using reflexive behavioral models as described above, several studies related to cocaine addiction have tested the effects of SKF 83959 on tasks involving conditioned behaviors in monkeys. Platt et al. (2000) found that the pretreatment of SKF 83959. 11.
(21) dose-dependently attenuated the cocaine-induced increases in response rates on 10-minutes of fixed-interval (FI) shock termination tasks, and produced rightward shifts in the cocaine doseresponse functions on fixed-ratio (FR) cocaine discrimination tasks in squirrel monkeys. The administration of SKF 83959 alone produced cocaine-reversible decreases in the overall response rates at doses that did not induce catalepsy or muscle rigidity (Platt et al., 2000). Similarly, using second-order FR 10 or FR 30 schedules within 10-minute intervals, Platt, Rowlett, and Spealman (2001) found the pretreatment of SKF 83959 to produce dose-related. 治 政 大 using the same second-order respectively. In another study on reinstatement to cocaine priming 立 decreases in the response rates on self-administration tasks for cocaine and food pellets. schedules, Khroyan, Barrett-Larimore, Rowlett, and Spealman (2000) demonstrated that SKF. ‧ 國. 學. 83959 on its own did not induce reinstatement of responding at the doses tested, while the co-. ‧. treatment of SKF 83959 with an effective priming dose of cocaine showed dose-dependent inhibition on the reinstatement of responding in the monkeys. From these findings, SKF 83959. y. Nat. er. io. sit. appeared to exhibit antagonist-like effects on cocaine-induced behaviors and demonstrated a general suppressive effect on the overall response rates of operant performance.. n. al. Ch. i n U. v. As the functional role and cellular mechanism of D1-D2 receptor heteromers remain. engchi. unclear, an investigation on the effects of SKF 83959 on operant behaviors is an intriguing attempt to determine the possible behavioral functions of D1-D2 receptor heteromer activation.. DA and Schedule-Controlled Behaviors DA transmission in the brain has been known to possess a critical role in reward- and motivation-mediated behaviors. The design and introduction of the operant chamber by Skinner. 12.
(22) (1938), which consisted of a box with at least one lever inside for collecting behavioral responses from subject animals, has been a classic animal model to test the relationship between DA transmission and incentive-motivated behaviors (Salamone & Correa, 2012). In this model, animals are usually deprived of a primary reinforcer such as food or water, in order for them to become motivated to learn and behave according to the schedules of reinforcement to obtain the reward. Different schedules of reinforcement provide means to study the animals’ choice behaviors under different external contexts (Morgan, 2010). Specific schedules of reinforcement. 治 政 大 related behavioral function (Sanger & Blackman, 1989). 立. are characterized by distinct behavioral components that can be used to probe the particular DA-. Two common types of reinforcement schedules are the continuous reinforcement, under. ‧ 國. 學. which a response is reinforced every time it occurs, and the partial or intermittent reinforcement,. ‧. when only parts of the responses are reinforced (Chance, 1979). Schedules of reinforcement vary on the basis of frequency ratios or time intervals; ratio schedules are based on the numbers of. y. Nat. er. io. sit. responses emitted by the subject, while interval schedules are based on responding after the passage of a specified time interval (Schoenfeld, Cumming, & Hearst, 1956; Zeiler, 1977). For. n. al. Ch. i n U. v. example, every nth response is reinforced regularly under FR schedules, whereas the animal’s. engchi. first response after a designated time interval of n seconds is regularly reinforced under FI schedules (Chance, 1979). Both ratio and interval schedules serve to test the acquisition and performance of operant behaviors in animals, yet the use of interval schedules additionally allows the measurement of their capabilities in timing performance. Interval timing refers to the ability to discriminate and produce temporal durations within the seconds to minutes range (Drew, Fairhurst, Malapani, Horvitz, & Balsam, 2003). Interval timing is also a DA-related function as striatal MSNs have. 13.
(23) been proposed to help in the discrimination of temporal durations by detecting the patterns of cortical oscillations from neuronal firing in the network of the nigrostriatal and mesocorticolimbic pathways (Meck et al., 2008). Finally, the option of differential schedules of reinforcement allows researchers to selectively reinforce some desirable aspects of animal behavior in their experimental design. The differential reinforcement of low rate (DRL) that decreases the animals’ response rates while maintaining a previously-acquired operant behavior is one of the representative examples of the differential schedules of reinforcement (Chance, 1979).. 治 政 大is in the generation of response A main difference between ratio and interval schedules 立. rates; ratio schedules tend to maintain higher response rates than interval schedules under the. ‧ 國. 學. same rates of reinforcement, and interval schedules are able to maintain responding when the. ‧. rates of reinforcement are very low, a characteristic that is not observed in ratio schedules (Baum, 1993). The present proposed study will use interval schedules as behavioral tasks to investigate. y. Nat. er. io. sit. the DA-related functions of reward-motivated operant behaviors and timing performance, namely the FI 30-s and DRL 10-s schedules of reinforcement. The former presents a reward. n. al. Ch. i n U. v. every 30 seconds after the trial starts, hence the animal eventually learns to start responding as. engchi. the end of the 30 seconds lapse approaches; the plots of the inter-response time (IRT) of FI schedules are usually scallop-shaped, reflecting an increase in responding as the designated interval approaches. In contrast, the DRL 10-s schedule presents a reward only when the animal makes a response that has been at least 10 seconds from its previous response; any responses within the 10 seconds lapse will reset the countdown. In other words, the animal will learn to withhold from responding during the set interval under DRL schedules in order to gain the reward. Hence the DRL 10-s schedule maintains a lower rate of responding in the animals than. 14.
(24) the FI 30-s schedule (Chang, Liao, Lan, & Shen, 2000; Chiang, 2006). Despite the difference in the generation of response rates, the FI 30-s and DRL 10-s schedules of reinforcement are characterized by different behavioral components and supposed to involve different neural mechanisms. Although both tasks require normal motor abilities for making responses, the FI 30-s schedule probes the animals’ abilities in timing over fixed intervals, while the DRL 10-s schedule has more stringent rules for timing over resettable periods. Moreover, the DRL 10-s schedule is characterized by a unique component of behavioral. 政 治 大. inhibition since the animals need to learn to withhold from responding for a certain length of time.. 立. ‧ 國. 學. The Operant Behavioral Pharmacology of DA-Related Drugs. ‧. sit. y. Nat. Previous pharmacological studies have utilized various direct or indirect DA agonists and. io. er. antagonists to test the effects of drug-induced enhancement or blockade in DA transmission on operant behaviors. The respective administration of selective D1R and D2R antagonists (SCH. al. n. v i n Creduce 23390 and YM 09151-2) was found to of lever-pressing for food on FR U h e nthegrates i h c. schedules in rats (Rusk & Cooper, 1994). It was generally concluded that the blockade of D1R and D2R decreased the hedonic effects of reward on animals’ responding (Beninger & Miller, 1998). Similarly, the respective administration of D1R- and D2R-selective agonists (SKF 38393 and N-0437) was also found to decrease the operant responding for food under FR schedules of reinforcement (Rusk & Cooper, 1989; Rusk & Cooper, 1988). In respective to interval-based operant tasks, the administration of DA agonists to rats trained under the peak interval procedure was found to speed up their internal clock, leading to. 15.
(25) an underestimation of time intervals; alternatively, the administration of DA antagonists slowed down their internal clock, resulting in overestimated time intervals (Drew et al., 2003). Multiple studies have found the administration of amphetamine-related compounds to shift the peak time of the performance on various DRL schedules, including DRL-15s, DRL-16s, and DRL-36s, to the left (Sanger, Key, & Blackman, 1974; Segal, 1962; Sabol, Richards, Layton, & Seiden, 1995). Similarly, the respective administration of D1R- and D2R-selective agonists (SKF 38393 and quinpirole) to rats was observed to result in a leftward shift on the response distributions of peak. 治 政 interval after cue presentation, and peak trials, when cues are大 presented for an extended length of 立 interval procedures, which consisted of learning trials, when rewards are given at a fixed time. time in the absence of rewards (Frederick & Allen, 1996). On FI tasks, amphetamine-related. ‧ 國. 學. compounds were found to induce an increase in response rates early on during the intervals,. ‧. which was also suggestive of a faster internal clock (Maricq, Roberts, & Church, 1981). In. y. Nat. contrast, the administration of D1R- and D2R-selective antagonists (SCH 23390 and eticlopride). er. io. sit. induced a rightward trend in the response distributions of peak interval procedures that did not reach statistical significance (Frederick & Allen, 1996). Also using the peak interval procedure,. n. al. Ch. i n U. v. Drew et al. (2003) compared the effects of the D1R antagonist SCH 23390 and D2R antagonist. engchi. haloperidol on two time intervals (12 and 36 sec) of operant performance; the administration of SCH 23390 and haloperidol both decreased the response rates, while haloperidol impaired the timing performance (Drew et al., 2003). In summary of the above findings, the respective activation and blockade of D1Rs and D2Rs both resulted in decreased response rates for food under FR schedules. In contrast, the pharmacological activation of D1Rs and D2Rs induced underestimated time intervals and a faster internal clock, whereas the blockade of D1Rs and D2Rs resulted in overestimated time intervals. 16.
(26) and a slower internal clock. Although the D1Rs and D2Rs exert opposite modulation over AC, they do not necessarily affect behaviors oppositely to each other. Hence it may be useful to examine the potential relationship between changes in behavioral performance and cellular signaling in order to better understand the functions of the DARs.. Objective of the Present Study. 政 治 大 D R monomers, the present study aimed to investigate the behavioral and biochemical changes 立. As previous research mainly examined the functions and neural mechanisms of D1R and. 2. ‧ 國. 學. that may be induced by the activation of the recently-identified D1-D2 receptor heteromers. The D1-D2 receptor heteromers were pharmacologically activated via the administration of SKF. ‧. 83959, a selective D1-D2 receptor heteromer agonist (Neumeyer et al., 2003).. sit. y. Nat. In Experiment 1, the operant performance on the FI 30-s and DRL 10-s schedules of. io. er. reinforcement were measured in rats under different doses of SKF 83959 administration. As stated earlier, these two interval-based tasks consisted of different behavioral components, whose. al. n. v i n C h drug-inducedUchanges in animal behavior. SKF contrast may provide insights to the particular engchi. 83959 was intraperitoneally (i.p.) injected at the doses of 0.01, 0.1, and 1.0 mg/kg relative to the vehicle treatment (0 mg/kg) in a within-subjects design to determine the dose effects of SKF 83959 on the respective schedules of operant behaviors. The doses chosen were in reference to Zhang et al. (2005). In Experiment 2, the brain tissues were collected from the rats after behavioral testing under the effective dose of SKF 83959 to determine if there were any drug-induced brain regionspecific changes in the patterns of selected protein expression using Western Blot. To do so, the. 17.
(27) rats were divided into two groups and received an i.p. injection of either the vehicle or the most effective dose of SKF 83959 from Experiment 1 in a between-subjects design. The behavioral test session was immediately followed by animal sacrifice and tissue collection. Brain tissues were collected from regions of the mesocorticolimbic system, including the PFC, DS, NAc, and dorsal hippocampus. The levels of expression of the following four proteins in their phosphorylated versus native forms in each of the four selected brain regions were analyzed: 1) CaMKIIα, a calcium-dependent protein kinase downstream of D1-D2 receptor heteromer. 政 治 大 kinase downstream of the classic AC-coupled D R pathway, to be used as an indicator of typical 立 activation and NMDAR activation, to be used as an indicator of calcium modulation; 2) PKA, a 1. ‧ 國. 學. D1R activation; 3) ERK, a kinase downstream of NMDAR activation that is also indirectly modulated by D1R activation; 4) CREB, a transcription factor in the cell nucleus that regulates. ‧. gene expression. Any task-specific or brain region-specific changes in the levels of the proteins. sit. y. Nat. selected above were investigated. Also, the statistical correlation between the behavioral indexes. io. D2 heteromer activation via SKF 83959 administration.. al. er. and protein levels was conducted to determine the possible trends that underlie the effects of D1-. n. v i n C hof naive rats was used In Experiment 3, a separate batch to test the dose effects of SKF engchi U. 83959 on locomotor activity using the open field test. The purpose of the locomotor activity test was to investigate whether SKF 83959 might have any debilitating impacts on the rats’ motor functions that may potentially impair their performance on the operant tasks. In Experiment 4, experiments of pharmacological antagonism tested the effects of SCH 23390 and eticlopride pretreatments in antagonizing the effects of SKF 83959 on operant behaviors. SCH 23390 and eticlopride are selective DAR antagonists binding on D1Rs and D2Rs respectively.. 18.
(28) Together, the present study investigated whether SKF 83959, a putative agonist for D1-D2 heteromers, would affect the behavioural performance on two interval-related schedules of reinforcement. Also, the biochemical effects of SKF 83959 on the levels of selected protein expression in the mesocorticolimbic DA system were investigated. Based on a previous behavioral study on SKF 83959, the compound exhibited DA antagonist-like effects on the operant response rates in monkeys (Platt et al., 2000). Hence the administration of SKF 83959 in the present study was hypothesized to decrease the response rates in rats on the operant tasks. It. 治 政 previous knowledge about the biochemical properties of SKF大 83959 (Rashid et al., 2007), it was 立. was of interest to find out whether the drug would affect temporal processes in the rats. Based on. hypothesized that the drug would not significantly change the levels of PKA. SKF 83959 was. ‧ 國. 學. hypothesized to exhibit greater impacts on the levels of pCaMKII or CaMKII in the regions. ‧. encompassing the mesocorticolimbic DA system. It was also of interest to investigate the drug effects on the levels of ERK and CREB in the brain DA terminals, as no such report has been. n. al. er. io. sit. y. Nat. made at this point.. Ch. engchi. 19. i n U. v.
(29) Methods Subjects The experiments were conducted using male Wistar rats purchased from Biolasco Taiwan Co., Ltd as subjects. They were housed individually in stainless steel hanging cages in a temperature- (23◦C) and humidity- (60%) controlled colony room under 12 hours of light and dark cycles (lights on from 8am to 8pm). Initially the rats had unrestricted access to water and food until their body weight reached 280-300 g; after which access to water was restricted to 5. 政 治 大. minutes per day and behavioral training commenced. During the subsequent training and. 立. experimental stages, access to water was maintained at 5 minutes per day, made available to the. ‧ 國. 學. rats at 30 minutes after the completion of the behavioral procedures. Access to food remained ad. ‧. io. sit. y. Nat. Apparatus. n. al. er. libitum.. Ch. i n U. v. Operant behaviors. Operant responses were measured using six equal operant chambers. engchi. (MED Associated, St. Albans, VT, USA), in the dimensions of 20 x 25 x 30 cm. The floors of the chambers were composed of 18 stainless steel rods with the diameter of 5 mm, each spaced 11 mm apart; the panels on the sides were made of aluminum, and the front and back walls of clear Plexiglas. The ceiling of the chambers consisted of clear Plexiglas. Each chamber contained a lever, a water-receiving dish connected to an external liquid dispenser, and a house light. The lever was a 4.5 x 2 cm piece of metal positioned 7 cm above the floor and 2 cm from the left corner of the front panel. The liquid dispenser was installed on the outside of each chamber, set to deliver 0.03 to 0.05 ml of tap water per reinforcement to the receiving dish 20.
(30) located at the center of the front panel 2 cm above the floor for each reinforced response. The light source in each chamber was provided by a light bulb of 2.5 cm diameter, located 12 cm above the floor and 2 cm from the left corner of the front panel, right above the lever. The chambers were enclosed in 60 x 40 x 58 cm plywood boxes with fans installed on the top righthand corners for ventilation and controlled background noise. The operant chambers were connected to computers for automatic control via the printer port and PCI interface card for input and output. The computer program for setting up the. 治 政 大task, the computer program Windows 98; Microsoft) (Liao & Cheng, 2007). During the FI 立. different reinforcement schedules and data recording was written in Visual Basic (version 6.0 for. collected three behavioral indexes, which included the inter-response time (IRT; the lapse of. ‧ 國. 學. time between responses), the number of total responses, the number of reinforced responses, and. ‧. the post-reinforcement pause (PRP; the lapse of time between a reinforced response and its next response). During the DRL task, the program similarly collected the IRT, the number of total. y. Nat. er. io. sit. responses, the number of reinforced and non-reinforced responses; additionally, the program recorded the number of burst responses (responses within 3 seconds from previous responding),. n. al. Ch. i n U. v. and calculated the peak time (the mean value of IRTs that fall within the 4 consecutive 1-sec bins. engchi. containing maximum response frequencies), the peak rate (the sum of response frequencies during the 4 bins divided by 4), and the modified response efficiency (MRE; the ratio between the number of reinforced responses and the number of total responses minus the number of burst responses) (Liao & Cheng, 2007). Locomotor activity. The open field locomotor activity test was conducted in an assembly of four identical black acrylic boxes (45 x 45 x 36 cm each). A charge coupled device (CCD) camera was installed above the center of the assembly at 52 cm from the ground. The. 21.
(31) camera was connected to a desktop computer, which recorded the distance travelled by the rats as well as their average and maximum speed.. Training of FI and DRL Behaviors The water-deprived rats started with the magazine training, during which they learned to associate water with the metal receiving dish. Then the rats continued on to three daily 30-minute. 政 治 大 associate lever presses with the appearance of the reinforcer (water) on the receiving dish. All of 立 sessions of shaping on the FR 1 schedule of reinforcement, from which the rats learned to. the rats were expected to make at least 65 lever presses in a 30 min training session to meet the. ‧ 國. 學. criterion of this stage. The rats were then randomly assigned into separate groups for training. ‧. under either the FI 30-s (n = 12) or DRL 10-s (n = 12) schedules of reinforcement.. sit. y. Nat. Rats in the FI group began daily training under one-hour sessions of the FI 10-s schedule. io. er. for 10 days until their average number of total responses reached 900. Then the rats continued with daily 30-minute training sessions under FI 30-s for 25 days until the average number of total. al. n. v i n Ch responses reached 600 before the commencement of pharmacological test sessions. In contrast, engchi U rats in the DRL group were trained by hourly sessions of the DRL 10-s schedule for 10 days followed by daily 30-minute training sessions for 25 days; the testing of drug-induced. performance began when their average number of total responses reached 200. Over the course of the operant training, stable baseline performance on both tasks were indicated by the criterion of less than 10% of variation on average response rates for three consecutive days.. Drugs 22.
(32) SKF 83959 hydrobromide (Tocris Bioscience) was dissolved in the vehicle containing 10% ethanol. SCH 23390 hydrochloride (RBI, Research Biochemicals Inc.) and eticlopride hydrochloride (Tocris Bioscience) were dissolved in 0.9% physiological saline. All drugs were i.p. injected at the volume of 1 ml/kg of body weight.. Western Blot. 政 治 大 lysis buffer for homogenization, centrifugation, and dilution with lysis buffer. The lysates were 立. The collected brain tissues were added with protease inhibitor, phosphatase inhibitor, and. tested for protein concentrations, prepared into samples of 1 μg/μl, and boiled for 10 minutes. ‧ 國. 學. at 90◦C before loading for Western blot. The sample lysates (20μl) were loaded onto 10% SDS. ‧. polyacrylamide gel for separation by electrophoresis, and they were later transferred onto a. sit. y. Nat. polyvinylidene fluoride (PVDF) membrane via electroblotting. The sections of the membrane. io. er. that contained the proteins of interest (according to molecular weight) were cut out into a strip. al. v i n C h membrane was then autoshaking under room temperature. The e n g c h i U incubated with the primary n. and immersed in 2% BSA (bovine serum albumin) for 1 hour of blocking at 40 rpm of. antibody (diluted 1:2000) on an autoshaker overnight at 4 ◦C. The primary antibodies included the rabbit anti-pCaMKII (Upstate), mouse anti-CaMKII (Millipore), rabbit anti-pERK1/2 (Cell Signaling), rabbit anti-ERK1/2 (Cell Signaling), rabbit anti-PKA (Upstate), mouse anti-pCREB (Millipore), and rabbit anti-CREB (Millipore). On the following day, the membrane was washed three times (for 5, 10, 10 minutes) in 0.1% TBST (Tris-buffered saline, 0.1% Tween 20) and incubated with the secondary antibody (1:4000, either anti-mouse or anti-rabbit IgG conjugated to horseradish peroxidase (HRP)) on a 40 rpm autoshaker for 1 hour under room temperature.. 23.
(33) Then the membrane was washed again for three times in 0.1% TBST before detection by chemiluminescent reaction with Immobilon Western chemilminescent HRP substrate (Millipore). The primary antibody for actin was used as an internal control and followed the same protocols. The protein bands were quantified by ImageJ (version 1.47, National Institutes of Health, USA).. Procedures. 政 治 大 When the rats were trained to stable baseline performance on the operant tasks, they were tested 立. Experiment 1: The dose effects of SKF 83959 on FI 30-s and DRL 10-s performance.. for the dose effects of SKF 83959. The dose treatments of SKF 83959 (0, 0.01, 0.1, and 1.0. ‧ 國. 學. mg/kg) were given via i.p. administration in a within-subjects design for the DRL 10-s (n = 11). ‧. and FI 30-s (n = 12) groups on four consecutive days. The drug administration was given 30. sit. y. Nat. minutes prior to the behavioral session. After injection, the rat was kept in a holding cage,. io. er. located in the behavior test room, for 30 minutes until it was placed into the operant chamber. A 30-minutes session under the FI 30-s or DRL 10-s schedule of reinforcement was conducted in. n. al. this dose-effect test.. Ch. engchi. i n U. v. Experiment 2: The effects of SKF 83959 on operant performance and levels of selected protein expression in mesocorticolimbic DA terminals. The rats from Experiment 1 underwent six days of retraining on their respective schedules of reinforcement until the performance returned to stable baseline. The rats within the FI or DRL group were then divided to receive either the vehicle (n = 6 in the FI 30-s groups and n = 5 in the DRL 10-s group) or the most effective dose of SKF 83959 (n = 6 for each schedule) in a between-subjects design. The rats in the FI 30-s group that made less than 3 lever presses in Experiment 1 (#15, 17, 20, 21, 23). 24.
(34) were randomly distributed to the vehicle group (#17, 21, 23) and the drug treatment group (#15, 20). The rat in the DRL 10-s group that made no lever presses in Experiment 1 (#3) was placed into the vehicle group in Experiment 2. Using the aforementioned subject assignment, there were no significant differences in the baseline numbers of total responses between groups before drug injections in Experiment 2. On the FI 30-s schedule, the vehicle group made 562 ± 98 responses, and the experimental group made 565 ± 53 responses (F = 0.0008, p > 0.05). On the DRL 10-s schedule, the vehicle group made 195 ± 9 responses while the experimental group made 193 ± 21. 政 治 大 same as the description in Experiment 立 1. Following the drug-treated behavior test, the rats were. responses (F = 0.0099, p > 0.05). The procedures of the drug test on operant behaviors were the. ‧ 國. 學. decapitated. Tissues were collected from the specified brain regions including the PFC, DS, NAc, and dorsal hippocampus (Figure 1). The collected brain tissues were treated with liquid nitrogen,. ‧. and stored in a -80◦C freezer until preparation for Western blot analysis.. y. Nat. io. sit. Experiment 3: The effects of SKF 83959 on locomotor activity. A separate batch of. n. al. er. naive rats (n = 18) was divided into 3 groups for either 0, 0.5, or 1.0 mg/kg of i.p. SKF 83959. i n U. v. injection (n = 6 each). After the respective drug injection, rats were placed in their holding cages. Ch. engchi. for 30 min before being placed into the open field boxes for a 30-min locomotor activity test. The average travel speed and total travel distance were recorded in 5-min intervals. Experiment 4: The effects of SCH 23390 and eticlopride pretreatment on SKF 83959-induced operant performance. The respective effects of pretreating SCH 23390, eticlopride, and their combination on SKF 83959-induced operant behaviors were tested by giving two (or three) i.p. drug injections during each session. The first injection was given 60 min before the behavioral test, and the second injection was given 30 min before the behavioral. 25.
(35) test. On the first test day, all rats were subjected to control treatments (the double injection of saline-vehicle). After a day of retraining, the rats went on to receive the drug treatments of saline-SKF 83959, DAR antagonist-saline, and DAR antagonist-SKF 83959 in the orders of a Latin square on test sessions each spaced apart by two days of retrain. SKF 83959 was injected at 1.0 mg/kg, which was the effective dose from Experiment 1 and Experiment 2. SCH 23390 and eticlopride were both tested at 0.02 and 0.06 mg/kg, based on previous studies from this lab (Cheng & Liao, 2007) and others (Fowler & Liou, 1998; Schindler & Carmona, 2002).. 政 治 大 locomotor activity test, and they立 were divided into two groups to receive operant training under SCH 23390. The rats from Experiment 3 started water deprivation two weeks after the. ‧ 國. 學. either the FI 30-s (n = 9) or DRL 10-s (n = 9) schedules of reinforcement, in the procedures as described earlier. After reaching the stable baseline, the rats were subjected to the tests with 0.02. ‧. mg/kg SCH 23390 pretreatments. The rats were retrained for three days, after which they were. sit. y. Nat. subjected to testing with 0.06 mg/kg SCH 23390 pretreatments.. n. al. er. io. Eticlopride. A separate batch of naïve rats (n = 18) were divided into the FI 30-s and. i n U. v. DRL 10-s groups for operant training as described above. When they reached stable baseline. Ch. engchi. performance, the rats were subjected to operant testing with 0.02 mg/kg eticlopride pretreatments. After three days of retraining, they were again subjected to testing with 0.06 mg/kg eticlopride pretreatments. The combination of SCH 23390 and eticlopride. After the eticlopride treatments, the rats were retrained for three days before being subjected to the combined pretreatments of 0.02 mg/kg SCH 23390 and 0.02 mg/kg eticlopride. The control treatments on the first day were a. 26.
(36) triple injection of saline-saline-vehicle. On experimental sessions, the double injections of SCH 23390 and eticlopride were given in alternating orders.. Statistical Analysis The data are presented in mean ± the standard error of the mean (SEM), and analyzed with ANOVA using Statistica (version 7.1, StatSoft). Post-hoc comparisons were conducted. 政 治 大 the exceptions of cases where the ANOVA yielded significant results and the Tukey’s HSD did 立. using Tukey’s honestly significant difference (HSD) test with a significance level of p < 0.05. In. ‧ 國. 學. not, the least significant difference (LSD) test was used to account for possible over-corrections by Tukey’s HSD when sample sizes were not large enough. Bivariate correlations between. ‧. behavioral measures and protein results were conducted with SPSS (version 16.0, SPSS Inc.).. sit. y. Nat. It should be noted that some rats appeared to completely cease operant responding upon. io. er. the administration of 1.0 mg/kg SKF 83959 (see Table 1). Cases of such low (number of total. al. responses less than five) or zero responses were still included in the analysis, because the. n. v i n C h may introduce bias exclusion of these cases in notable numbers e n g c h i U to the results. Hence all of the actual numbers were used in the analysis of response-based indexes, while the group mean was substituted in these cases of missing data on the other indexes: PRP on the FI task, peak time, peak rate, and MRE on the DRL task. For some of the rats whose levels of protein expression obviously deviated from the group mean, their protein data were considered as outliers and removed from the analysis of that particular protein. This did not exclude them from the behavioral analysis as the deviation was attributed to possible human error during the process of Western blotting.. 27.
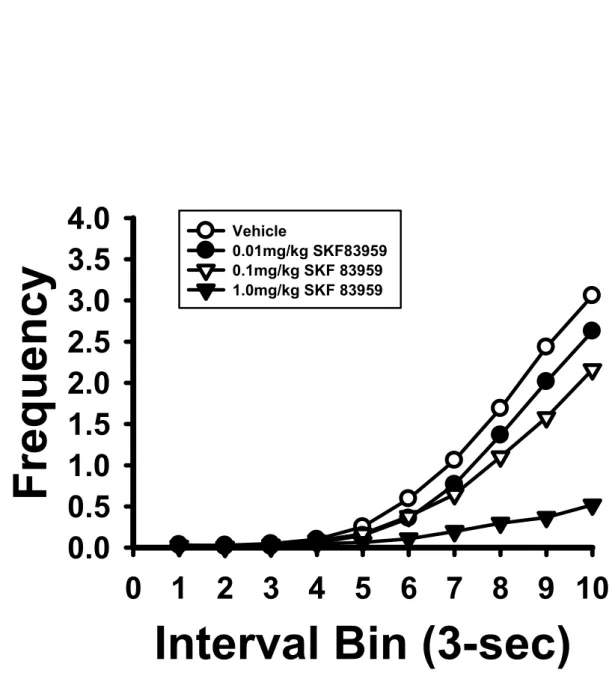
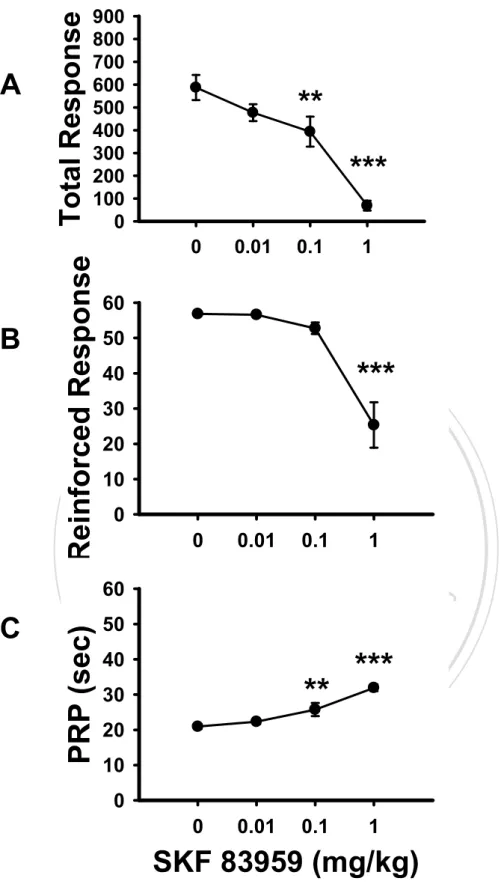
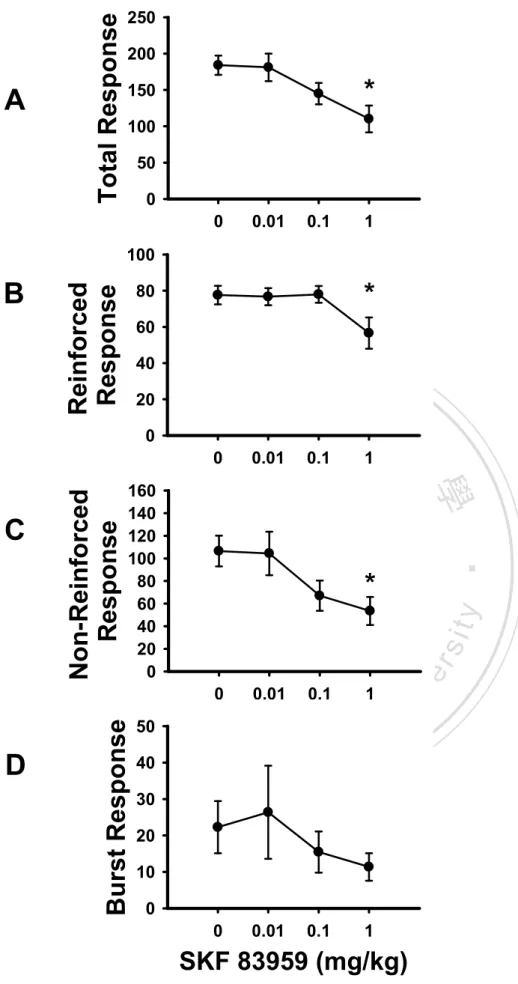
(37) Results. Experiment 1: The Dose Effects of SKF 83959 on FI 30-s and DRL 10-s Performance The FI 30-s task. As illustrated in Figure 2, the rats trained under the FI 30-s schedule exhibited a dose-dependent decrease in response rates on the IRT curve of a 30-s interval as they received higher doses of SKF 83959 injections. One-way ANOVA showed that drug dose had significant effects on the numbers of total responses, F(3, 33) = 32.95, p < 0.001, reinforced. 政 治 大 indicated by Tukey’s HSD post-hoc test, the rats emitted significantly fewer numbers of total 立. responses, F(3, 33) = 22.51, p < 0.001, and the duration of PRP, F(3, 33) = 26.11, p < 0.001. As. ‧ 國. 學. responses when they were injected with SKF 83959 at the medium dose (0.1 mg/kg; p < 0.01) and the high dose (1.0 mg/kg; p < 0.001) (Figure 3A). The rats also showed significantly fewer. ‧. numbers of reinforced responses upon high dose SKF 83959 administration (1.0 mg/kg; p <. sit. y. Nat. 0.001; Figure 3B); their durations of PRP were significantly increased after injections with. io. er. medium and high dose SKF 83959, p < 0.01 and p < 0.001 respectively (Figure 3C). As noted in. al. Table 1, three of the twelve rats tested had completely ceased to respond on the lever during the. n. v i n C hadministrations, while test session upon 1.0 mg/kg SKF 83959 e n g c h i U an additional number of two rats emitted equal to or less than 2 lever presses during the session after high dose injection. The DRL 10-s task. The rats trained under the DRL 10-s schedule similarly showed a dose-dependent decrease in response rates on the IRT curve as they were injected with higher doses of SKF 83959, while the peak time remained relatively in place (Figure 4). One-way ANOVA found the drug dose to have significant effects on three of the four response-based indexes, including the numbers of total responses, F(3, 30) = 4.53, p < 0.01, reinforced responses, F(3, 30) = 3.49, p < 0.05, and non-reinforced responses, F(3, 30) = 3.49, p < 0.05. Tukey’s HSD. 28.
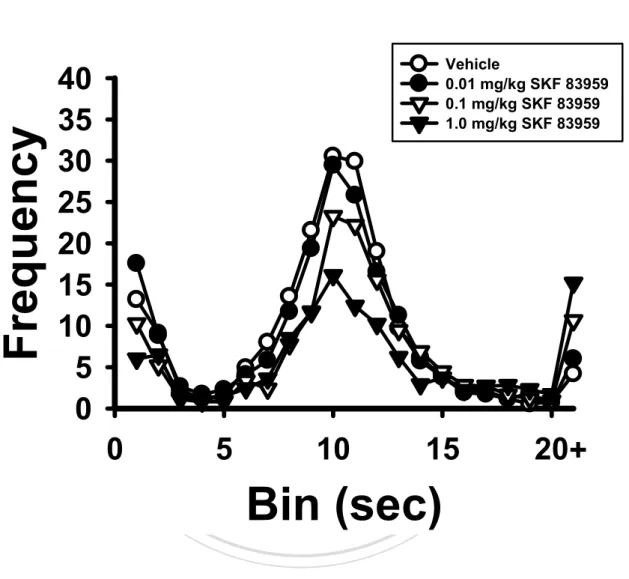
(38) post-hoc test showed that the total responses were significantly decreased when the rats were treated with high dose SKF 83959, p < 0.05 (Figure 5A). Post-hoc by LSD also found significantly reduced numbers of reinforced responses, p < 0.05 (Figure 5B) and non-reinforced responses, p < 0.05 (Figure 5C) under high dose SKF 83959 administration. The number of burst responses was not affected by SKF 83959, p > 0.05 (Figure 5D). In terms of the other DRL indexes, the peak rate was significantly affected by the doses of SKF 83959, F(3, 30) = 6.58, p < 0.01. Tukey’s HSD showed significantly decreased peak rate under high dose SKF 83959. 治 政 大test, one of the eleven rats tested administrations (Figure 6B & 6C respectively). During the DRL 立. injection, p < 0.01 (Figure 6A). The peak time and MRE were unaffected by SKF 83959. emitted no responses upon high dose SKF 83959 injection.. ‧ 國. 學. Experiment 2: The Effects of SKF 83959 on Operant Performance and Levels of Selected. Nat. y. ‧. Protein Expression in Mesocorticolimbic DA Terminals. sit. Operant performance. The behavioral effects of 1.0 mg/kg SKF 83959 replicated the. er. io. findings of Experiment 1 to a great extent.. al. n. v i n C h7, the administration FI 30-s task. As shown in Figure of 1.0 mg/kg SKF 83959 reduced engchi U. the response rates on the IRT curve. In comparison to the vehicle group, the SKF 83959-treated rats showed significantly decreased numbers of total responses, F(1, 10) = 23.49, p < 0.001 (Figure 8A) and reinforced responses, F(1, 10) = 16.12, p < 0.01 (Figure 8B). The drug-treated rats exhibited significantly increased durations of PRP, F(1, 10) = 18.53, p < 0.01 (Figure 8C). One of the six rats in the experimental group emitted no responses upon SKF 83959 injection. DRL 10-s task. Similarly, the administration of 1.0 mg/kg SKF 83959 also reduced the frequency of lever presses on the IRT curve (Figure 9). The drug-treated rats showed. 29.
數據

+7
相關文件
6 《中論·觀因緣品》,《佛藏要籍選刊》第 9 冊,上海古籍出版社 1994 年版,第 1
The first row shows the eyespot with white inner ring, black middle ring, and yellow outer ring in Bicyclus anynana.. The second row provides the eyespot with black inner ring
You are given the wavelength and total energy of a light pulse and asked to find the number of photons it
1, the Educational Research Establishment (ERE) of the Education Department undertook four research projects on the medium of instruction in secondary schools, three of which
好了既然 Z[x] 中的 ideal 不一定是 principle ideal 那麼我們就不能學 Proposition 7.2.11 的方法得到 Z[x] 中的 irreducible element 就是 prime element 了..
volume suppressed mass: (TeV) 2 /M P ∼ 10 −4 eV → mm range can be experimentally tested for any number of extra dimensions - Light U(1) gauge bosons: no derivative couplings. =>
incapable to extract any quantities from QCD, nor to tackle the most interesting physics, namely, the spontaneously chiral symmetry breaking and the color confinement..
• Formation of massive primordial stars as origin of objects in the early universe. • Supernova explosions might be visible to the most
