Research Article
Selection of alkaline phosphatase-positive induced pluripotent
stem cells from human amniotic fluid-derived cells by
feeder-free system
☆
Huai-En Lu
a,b, Ming-Song Tsai
c,d, Yao-Chen Yang
a, Chen-Ching Yuan
a, Tzu-Hao Wang
e,
Xi-Zhang Lin
f, Ching-Ping Tseng
b,⁎
, Shiaw-Min Hwang
a,⁎⁎
aBioresource Collection and Research Center, Food Industry Research and Development Institute, Hsinchu 30062, Taiwan, ROC bDepartment of Biological Science and Technology, National Chiao Tung University, Hsinchu 30068, Taiwan, ROC
cPrenatal Diagnosis Center, Cathay General Hospital, Taipei 10630, Taiwan, ROC d
School of Medicine, Fu Jen Catholic University, Taipei 24205, Taiwan, ROC
eDepartment of Obstetrics and Gynecology, Chang Gung Memorial Hospital, Lin-Kou Medical Center, Taoyuan 33305, Taiwan, ROC fDepartment of Internal Medicine, College of Medicine, National Cheng Kung University, Tainan 70403, Taiwan, ROC
A R T I C L E I N F O R M A T I O N A B S T R A C T Article Chronology:
Received 16 February 2011
Revised version received 14 May 2011 Accepted 17 May 2011
Available online 26 May 2011
Generation of induced pluripotent stem (iPS) cells from somatic cells has been successfully achieved by ectopic expression of four transcription factors, Oct4, Sox2, Klf4 and c-Myc, also known as the Yamanaka factors. In practice, initial iPS colonies are picked based on their embryonic stem (ES) cell-like morphology, but often may go on to fail subsequent assays, such as the alkaline phosphate (AP) assay. In this study, we co-expressed through lenti-viral delivery the Yamanaka factors in amniotic fluid-derived (AF) cells. ES-like colonies were picked onto a traditional feeder layer and a high percentage AF-iPS with partial to no AP activity was found. Interestingly, we obtained an overwhelming majority of fully stained AP positive (AP+) AF-iPS colonies when colonies were first seeded on a feeder-free culture system, and then transferred to a feeder layer for expansion. Furthermore, colonies with no AP activity were not detected. This screening step decreased the variation seen between morphology and AP assay. We observed the AF-iPS colonies grown on the feeder layer with 28% AP+ colonies, 45% AP partially positive (AP+/−) colonies and 27% AP negative (AP−) colonies, while colonies screened by the feeder-free system were 84% AP+ colonies, 16% AP+/− colonies and no AP− colonies. The feeder-free screened AP+ AF-iPS colonies were also positive for pluripotent markers, OCT4, SOX2, NANOG, TRA-1-60, TRA-1-81, SSEA-3 and SSEA-4 as well as having differentiation abilities into three germ layers in vitro and in vivo. In this study, we report a simplistic, one-step method for selection of AP+ AF-iPS cells via feeder-free screening.
© 2011 Elsevier Inc. All rights reserved. Keywords:
Amniotic fluid
Induced pluripotent stem cells Feeder free
Alkaline phosphatase
☆ The authors indicate no potential conflicts of interest. ⁎ Corresponding author. Fax: +886 3 5729288. ⁎⁎ Corresponding author. Fax: +886 3 5224171.
E-mail addresses:[email protected](C.-P. Tseng),[email protected](S.-M. Hwang). 0014-4827/$– see front matter © 2011 Elsevier Inc. All rights reserved.
doi:10.1016/j.yexcr.2011.05.017
a v a i l a b l e a t w w w . s c i e n c e d i r e c t . c o m
Introduction
Generation of human induced pluripotent stem (iPS) cells from somatic cells has been successfully achieved by ectopic expression of two combinations of specific transcription factors, Oct4, Sox2, Klf4 and c-Myc or Oct4, Sox2, Nanog and Lin28 [1,2]. By transduction of these factors into cells, it is possible to reprogram the differentiated cells into an embryonic stem (ES) cell-like pluripotent state and can differentiate into all major cell types represented by the three embryonic germ layers. More impor-tantly, this biological breakthrough has the potential to create not only new human disease models[3–5], but also allow for patient-specific therapies devoid of immune rejection and ethical concerns. There are many established methods to improve the generation of iPS cells, including transient transduction by plasmids[6], non-integrating adenoviral vector[7], three or less transcription factors combined with small molecules [8,9], polycistronic cassette containing all four factors[10,11], a piggyBac transposition system [12,13], direct delivery of reprogramming proteins [14] and synthetic modified mRNA [15]. In addition, different cell types have been reported to reprogram more readily than fibroblasts, such as neural progenitor cells[16,17], keratino-cytes[18], melanocytes[19], adipose stem cells[20]and amniotic fluid-derived (AF) cells[21,22]. Of the cell types that have been reprogrammed, AF cells pose more of an interest due to their earlier developmental stage and clonogenicity[23,24]. Zhou et al. (2009) and Galende et al. (2010) both claimed high efficiency and rapid reprogramming to produce iPS cells from human AF cells [21,22].
However, studies on iPS cells have a common, recurrent predicament in which cells that are reprogrammed exist in varied states of reprogramming quality. Fully reprogrammed iPS cells have a set of stringent assays based on similarities to ES cells consisting of global and specific ES gene promoter methylation status (demethylated Oct4, Nanog), ES-like genome wide expres-sion profile, viral silencing, germ layer differentiation, teratoma formation and the recently uncovered epigenetic status[25,26]. Yet, with the ongoing advancements in ES research, concisely defining a fully reprogrammed iPS cell has become more and more difficult, where previously set standards may no longer suffice. But regardless of the increasing requirements that a true iPS cell has to meet, there are preliminary rounds of assays that can discriminate the good from the bad. In practice, one of the first assays performed in assessing the quality of the iPS colonies is the alkaline phosphatase (AP) activity stain. Normally, iPS colonies will exhibit AP staining ranging from“negative (−)”, “partially stained (+/−)” to “positive stained (+)”. AP− iPS cells tend to have similar ES-like morphology but lack some of the pluripotent markers such as SSEA4 and TRA-1-60[20]. AP+ iPS cells have the potential to be bona fide pluripotent cells with full ES cell markers and differentiation potentials. With the amount of iPS colonies generated, only a portion will yield colonies that are potentially fully reprogrammed or nearly ES-like, but the methods for isolating these colonies tend to be labor intensive and time consuming. In our study, we have also observed the heterogeneous mixture of AP+, AP+/−, and AP− iPS colonies derived from AF cells by lentiviral delivery of the Yamanaka factors. Ten days post-transduction, colonies with ES-like morphology were picked for further growth and screening on a feeder-free system. We found
that over 80% iPS colonies screened with the feeder-free system were AP+ versus 28% on directly seeded on the feeder layer system. Also, AP− colonies were not found in the group screened with feeder-free culture. The feeder-free AP+ colonies were further characterized and positive for their pluripotent markers and differentiation abilities in vitro and in vivo. The AP+ iPS colonies positive for standard ES assays obtained in this study suggest feeder-free screening can serve as a novel, yet simplistic selection method for AP+ iPS colonies that may more likely yield fully reprogrammed iPS cells.
Materials and methods
Cell culture of amniotic fluid-derived cells
AF samples were obtained with informed patient consents and the protocols of this study were approved under the Institutional Review Board (IRB) of Cathay General Hospital, Taipei, Taiwan. The AF cells were isolated from second-trimester amniocentesis according to our previous report[27]. AF cells were cultured and passaged routinely at 80−90% confluence with alpha-modified minimum essential medium (α-MEM, Hyclone, Logan, UT) containing 20% fetal bovine serum (FBS, Hyclone), 4 ng/mL basic fibroblast growth factor (bFGF, Peprotech, London, UK), 50 U/mL penicillin and 50μg/mL streptomycin (Sigma, St. Louis, MO) at 37 °C and 5% CO2in a humidified incubator. AF cells at passage 4–6 were used for the generation of induced pluripotent stem cells.
Generation and selection of AF-iPS cell
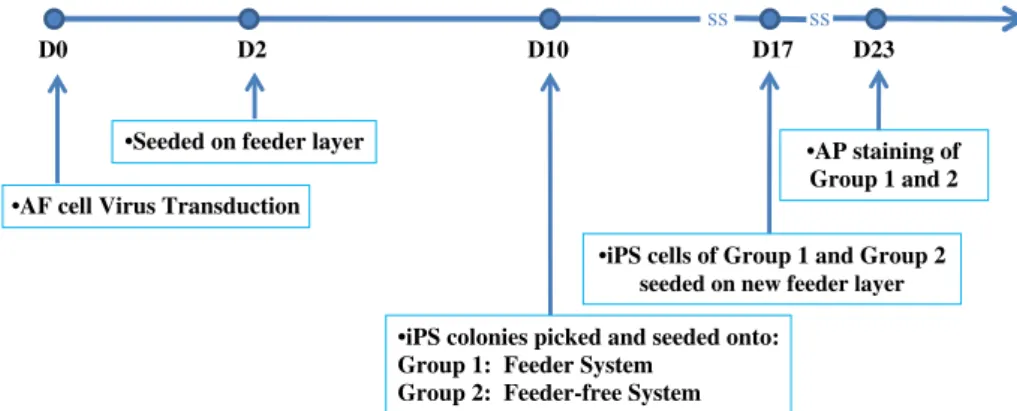
Pre-prepared lentivirus packaged with Yamanaka factors (Oct4, Sox2, Klf4, and c-Myc) was purchased from Vectorite Biomedica Inc. (VBI, Tapei, Taiwan). The virus titer used for transduction was 1 × 109 with multiplicity of infection (MOI) of 20. For viral transduction, 1 × 105AF cells were seeded on each well of 12-well culture plates. The viral mixture was then added into the culture medium with 2μg/mL polybrene (Sigma) and incubated for two days. On day 2, the cells were split on 100 mm tissue culture dishes coated with irradiated mouse embryonic fibroblasts (MEF, provided by Bioresource Collection and Research Center, Hsinchu, Taiwan) as feeder layer (1 × 106cells/ dish). Next day, the culture medium was changed to human ES cell culture medium composed of Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM/F12, Invitrogen, Carlsbad, CA) medium supplemen-ted with 20% KnockOut serum replacement (KOSR, Invitrogen), 0.1 mM nonessential amino acids (NEAA, Invitrogen), 1 mM GlutaMax-1 (Invitrogen), 0.1 mM 2-mercaptoethanol (Invitro-gen), 10 ng/mL bFGF, 50 U/mL penicillin, and 50μg/mL strepto-mycin. On day 10 post-transduction, singular ES-like colonies were picked based on morphology and cultured either on irradiated MEF feeder layer with ES culture medium (Group 1: Feeder System) or on Matrigel™ (BD Bioscience, Bedford, MA)-coated surface with mTeSR™1 (StemCell Technologies,Vancouver, Cana-da) medium (Group 2: feeder-free system). To assess the reprogrammed cells in Group 2, we designed a grading system with grades 0 to 3, according to the morphology of colonies. Grade 0 colonies were loosely attached with no ES-like morphology. Grade 1 colonies were attached with signs of regional and/or peripheral differentiation. Grade 2 colonies were ES-like but in a
loosely clumped state. Grade 3 colonies were ES-like with a visible, distinct periphery. On day 17, AF-iPS with ES-like morphology comprising of all the colonies in Group 1 and grade 2 and 3 colonies in Group 2 were reseeded on an irradiated MEF feeder layer for expansion and further characterization. The workflow chart is shown inFig. 1.
Alkaline phosphatase and immunofluorescence staining
For detecting AP activity, the AP detection kit was obtained from Millipore (Millipore, Billerica, MA) and performed following the manufacturer's instruction. For immunofluorescence staining, cells were fixed with 4% paraformaldehyde (Sigma), permeabilized with 0.05% Triton X-100 (Sigma) if necessary and blocked with 10% donkey serum. For the identification of ES markers in AF-iPS colonies, colonies were stained with primary antibodies against SOX2 (1:200, Millipore), NANOG (1:500, Abcam, Cambridge, MA), SSEA4 (1:100, Abcam), SSEA3 (1:100, Abcam), TRA-1-60 (1:200, Abcam), TRA-1-81 (1:200, Abcam), and OCT4 (1:400, Abcam). For the detection of lineage markers in differentiated AF-iPS cells in vitro, cells were stained with primary antibodies againstα-smooth muscle actin (1:100, Abcam), α-fetoprotein (1:500, Sigma), albumin (1:200, Abcam), nestin (1:100, R&D Systems, Minneap-olis, MN), troponin T-C (1:100, Santa Cruz Biotechnology, Santa Cruz., CA),β3-tubulin(1:400, Sigma) followed by staining with secondary AlexaFluor-conjugated antibodies (Invitrogen) and observed under fluorescent inverted microscope (Axio Observer, Zeiss, Germany).
Karyotype analysis
Karyotype analysis of AF-iPS cells was performed at passage 8 at the laboratory of Prenatal Diagnosis, Cathay General Hospital, Taipei, Taiwan using standard protocols for G-banding of chromo-somes. The karyotype description followed the International System for Human Cytogenetic Nomenclature (ISCN:2009, Karger Ag, Basel,http://atlasgeneticsoncology.org/ISCN09/ISCN09.html).
RT-PCR
Total RNA of each sample was extracted using TRIzol reagent (Invitrogen) and performed reverse transcription using the RevertAid™ H Minus First Strand cDNA Synthesis Kit (Fermentas,
Glen Burnie, MD). Specific cDNA was amplified by PCR using DreamTaq PCR Master Mix (Fermentas) and then, analyzed by gel electrophoresis. The primers and conditions used were reported previously[1,28].
In vitro differentiation
For the generation of embryoid bodies (EB), AF-iPS cells were treated with 200 U/mL collagenase IV (Biochrom, Cambridge, UK) in DMEM/F12 basal medium and suspended in ultra-low attach-ment 6-well microplates (Corning, Lowell. MA) with DMEM/F12 supplemented with 20% KOSR, 0.1 mM NEAA, 1 mM GlutaMax-1, 0.1 mM 2-mercaptoethanol, 50 U/mL penicillin, and 50μg/mL streptomycin for 7 days. EBs were then transferred onto Matri-gel-coated plates and cultured for another 10 days. The spontane-ous differentiating AF-iPS cells were detected for ectodermal (nestin andβ3-tubulin), mesodermal (troponin T-C and α-smooth muscle actin), and endodermal (α-fetoprotein and albumin) markers by immunofluorescence staining as mentioned above.
In vivo differentiation
The animal study conducted conformed to the Animal Protection Law (2010.01.10 Amended) published by the Council of Agricul-ture, Taiwan and approved by the Animal Care and Use Committee of Food Industry Research and Development Institute, Hsinchu, Taiwan. For teratoma formation (n = 3), 2 × 106AF-iPS cells were injected intramuscularly into the NOD-SCID mice (Biolasco, Taipei, Taiwan). Seven to eight weeks post-injection, teratomas were harvested and fixed with 10% formaldehyde (Sigma). Tissue sections were embedded in paraffin and stained with hematoxylin and eosin. The histopathological analysis was performed by Taipei Institute of Pathology (Taipei, Taiwan).
Results
Generation of AF-iPS cells
AF cells were transducted with lentiviral mixture which included OCT4, SOX2, KLF4 and cMYC genes separately at the ratio of 1:1:1:1. Two days post-infection, the AF cells were split onto feeder layer plates. We observed ES-like morphology colonies appearing on
•AF cell Virus Transduction •Seeded on feeder layer
•iPS colonies picked and seeded onto: Group 1: Feeder System
Group 2: Feeder-free System
D0 D2 D10 D17 D23
ss
ss
•AP staining of Group 1 and 2
•iPS cells of Group 1 and Group 2 seeded on new feeder layer
Fig. 1– Schematic representation of generation and selection of iPS cells from AF cells. On day 10, initial iPS cells were picked and seeded onto two groups: Group 1, feeder system; Group 2, feeder-free system, Matrigel coated surface. On day 17, selected iPS cells from both groups were plated on a feeder layer for further AP staining.
day 7 (efficiency: 0.1–0.5%) followed by manually picking of select colonies (Fig. 3A) for further growth and characterization on day 10 post-transduction. These selected colonies were seeded on either a feeder layer system with ES medium (Group 1) or on a feeder-free system, with a surface Matrigel-coated with mTeSR-1 medium (Group 2). All of the seeded cells on Group 1 grew and maintained similar morphology to an ES colony, while the colonies seeded on Group 2 grew with morphological differences (Fig. 2). Grade 0 and 1 colonies were disqualified for further characteriza-tion. The reprogrammed cells of Group 1 and grades 2 and 3 of Group 2 were reseeded onto a new feeder layer for expansion and AP activity staining (Figs. 3B and C).
Selection of fully stained AP+ AF-iPS
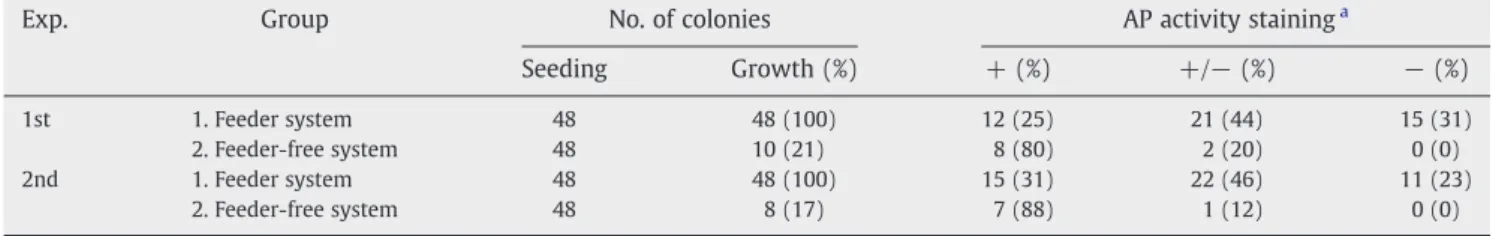
To quickly identify the quality of AF-iPS colonies, AP activity staining was performed for the reprogrammed cells from Group 1 and grades 2 and 3 of Group 2. Colonies stained for AP were classified into three groups:“AP+” colonies were AF-iPS colonies exhibiting strong and homogeneous AP activity, one of the main characteristics of pluripotency.“AP+/−” colonies were partially stained colonies exhibiting partial, heterogeneous AP activity. “AP−” colonies were colonies that exhibited very little to no AP activity (Fig. 3D). Tallying the results from the AP assessment resulted in 80–88% of “AP+” AF-iPS colonies from the feeder-free system versus 25–31% from the feeder system (Table 1). No“AP−” AF-iPS colonies were found from the feeder-free system. Compar-atively, roughly two thirds of the ES-like reprogrammed colonies from Group 1 showed“AP+/−” to “AP−” staining.
Characterization of AF-iPS cells selected from feeder-free system
After selection by the feeder-free system as grade 3 colonies, AP+ AF-iPS colonies were maintained in the traditional feeder system and further characterized for their expression of human ES cell markers by immunofluorescence staining analysis, including OCT4, SOX2, NANOG, SSEA-3, SSEA-4, TRA-1-60 and TRA-1-81. The results showed that the AP+ AF-iPS colonies from the feeder-free system were consistent with human ES characteristics (a typical example as shown inFig. 4A). Gene expressions of OCT4, SOX2 and NANOG of three AF-iPS lines from different donors were also confirmed by RT-PCR analysis (Fig. 4B). The AF-iPS cells maintained normal chromosomal karyotypes at passage 8 after selection (a typical example as shown inFig. 4C).
In vitro and in vivo differentiation of AF-iPS cells
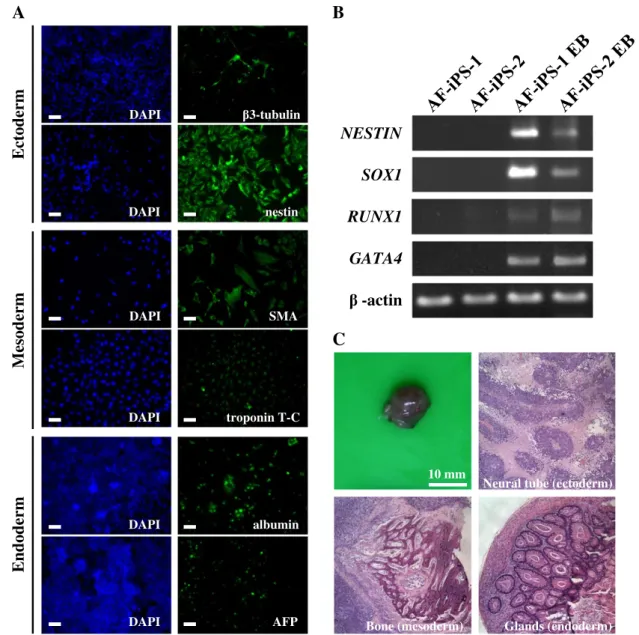
To determine the pluripotency of the AF-iPS cells selected by the feeder-free system, differentiation capabilities into three germ layers were performed in vitro and in vivo. EBs were formed in suspension on ultralow attachment plates for one week and then seeded on Matrigel-coated plates for adherence and further differentiation in vitro. The resulting cell populations were stained positively for the lineages of three germ layers, including α-smooth muscle actin (SMA) and troponin T-C (mesoderm), nestin and β3-tubulin (ectoderm), α-fetoprotein (AFP) and albumin (endoderm) (Fig. 5A). The gene expressions of the three germ layers were also confirmed by RT-PCR analysis of RUNX1 (mesoderm), GATA4 (endoderm), NESTIN and SOX1 (ectoderm) (Fig. 5B). To investigate the teratoma-forming ability of AF-iPS cells in vivo, we injected 2 × 106AF-iPS cells intramuscularly into NOD/SCID mice. Typical teratoma-like masses were observed and harvested for histopathological analysis after six to eight weeks post-transplantation. We observed the teratoma comprised of tissues from all three germ layers, such as the glands (endoderm), bone (mesoderm), and neural tube (ectoderm) (Fig. 5C).
Discussion
In this study, we reported an efficient method for the generation and selection of AP+ iPS cells from AF cells. Early studies involving amniotic fluid-derived stem cells demonstrated a specific popu-lation of CD-117 expressing cells that were able to differentiate into the three germ layers in vitro, but were not able to do so with in vivo[27]. Despite the existence of this subpopulation of pluripotent cells, it has been noted that only 1% of the collected AF cells are likely to be positive for some pluripotent markers, while the remaining heterogeneous mixture is comprised of committed progenitor cells and fully differentiated cells[29,30]. Furthermore, prolonged passaging of AF-derived stem cells demonstrated a sustained loss of pluripotent marker expressions[31]. Rather than isolating the already small subpopulation with unstable pluripo-tent markers found in amniotic fluid, generation of iPS from AF cells may serve as an alternative method to acquire pluripotent AF cells. In light of this, AF cells have been recent targets for reprogramming to a pluripotent state and have exhibited higher efficiencies than fibroblasts, suggesting AF cells may be a more suitable source for iPS cells and future patient-specific therapies
Grade 3
Grade 2
Grade 1
Grade 0
Grade 0 Loosely attached cells with no ES-like morphology
Grade 1 Attached cell colony with signs of regional and/or peripheral differentiation
Grade 2 ES-like colony in a loosely clumped state
Grade 3 ES-like colony with distinct periphery
Fig. 2– Grading system for AF-iPS on feeder free system. Grade score is in accordance with the colony morphology of AF-iPS on Matrigel coated surface.
[21,22]. There has been reported different methylation status of key senescence related genes that are over expressed in fibroblasts from older donors, which in turn has been shown to be a major barrier in iPS generation, suggesting donor age may affect the efficiency and quality of iPS generation [32,33]. In practice, amniotic fluids are routinely collected from second-trimester amniocentesis for karyotype analysis. The AF cells can be easily isolated and cultured without interrupting diagnostic procedures
[23,34], and thus have no ethical concerns other than those normally required for donation of human tissue for research including generation of iPS lines. Therefore, the potential isolation, storage, and reprogramming of prenatal AF cells may serve as a better candidate for future iPS generation.
Our initial results were consistent with previous studies dem-onstrating that reprogramming in AF cells takes place more efficiently than most other cell types[21,22]. However, we found a
AF-iPS from Group 2
AF-iPS from Group 1
AP +
AP +/-
AP -
A
B
C
D
Fig. 3– Morphology of AF-iPS colonies and AP staining. (A) Typical morphology of AF-iPS colony on day 10 post transduction of Yamanaka factors. Colonies were picked and seeded on either a feeder system (Group 1) or feeder-free system (Group 2). (B) Typical morphology of Group 1 AF-iPS colony prior to AP staining. (C) Typical morphology of Group 2 AF-iPS colony prior to AP staining. (D) AP staining of AF-iPS colonies from respective Group 1 and Group 2.
Table 1– Frequency of AF-iPS colonies in different stainings of alkaline phosphatase from feeder and feeder-free systems.
Exp. Group No. of colonies AP activity staininga
Seeding Growth (%) + (%) +/− (%) − (%)
1st 1. Feeder system 48 48 (100) 12 (25) 21 (44) 15 (31)
2. Feeder-free system 48 10 (21) 8 (80) 2 (20) 0 (0)
2nd 1. Feeder system 48 48 (100) 15 (31) 22 (46) 11 (23)
2. Feeder-free system 48 8 (17) 7 (88) 1 (12) 0 (0)
a AP: alkaline phosphatase; +: positive; +/
high percentage of AF-iPS colonies with partial to no AP activity, which was also noted by Li et al.[8]. This situation is not limited to amniotic fluid cells but all reprogramming cells. A variation exists between colonies exhibiting ES-like morphology and AP activity, and methods of decreasing this variation would benefit and speed up iPS research. There are currently no reliable methods to quickly and accurately distinguish the AP+ iPS colonies from the AP− colonies. With the advancing development of feeder-free systems, hES and iPS cells can be cultured and maintained on a Matrigel-coated surface and serum-free medium[20,35,36]. In order to approach clinical applications, the generation of iPS cells eventually needs to be on a chemically defined, feeder-free and serum-free medium. This served as the basis for our study of AF-iPS cells in feeder and feeder free environments, which also yielded a very useful and straightforward screening method. According to this selection method, the initial ES-like reprogrammed colonies were picked and cultured onto a feeder-free, Matrigel-coated surface combined with mTeSR-1 serum-free medium. A grading system was devised in accordance to colony morphology. Although the total count of grade 2 and 3 colonies was
only 20% of the feeder system, the high percentage of good quality AP+ colonies suggest iPS colonies could be successfully screened visually and rather simplistically. This selection method also could screen against AP− colonies. The selected AP+ AF-iPS cells were confirmed to express typical pluripotent markers and could differentiate into the cells of three germ layers in vitro and in vivo.
Despite current findings by using other methods to promote the efficiency of cell reprogramming, distinguishing AP+ pluripotent colonies from AP+/− colonies and AP− colonies from initial iPS cell generation still remains a challenge. In practice, ES-like colony morphology in the feeder system does not necessarily predict the level AP staining. By using a Matrigel-coated surface and mTeSR-1 medium system as a selecting platform, it is possible to reliably obtain high quality AF-iPS cells with greater efficiency without introducing exterior factors, such as reporter genes that may further compromise cell integrity. Furthermore, it has been found that during the process of iPS generation, culturing of iPS under serum-free and feeder-free culture conditions may induce further epigenetic modifications sustaining undifferentiated growth of pluripotent cells[37].
DAPI DAPI DAPI OCT4 SOX2 NANOG DAPI DAPI SSEA3 SSEA4 DAPI DAPI TRA-1-60 TRA-1-81
B
C
OCT4 SOX2 NANOG -actinA
Fig. 4– Characterization of AP+ AF-iPS colonies from a feeder-free system. (A) Immunofluorescent staining of pluripotent markers on AP+ AF-iPS colonies, including nuclear antigens for OCT4, SOX2, NANOG and cell surface antigens for SSEA-3, SSEA-4, TRA-1-60 and TRA-1-81. Scale bar, 50μm. (B) Gene expression of OCT4, SOX2 and NANOG of AF-iPS cells from different donors by RT-PCR analysis. ES cell as positive control. AF cells, unreprogrammed cells. (C) Karyotyping analysis of a typical AP+ AF-iPS cells. The cells maintained normal karyotype at passage number 8 after feeder-free selection.
Conclusion
In conclusion, we have verified that AF cells not only reprogram rapidly to a pluripotent state, but have also found that fully stained AP+ AF-iPS colonies and AP− AF-iPS can be readily selected and screened for by subculture on a Matrigel-coated surface, feeder free system. By being able to efficiently generate and select for AP+ AF-iPS cells, researchers may have more optimized methods for iPS generation, which cuts down both time and labor. Simplistic and standardized methods also
provide consistent quality and quantity of iPS cells needed for the basic study of underlying molecular processes, investigation of congenital disorders, prenatal cellular therapy and various recessive genetic disorders.
Acknowledgments
We thank Davey Leu for critically reading and commenting on the manuscript. This work was supported by a grant from the Ministry of Economic Affairs, Taiwan (99-EC-17-A-17-R7-0525).
Mesoderm
SMA DAPI troponin T-C DAPIEctoderm
3-tubulin DAPI nestin DAPIEndoderm
albumin DAPI DAPI AFPA
B
C
10 mm Bone (mesoderm)Neural tube (ectoderm)
Glands (endoderm)
RUNX1
SOX1
GATA4
-actin
NESTIN
Fig. 5– Differentiation potentials of AP+ AF-iPS colonies in vitro and in vivo. (A) Immunofluorescent staining of EB differentiation. EBs were formed on ultralow attachment plates for 7 days and then seeded on Matrigel-coated plates for 10 days in vitro. The resulting cell populations were stained for the markers of three germ layers, includingα-smooth muscle actin (SMA) and troponin T-C (mesoderm), nestin andβ3-tubulin (ectoderm), α-fetoprotein (AFP) and albumin (endoderm). Scale bar, 50 μm. (B) Gene expression of three germ layer markers for AF-iPS EB differentiated cells by RT-PCR analysis, including NESTIN, SOX1 (both for ectoderm), RUNX1 (mesoderm) and GATA4 (endoderm). Corresponding negative controls can be seen in AF-iPS-1 and AF-iPS-2. (C) Teratoma formation of AF-iPS cells into NOD/SCID mice in vivo. AF-iPS cells, 2 × 106cells were intramuscularly injected into NOD/SCID mice. Typical teratoma-like masses were harvested for histopathological analysis after six to eight weeks post-transplantation (n= 3). The tissues of three germ layers were presented. Hematoxylin–eosin (H&E) staining of teratoma sections.
R E F E R E N C E S
[1] K. Takahashi, K. Tanabe, M. Ohnuki, M. Narita, T. Ichisaka, K. Tomoda, S. Yamanaka, Induction of pluripotent stem cells from adult human fibroblasts by defined factors, Cell 131 (2007) 861–872.
[2] J. Yu, M.A. Vodyanik, K. Smuga-Otto, J. Antosiewicz-Bourget, J.L. Frane, S. Tian, J. Nie, G.A. Jonsdottir, V. Ruotti, R. Stewart, I.I. Slukvin, J.A. Thomson, Induced pluripotent stem cell lines derived from human somatic cells, Science 318 (2007) 1917–1920. [3] K. Saha, R. Jaenisch, Technical challenges in using human induced
pluripotent stem cells to model disease, Cell Stem Cell 5 (2009) 584–595.
[4] I.H. Park, N. Arora, H. Huo, N. Maherali, T. Ahfeldt, A. Shimamura, M.W. Lensch, C. Cowan, K. Hochedlinger, G.Q. Daley, Disease-specific induced pluripotent stem cells, Cell 134 (2008) 877–886. [5] X. Carvajal-Vergara, A. Sevilla, S.L. D'Souza, Y.S. Ang, C. Schaniel, D.F. Lee, L. Yang, A.D. Kaplan, E.D. Adler, R. Rozov, Y. Ge, N. Cohen, L.J. Edelmann, B. Chang, A. Waghray, J. Su, S. Pardo, K.D. Lichtenbelt, M. Tartaglia, B.D. Gelb, I.R. Lemischka, Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syndrome, Nature 465 (2010) 808–812.
[6] K. Okita, M. Nakagawa, H. Hyenjong, T. Ichisaka, S. Yamanaka, Generation of mouse induced pluripotent stem cells without viral vectors, Science 322 (2008) 949–953.
[7] M. Stadtfeld, M. Nagaya, J. Utikal, G. Weir, K. Hochedlinger, Induced pluripotent stem cells generated without viral integration, Science 322 (2008) 945–949.
[8] W. Li, W. Wei, S. Zhu, J. Zhu, Y. Shi, T. Lin, E. Hao, A. Hayek, H. Deng, S. Ding, Generation of rat and human induced pluripotent stem cells by combining genetic reprogramming and chemical inhibitors, Cell Stem Cell 4 (2009) 16–19.
[9] D. Huangfu, K. Osafune, R. Maehr, W. Guo, A. Eijkelenboom, S. Chen, W. Muhlestein, D.A. Melton, Induction of pluripotent stem cells from primary human fibroblasts with only Oct4 and Sox2, Nat. Biotechnol. 26 (2008) 1269–1275.
[10] L. Shao, W. Feng, Y. Sun, H. Bai, J. Liu, C. Currie, J. Kim, R. Gama, Z. Wang, Z. Qian, L. Liaw, W.S. Wu, Generation of iPS cells using defined factors linked via the self-cleaving 2A sequences in a single open reading frame, Cell Res. 19 (2009) 296–306. [11] B.W. Carey, S. Markoulaki, J. Hanna, K. Saha, Q. Gao, M. Mitalipova,
R. Jaenisch, Reprogramming of murine and human somatic cells using a single polycistronic vector, Proc. Natl Acad. Sci. U. S. A. 106 (2009) 157–162.
[12] K. Woltjen, I.P. Michael, P. Mohseni, R. Desai, M. Mileikovsky, R. Hamalainen, R. Cowling, W. Wang, P. Liu, M. Gertsenstein, K. Kaji, H.K. Sung, A. Nagy, piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells, Nature 458 (2009) 766–770. [13] K. Kaji, K. Norrby, A. Paca, M. Mileikovsky, P. Mohseni, K. Woltjen,
Virus-free induction of pluripotency and subsequent excision of reprogramming factors, Nature 458 (2009) 771–775.
[14] H. Zhou, S. Wu, J.Y. Joo, S. Zhu, D.W. Han, T. Lin, S. Trauger, G. Bien, S. Yao, Y. Zhu, G. Siuzdak, H.R. Scholer, L. Duan, S. Ding, Generation of induced pluripotent stem cells using recombinant proteins, Cell Stem Cell 4 (2009) 381–384.
[15] L. Warren, P.D. Manos, T. Ahfeldt, Y.H. Loh, H. Li, F. Lau, W. Ebina, P.K. Mandal, Z.D. Smith, A. Meissner, G.Q. Daley, A.S. Brack, J.J. Collins, C. Cowan, T.M. Schlaeger, D.J. Rossi, Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA, Cell Stem Cell 7 (2010) 618–630. [16] J. Silva, O. Barrandon, J. Nichols, J. Kawaguchi, T.W. Theunissen, A.
Smith, Promotion of reprogramming to ground state pluripotency by signal inhibition, PLoS Biol. 6 (2008) e253.
[17] S. Eminli, J. Utikal, K. Arnold, R. Jaenisch, K. Hochedlinger, Reprogramming of neural progenitor cells into induced pluripotent stem cells in the absence of exogenous Sox2 expression, Stem Cells 26 (2008) 2467–2474.
[18] T. Aasen, A. Raya, M.J. Barrero, E. Garreta, A. Consiglio, F. Gonzalez, R. Vassena, J. Bilic, V. Pekarik, G. Tiscornia, M. Edel, S. Boue, J.C. Izpisua Belmonte, Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes, Nat. Biotechnol. 26 (2008) 1276–1284.
[19] Y. Amoh, M. Kanoh, S. Niiyama, Y. Hamada, K. Kawahara, Y. Sato, R.M. Hoffman, K. Katsuoka, Human hair follicle pluripotent stem (hfPS) cells promote regeneration of peripheral-nerve injury: an advantageous alternative to ES and iPS cells, J. Cell. Biochem. 107 (2009) 1016–1020.
[20] N. Sun, N.J. Panetta, D.M. Gupta, K.D. Wilson, A. Lee, F. Jia, S. Hu, A.M. Cherry, R.C. Robbins, M.T. Longaker, J.C. Wu, Feeder-free derivation of induced pluripotent stem cells from adult human adipose stem cells, Proc. Natl Acad. Sci. U. S. A. 106 (2009) 15720–15725.
[21] C. Li, J. Zhou, G. Shi, Y. Ma, Y. Yang, J. Gu, H. Yu, S. Jin, Z. Wei, F. Chen, Y. Jin, Pluripotency can be rapidly and efficiently induced in human amniotic fluid-derived cells, Hum. Mol. Genet. 18 (2009) 4340–4349.
[22] E. Galende, I. Karakikes, L. Edelmann, R.J. Desnick, T. Kerenyi, G. Khoueiry, J. Lafferty, J.T. McGinn, M. Brodman, V. Fuster, R.J. Hajjar, K. Polgar, Amniotic fluid cells are more efficiently reprogrammed to pluripotency than adult cells, Cell. Reprogram. 12 (2010) 117–125.
[23] M.S. Tsai, J.L. Lee, Y.J. Chang, S.M. Hwang, Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol, Hum. Reprod. 19 (2004) 1450–1456.
[24] M.S. Tsai, S.M. Hwang, Y.L. Tsai, F.C. Cheng, J.L. Lee, Y.J. Chang, Clonal amniotic fluid-derived stem cells express characteristics of both mesenchymal and neural stem cells, Biol. Reprod. 74 (2006) 545–551.
[25] M. Masip, A. Veiga, J.C. Izpisua Belmonte, C. Simon, Reprogram-ming with defined factors: from induced pluripotency to induced transdifferentiation, Mol. Hum. Reprod. 16 (2010) 856–868. [26] A. Meissner, Epigenetic modifications in pluripotent and
differentiated cells, Nat. Biotechnol. 28 (2010) 1079–1088. [27] M.S. Tsai, S.M. Hwang, K.D. Chen, Y.S. Lee, L.W. Hsu, Y.J. Chang, C.N.
Wang, H.H. Peng, Y.L. Chang, A.S. Chao, S.D. Chang, K.D. Lee, T.H. Wang, H.S. Wang, Y.K. Soong, Functional network analysis of the transcriptomes of mesenchymal stem cells derived from amniotic fluid, amniotic membrane, cord blood, and bone marrow, Stem Cells 25 (2007) 2511–2523.
[28] B.K. Kim, S.E. Kim, J.H. Shim, D.H. Woo, J.E. Gil, S.K. Kim, J.H. Kim, Neurogenic effect of vascular endothelial growth factor during germ layer formation of human embryonic stem cells, FEBS Lett. 580 (2006) 5869–5874.
[29] S. Da Sacco, S. Sedrakyan, F. Boldrin, S. Giuliani, P. Parnigotto, R. Habibian, D. Warburton, R.E. De Filippo, L. Perin, Human amniotic fluid as a potential new source of organ specific precursor cells for future regenerative medicine applications, J. Urol. 183 (2010) 1193–1200.
[30] A. Jezierski, A. Gruslin, R. Tremblay, D. Ly, C. Smith, K. Turksen, M. Sikorska, M. Bani-Yaghoub, Probing stemness and neural commitment in human amniotic fluid cells, Stem Cell Rev. 6 (2010) 199–214.
[31] J.M. Miranda-Sayago, N. Fernandez-Arcas, C. Benito, A. Reyes-Engel, J. Carrera, A. Alonso, Lifespan of human amniotic fluid-derived multipotent mesenchymal stromal cells, Cytotherapy 13 (2011) 572–581.
[32] C.M. Koch, C.V. Suschek, Q. Lin, S. Bork, M. Goergens, S. Joussen, N. Pallua, A.D. Ho, M. Zenke, W. Wagner, Specific age-associated DNA methylation changes in human dermal fibroblasts, PLoS One 6 (2011) e16679.
[33] A. Banito, S.T. Rashid, J.C. Acosta, S. Li, C.F. Pereira, I. Geti, S. Pinho, J.C. Silva, V. Azuara, M. Walsh, L. Vallier, J. Gil, Senescence impairs successful reprogramming to pluripotent stem cells, Genes Dev. 23 (2009) 2134–2139.
[34] P. De Coppi, G. Bartsch Jr., M.M. Siddiqui, T. Xu, C.C. Santos, L. Perin, G. Mostoslavsky, A.C. Serre, E.Y. Snyder, J.J. Yoo, M.E. Furth, S. Soker, A. Atala, Isolation of amniotic stem cell lines with potential for therapy, Nat. Biotechnol. 25 (2007) 100–106. [35] T.E. Ludwig, V. Bergendahl, M.E. Levenstein, J. Yu, M.D. Probasco, J.A.
Thomson, Feeder-independent culture of human embryonic stem cells, Nat. Methods 3 (2006) 637–646.
[36] T.E. Ludwig, M.E. Levenstein, J.M. Jones, W.T. Berggren, E.R. Mitchen, J.L. Frane, L.J. Crandall, C.A. Daigh, K.R. Conard, M.S. Piekarczyk, R.A.
Llanas, J.A. Thomson, Derivation of human embryonic stem cells in defined conditions, Nat. Biotechnol. 24 (2006) 185–187.
[37] M.H. Chin, M.J. Mason, W. Xie, S. Volinia, M. Singer, C. Peterson, G. Ambartsumyan, O. Aimiuwu, L. Richter, J. Zhang, I. Khvorostov, V. Ott, M. Grunstein, N. Lavon, N. Benvenisty, C.M. Croce, A.T. Clark, T. Baxter, A.D. Pyle, M.A. Teitell, M. Pelegrini, K. Plath, W.E. Lowry, Induced pluripotent stem cells and embryonic stem cells are distinguished by gene expression signatures, Cell Stem Cell 5 (2009) 111–123.