行政院國家科學委員會專題研究計畫成果報告
澱粉顆粒中磷脂質-直鏈澱粉錯合物的澱粉液化酵素感受性
Susceptibility of amylose-phospholipids complexes to hydr olysis by
alpha-amylase
計畫編號:NSC 87-2313-B-002-123
執行期限:86年08月01日至87年07月31日
主持人:呂廷璋 tjlu@ccms.ntu.edu.tw 執行機構及單位名稱:國立台灣大學
一、中文摘要 以細菌性α-澱粉液化酵素(α-amylase, E.C.3.2.1.1)降解小麥澱粉,在降解的過程中 澱粉顆粒的 x-射線繞射圖譜強度在 60%的 顆粒被降解後才有減弱的情形,同時部份降 解澱粉顆粒的直鏈澱粉含量隨降解的程度增 加只有小幅度的增加,在降解率 60%時才 有 3%的增加量,顯示α-澱粉液化酵素在降 解澱粉顆粒時對直、支鏈澱粉與澱粉顆粒的 結晶、非結晶區都沒有明顯的選擇性。由 31 P-核磁共振分析與總磷含量測定顯示,小 麥澱粉顆粒中的磷主要以磷脂質的形態存 在,當顆粒降解時,殘留顆粒中磷脂質的含 量隨降解的程度增加而增加,顯示澱粉顆粒 中含有磷脂質的部份對α-澱粉液化酵素感受 性較其他部份低,配合直鏈澱粉濃度的增加 量可以推斷因為磷脂質與直鏈澱粉形成錯合 物而降低了對酵素的感受性。 關鍵詞:小麥澱粉顆粒,α-澱粉液化酵 素,澱粉磷脂質,澱粉酵素降解。 Abstr actWheat starch granules was subjected to bacterialα-amylase (E.C.3.2.1.1) hydrolysis. α-amylase had not preference for neither components, amylose and amylopectin, nor regions, amorphous or crystalline. Changes of composition ratio of starch components and x-ray diffraction properties are minor comparing to the degradation rate. The amylose content
of residual starch granules increased only 3% and the intensity of x-ray diffraction also slightly altered as the 60% of starch granules had been eroded. According results of colorimetry and 31P-NMR analysis, phospholipids were the major form of phosphorous in starch granules. The phospholipids were resistant to enzymatic degradation comparing to other parts of starch granules. The results indicated that the
endogenous phospholipids in starch granules were complexing with amylose to decrease its
susceptibility to α-amylase.
Keywor ds: wheat starch granules, bacterial α -amylase, phospholipids, amylolysis.
二、緣由與目的
澱粉顆粒中含有少量的脂肪,在過去的研 究中較少被研究。在玉米澱粉顆粒含有少量 的游離脂肪酸(free fatty acids, FFA) [1],而 Lysolecithin 的含量以在高直鏈穀類澱粉樣 品中較高,而在糯性穀類澱粉樣品中則含很 少的量。因為 FFA 和 lysophospho-lipids(LPL)能和直鏈澱粉形成錯合物,因此 很自然的被認為在澱粉顆粒中也是以錯合物 的型態存在 [2, 3],這一個假設借由13 C-CP/MAS-NMR 分析及其它補助的證據已經 得到證實 [4, 5, 6]。 由於 FFA 和 LPL 與直鏈澱粉形成錯合
物,因此不易被氧化或被一般溶劑所抽出。 另一方面,這種錯合也使直鏈澱粉不易被熱 水從澱粉顆粒中萃取出來。由 Morrison 等 人一系列的報告可以知道 LPL 的存在直接 影響直鏈澱粉糊化和回凝時的特性。 大部分脂肪-直鏈澱粉錯合物為不水溶的 非結晶型態(type I), 但他們可以經由回火處 理(annealing)而變為部分結晶(semicrystalline form)的型態(type II)。在大部分穀類澱粉中 脂肪-直鏈澱粉錯合物為 type I,此錯合物若 在水中加熱至 94-100o C,則脂肪和直鏈澱 粉會解離而分開,若糊化後再形成的 type II 錯合物,則具有明顯的 X-ray V 型繞射圖 譜,若在水中加熱必須昇高溫度至 100 -125oC 才會解離 [7]。 在小麥、大麥、裸麥及 triticale 澱粉中所 含的脂肪幾乎都是 LPL,而其它的穀類則有 一定的 FFA 和 LPL 的組成比例 [8](表一)。 Morrison 並指出脂肪含量和直鏈澱粉的含量 有密切的關係,且直鏈澱粉存在於澱粉顆粒 的型態影響了澱粉的特性 [8]。此外,馬鈴 薯直鏈澱粉和脂肪酸及 lyso-phosphatidylcholine 在試管中所生成的錯合 物(type II)亦對葡萄醣澱粉分解酵素有很好 的抗性 [9]。 本計劃的目的是探討在澱粉顆粒中自然 的磷脂質-直鏈澱粉錯合物(type I)之澱粉液 化酵素感受性。小麥澱粉為主要的實驗原 料,因為其所含的澱粉脂肪絕大部分都是 LPL [8],因此可以利用比色法分析 [10] 和 31 P-NMR 分析 [11] 更精確的偵測其磷脂質 含量的在澱粉顆粒降解時的變化,借以瞭解 小麥中磷脂質-直鏈澱粉錯合物的感受性。 利用31 P-NMR 分析和比色法分析,雖然不 能直接測量磷脂質的種類和其組成百分比, 但另一方面也避免了,溶劑萃取與酸水解等 繁雜步驟所帶來的誤差。此外,也將配合掃 瞄式電子顯微鏡觀察澱粉顆粒降解時形態的 變化,試圖明瞭磷脂質-直鏈澱粉錯合物在 澱粉顆粒中的分佈情形。 三、材料與方法 1. 實驗材料:本實驗所選用Roquette公司 (Lille, France)的小麥澱粉為樣品。細菌 澱粉液化酵素(α-amylase, Bacillus sp.) 購自Sigma Chemicals (St. Louis, MO)。 2. 澱粉X-射線繞射圖譜:按照Zoble [12]
在Method in Carbohydrate Chemistry, Vol. IV 所記載的步驟記錄從2θ=4至30 degree的圖譜。 3. 澱粉降解反應:稱取2g澱粉樣品分散懸 浮於50ml 50mM磷酸鹽緩衝液中(α-澱 粉液化酵素,pH 6.9,含0.02% sodium azide 以抑制微生物生長),再加入10單 位(Units) α-澱粉液化酵素(1U為於20°C, pH 6.9下每3分鐘可釋出1.0 mg 麥芽 糖),均勻混合後,置於37°C恆溫振盪器 中,於適當間隔時間取出少量上澄清 液,以phenol-sulfuric acid法[13]測量可 溶性醣類總量,再以溶性醣類總量計算 出澱粉降解量。 4. 磷脂質含量分析:總磷含量分析按 Morrison [10]的方法進行將澱粉以濃硫 酸分解將磷脂質轉變為無機磷再以呈色 反應測定磷含量。磷的存在型式以31 P -NMR分析,按照Lim et al [11]敘述的方 法進行。31 P-NMR圖譜可以分辨磷原子 在不同的分子上,如磷脂質、無機磷和 澱粉分子上的磷酸單酯 [11, 14, 15] 。 5. 澱粉組成分膠體分子篩層析分析:選用 SepharoseCL-2B管柱(2.6 x 80 cm)為固定 相,以1 mM NaOH+25 mM NaCl為流動 相,流速30 ml/hr,注入樣品量為10 mg 並加入1 mg葡萄糖為標示物。 四、結果與討論 由31 P-NMR分析的結果顯示,所選用的小
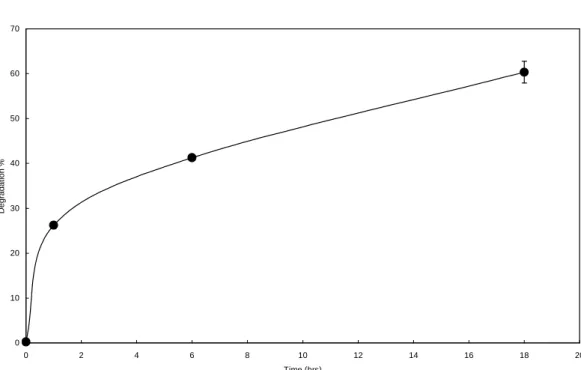
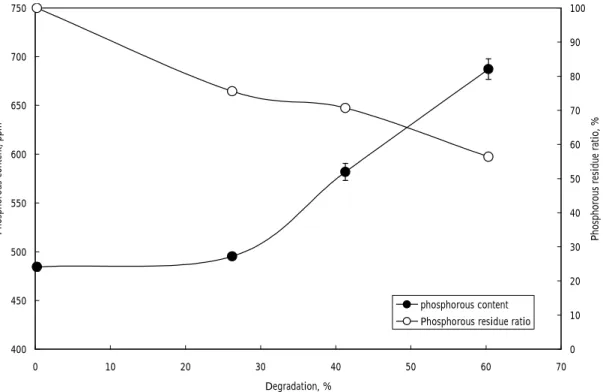
麥澱粉樣品中的磷主要以磷脂質的形態存在 (圖一),其chemical shift 在0~1.2 ppm 之 間,並含有~7%無機磷(inorganic phosphorous, Pi)。由比色分析法的結果顯 示所選用的小麥澱粉的總磷含量約為490 ppm,若依據Morrison [16]所提的公式計算 (LPL = P x 16.3)樣品中的LPL的含量約為 0.7%。以細菌(Bacillus sp.)α-amylase 降解 小麥澱粉顆粒時,殘留的顆粒中的磷含量, 隨降解程度的增加而增加,顯示在過程中含 有LPL的直鏈澱粉的降解速率較澱粉顆粒的 其它部分為慢,使LPL不易被釋出(圖二、 三)。若換算磷在殘餘的澱粉顆粒中的殘留 量,發現磷脂質的釋出率雖然較澱粉顆粒的 降解速率慢,但仍有一部分的磷脂質在酵素 降解過程中被釋出,其釋出比率與降解程度 略成直線關係(圖三)。由圖二的降解趨勢 可以觀察到澱粉顆粒在初期以較快的速度被 α-amylase所分解,之後速度變慢趨於平 緩,所能被快速降解的比例約為50至60%之 間。 由x-光線繞射分析結果顯示小麥澱粉具有 A型繞射圖譜,在α-amylase 降解過程其繞 射圖譜並沒有明顯的改變,經18小時的降解 反應後(60%的澱粉顆粒已被分解),繞射 圖譜的強度有減若的現象,顯示α-amylase 降解小麥澱粉顆粒時對顆粒的結晶與非結晶 區並沒有特定的選擇性,這和以酸水解的過 程不同,在降解率達到60%時因為大部分的 顆粒結構以被分解,因此繞射的強度也減 弱。 利用膠體分子篩層析分析可以按分子量大 小將澱粉的直鏈與支鏈澱粉組成分離,並觀 察在α-amylase 降解小麥澱粉顆粒時對顆粒 組成的變化。結果顯示在降解的過程中直鏈 與支鏈澱粉組成比例只有些微的增加,(圖 五、六),從原來的29.2%增加為32.3%,這 在一次證明α-amylase對直、支鏈澱粉並沒 有明顯的選擇性,且支鏈澱粉在分解過程中 並沒有出現大量的斷裂的小分子,所以利用 碘 染 色 後 並 無 最 大 吸 收 波 長 的 偏 移 ( 圖 六),這顯示在α-amylase 降解的過程終將 澱粉的生長環(growth ring)依次的一層一層 的分解。至於增加的直鏈澱粉的量,推測是 因為直鏈澱粉與磷脂質結合而受到保護,因 此不易被分解這和總磷量的變化結果有一致 性。 由本實驗的結果顯示所選用的小麥澱粉 樣品的磷主要存在於磷脂質中,因為這種內 生性的磷脂質和直鏈澱粉以錯合物的形態存 在,所以當澱粉顆粒被細菌性α-amylase降 解時,磷脂質被釋出的速率比澱粉顆粒降解 的速率慢,但由於磷脂質的含量很少,因此 仍有大部分的直鏈澱粉與支鏈澱粉以幾乎相 同的比例被α-amylase分解,由於降解時顆 粒間的差異很大因此上,無法確定磷脂質存 在澱粉顆粒的位置,若可將降解形態的澱粉 顆粒分離出來,將可以更精確的知道磷脂質 在小麥顆粒的分佈情形,是一個值得研究的 主題。 五、計畫成果自評 本計畫的執行整體而言按原計畫的內容 進行,在實驗的進行初期確立磷脂質的定量 方法時曾有一些困難,但結果顯示以比色分 析法配合31 P-NMR分析可以精確的測定澱粉 中的磷脂質含量,並可由31 P-NMR觀察到澱 粉中無機磷的存在並在計算中加以扣除,較 傳統以酸水解與溶劑萃取的方法不驟較簡 化,但由於31 P-NMR的樣品前處理較麻煩, 必須事先預備為其缺點。計畫的結果顯示磷 脂質在澱粉顆粒中的分佈並不均勻,且因與 直鏈澱粉形成錯合物的關係使直鏈澱粉不易 被澱粉液化酵素降解掉,這些結果使我們對 澱粉顆粒的結構又有多一分的認識。此外, 由結果顯示α-amylase對直、支鏈澱粉與澱 粉顆粒的結晶、非結晶區都沒有明顯的選擇 性,是與酸水解的過程不一樣的,更提供我 們 對 α -amylase降 解澱粉行為 的更一層 認 識。我們將會把本計畫的初步結果在今年十 二月份於國立屏東科技大學舉行的中華民國 食品科學技術學會第28屆年會中發表。
五、參考文獻
[1] Schoch, T. J. 1942. Non-carbohydrate substances in cereal starch. J. Am. Chem. Soc. 64:2954.
[2] Morrison, W. R. 1978. Cereal lipids. Adv. Cereal Sci. Technol. 2:221.
[3] Morrison, W. R. 1988. Lipids. In: Wheat Chemistry and Technology, 3rd ed. Vol. 1. Y. Pomeranz, Ed. American Association of Cereal Chemists, St. Paul, MN.
[4] Morrison, W. R., Law, R. V. and Snape, C. E. 1993. Evidence for inclusion complexes of lipids with V-amylose in maize, rice and oat starches. J. Cereal Sci. 18:107. [5] Morrison, W. R., Tester, R. F. and Gidley,
M. J. and Karkalas, J. 1993. Resistance to acid hydrolysis of lipid-complexed
amylose and lipid-free amylose in lintnerized waxy and non-waxy barley starches. Carbohdr. Res. 245:289.
[6] Morrison, W. R., Tester, R. F., Snape, C. E., Law, R. and Gidley, M. J. 1993. Swelling and gelatinization of cereal starches, IV. Some effects of lipid-complexed amylose and free amylose in waxy and normal barley starches. Cereal Chem. 70:385.
[7] Raphaelides, S. R. and Karkalas, J. 1988. Thermal dissociation of amylose-fatty acid complexes. Carbohydr. Res. 172:65. [8] Morrison, W. R. 1995. Starch lipids and
how they relate to starch granule structure and functionality. Cereal Foods World 40:437.
[9] Kitahara, K., Suganuma, T. and Nagahama, T. 1996. Susceptibility of amylose-lipid complexes to hydrolysis by glucoamylase
from Rhizopus niveus. Cereal Chem. 73:
428-432.
[10] Morrison, W. R. 1964. A fast, simple and reliable method for the
microdetermination of phosphorous in biological materials. Anal. Biochem. 7:218. [11] Lim, S.-T., Kasemsuwan, T. and Jane, J.
1994. Characterization of phosphorus in
starch by 31P-Nuclear Magnetic Resonance
Spectroscopy. Cereal Chem. 71:488-493. [12] Zobel, H. F. (1964) X-ray analysis of
starch granules. In “Methods in Carbohydrate Chemistry.” Vol. IV, p. 109-113. Ed. Whistler, R.L. et al., Academic Press, N. Y., NY.
[13] Dubois, M., Gilles, K. A., Hamilton, J.K., Rebers, P. and Smith, F. (1956)
Calorimetric method for determination of sugar and related substance. Anal. Chem. 28: 350-356.
[14] Lim, S. and Seib, P. A. 1993. Location of phosphate esters in a wheat starch
phosphate by 31P-nuclear magnetic
resonance spectroscopy. Cereal Chem. 70:145-152.
[15] Bay-Smidt, A. M., Wischmann, B., Olsen, C. E. and Nielsen, T. H. 1994. Starch bound phosphate in potato as studied by a simple method for determination of
organic phosphate and 31P-NMR. Starch
46:167-172.
[16] Morrison, W. R. 1988. Starch lipids: A review. J. Cereal Sci. 8:1.
Figure 1. 31P-nuclear magnetic resonance spectrum of wheat starch.
Figure 2. Time course of wheat starch degradation by bacterial α-amylase.
0 10 20 30 40 50 60 70 0 2 4 6 8 10 12 14 16 18 20 Time (hrs) Degradation %
Figure 3. Change in phosphorous content in wheat starch during degradation of bacterial α -amylase.
Figure 4. X-ray diffratograms of wheat starches degraded by bacterial α-amylase for different
period.
4 9 14 19 24 29 2-theta degree
Itensity (arbitary unit)
intact 6 hrs 18 hrs 400 450 500 550 600 650 700 750 0 10 20 30 40 50 60 70 Degradation, % Phosphorous content, ppm 0 10 20 30 40 50 60 70 80 90 100
Phosphorous residue ratio, %
phosphorous content Phosphorous residue ratio
Figure 5 Sepharose CL-2B chromatogram of wheat starch.
Figure 6. Sepharose CL-2B chromatogram of wheat starch residues after 18 hours amylolysis.
0 50 100 150 200 10 20 30 40 50 60 70 80 90 100 Fraction No.
Carbohydrate concentration ug/ml
a Glucose 0 50 100 150 200 250 10 20 30 40 50 60 70 80 90 100 Fraction No.
Carbohydrate concentration ug/ml
0 100 200 300 400 500 600 700 800 (nm) Glucose λ max