行政院國家科學委員會專題研究計畫 期中進度報告
磷酸化酪胺酸的訊息傳遞對角膜內皮細胞生理機制的影響
(1/3)
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-002-091- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院眼科 計畫主持人: 胡芳蓉 共同主持人: 陳偉勵 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 5 月 3 日
磷酸化酪氨酸的訊息傳遞對角膜內細胞生理機制的影
響---第一年精簡報告
Abstract
Purpose: Contact inhibition is an important mechanism for maintaining corneal endothelium in a non-replicative state. Protein tyrosine phosphatases (PTPs) play a role in regulating the integrity of cell-cell contacts, differentiation, and growth. In this study, we aimed to evaluate whether phosphatases are involved in the maintenance of contact-dependent inhibition of proliferation in corneal endothelial cells and to identify candidate PTPs that are expressed in these cells and might be involved in regulation of contact inhibition.
Methods: Confluent cultures of rat corneal endothelial cells or endothelium in ex
vivo corneas were treated with the general phosphatase inhibitor, sodium
orthovanadate (SOV). Immunocytochemistry (ICC) evaluated the effect of SOV on cell-cell contacts by staining for ZO-1, and on cell cycle progression by staining for Ki67. Transverse sections of rat cornea and cultured rat corneal endothelial cells were used to test for expression of the candidate PTPs: PTP-mu, PTP-LAR, PTP1B, SHP-1, SHP-2, and PTEN using ICC and either Western blots or RT-PCR.
Results: ZO-1 staining demonstrated that SOV induced a time-dependent release of cell-cell contacts in confluent cultures of corneal endothelial cells and in the
endothelium of ex vivo corneas. Staining for Ki67 indicated that SOV promoted limited cell cycle progression in the absence of serum. PTP-mu, PTP1B, SHP-1,
SHP-2, and PTEN, but not PTP-LAR, were expressed in rat corneal endothelial cells
in situ and in culture. The subcellular location of PTP-mu and PTP1B differed in
subconfluent and confluent cells, while that of SHP-1, SHP-2, and PTEN was similar, regardless of confluent status. Western blots confirmed the expression of PTP1B, SHP-1, SHP-2, and PTEN. RT-PCR confirmed expression of PTP-mu mRNA.
Conclusion: Phosphatases are involved in regulation of junctional integrity and of cell proliferation in corneal endothelial cells. PTP-mu, PTP1B, SHP-1, SHP-2, and PTEN are expressed in rat corneal endothelium and may be involved in regulation of contact inhibition in these normally non-proliferating cells.
2. Materials and Methods
2.1. Antibodies and Reagents
Rabbit-anti-human SHP-1, rabbit-anti-human SHP-2, goat-anti-rat PTP-LAR, mouse-anti-human PTEN, horseradish peroxidase-conjugated donkey-anti-rabbit IgG, goat-anti-mouse IgG, and donkey-anti-goat IgG, as well as blocking peptides specific for SHP-1, SHP-2, PTP-LAR and PTEN, were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA). Mouse anti-human-PTP1B was obtained from Oncogene Sciences (Cambridge, MA). Polyclonal rabbit-anti-human zonula
occludins-1 (ZO-1) and mouse-anti-human Ki67 were purchased from Zymed
Laboratories (San Francisco, CA). Monoclonal antibody against the intracellular (SK7) domains ofhuman PTP-mu was kindly provided by Dr. Susan M. Brady-Kalnay
(Case Western Reserve University, Cleveland, OH). Rabbit anti-human non-muscle myosinwas obtained from Biomedical Technologies, Inc. (Stoughton, MA). Sodium orthovanadate (SOV), ethylene glycol-bis (β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid (EGTA), ethylenediaminetetraacetic acid (disodium EDTA), and 100X
antibiotic/antimycotic solution were from Sigma (St. Louis, MO). FITC-conjugated donkey anti-rabbit, anti-mouse, and anti-goat secondary antibodies and donkey serum were purchased from Jackson ImmunoResearch (West Grove, PA). Mounting medium containing propidium iodide (PI) was purchased from Vector Laboratories
(Burlingame, CA). Medium-199 and gentamicin were obtained from Invitrogen/Life Technologies (Carlsbad, CA). Fetal bovine serum (FBS) was purchased from Atlanta Biologicals (Norcross, GA). Bovine serum albmin (BSA) was purchased from Fisher Scientific (Pittsburgh, PA).
2.2. Culture of Rat Corneal Endothelial Cells
Corneas were obtained from adult male Sprague-Dawley rats, which were treated in accordance with the ARVO Statementfor the Use of Animals in Ophthalmic and Vision Research. Endothelial cells were grown in explant culture as previously described (Chenet al., 1999). Toremove the epithelial cell layers, corneas were incubated for1 hr at 37°C in 2.5 mM EDTA to disrupt hemidesmosomes, followed by gentle removal of the epithelium. For explant culture of theendothelium, corneas were cut in half and placed endothelium-sidedown in a 6-well tissue culture plate. Pieces were allowed toattach to the culture plate for approximately 5 min,after which 1 drop of culture medium was placed over the tissue. Culture medium consisted of Medium-199, 50 µg/ml gentamicin, and 10% FBS. Corneal pieces were incubated overnightat 37°C in a 5% CO2 humidified atmosphere.On the following day, 1 ml culture medium was gently added andcultures were incubated undisturbed. After 5 days,2 ml of medium were added per well. Medium was changed every otherday thereafter. After approximately 10 days, when a sufficientnumber of endothelial cells had migrated off the cornea, thecorneal pieces were carefully removed, and the remaining endothelialcells were grown to confluence. Primary cultured cells were then trypsinized, resuspended in culture medium, and seeded into 2-well chamber slides or into T75 flasks at a density of 2.5×105
cells per flask.
2.3. Immunocytochemical Localization
For immunocytochemical (ICC) analysis of cultured cells, cells were seeded onto chamber slides and either used as a non-confluent culture or grown to confluence. After washing three times with phosphate-buffered saline (PBS), cells were fixed in methanol at –20°C for 15 min followed by a further wash with PBS. The fixed cells were then permeabilized with 1% Triton X-100 in PBS for 10 min. After a PBS wash,
cells were pre-incubated for 10 min in 2% BSA in PBS (5% donkey serum when goat antibody was used) to block non-specific binding, and then incubated with primary antibody at room temperature for 2 hrs. Concentrations of primary antibodies were as follows: anti-ZO-1=1:150; anti-Ki67=1:100; anti-PTP-mu=1:400;
anti-PTP-LAR=1:200; anti-PTP1B=1:100; anti-SHP-1=1:400; anti-SHP-2=1:400; and anti-PTEN=1:100. According to the suppliers, all primary antibodies used in these studies were cross-reactive with the appropriate rat antigen. Following primary
antibody incubation, cells were washed with PBS, incubated for 10 min with blocking buffer, and then incubated for 1 hr with a 1:200 dilution of the appropriate
FITC-conjugated secondary antibody. To assess specificity of the staining, cells were either processed without primary antibody or incubated with primary antibody that had been pre-absorbed overnight with a 5X concentration of blocking peptide. Cells were then washed with PBS and mounted in medium containing propidium iodide (PI) for visualization of nuclei. All experiments were repeated three times to ensure
consistent results. Immunolocalization in the endothelium of ex vivo corneas was the same as in cultured cells except that 4% BSA in PBS was used in the blocking buffer. (I will insert a description of our new confocal microscope.)
ICC studies were also conducted using transverse sections of rat cornea. Corneas were obtained from adult male Sprague-Dawley rats. Corneas were fixed, sectioned, and immunostained according to published methods (Joyce et al, 1998). The
localization pattern in both cultured cells and transverse sections was observed by fluorescence microscopy using an Eclipse E800 Nikon Microscope with a VFM Epi-Fluorescence Attachment (Nikon Inc., Melville, NY) equipped with a Spot digital camera and Spot version 1.1 CE software (Diagnostic Instruments, Sterling Heights, MI). Experiments were conducted using tissue sections from three different rat corneas to ensure consistent results.
2.4. Protein Extraction and Immunoblotting
Cultured cells grown in T75 flasks were trypsinized, suspended in buffer containing 1% Triton X-100, 250 mM NaCl, 2 mM EDTA, 50 mM Tris-HCl, 10 µg/ml leupeptin, and 1 mM phenylmethylsulfonyl fluoride (all from Sigma), and homogenized. Equal amounts of extracted protein were loaded on 4-12%
polyacrylamide gels (Invitrogen, Carlsbad, CA) for SDS-PAGE. After electrophoresis, proteins were transferred to polyvinylidene difluoride (PVDF) membranes (Millipore Corp., Bedford, MA). Non-specific binding was blocked by incubation overnight at 4°C with 5% nonfat dry milk in PBS. Membranes were then incubated at room temperature for 2 hrs with anti-PTP-LAR, anti-PTP-1B, anti-SHP-1, anti-SHP-2 or anti-PTEN primary antibodies at the same dilutions indicated for ICC, with or without pre-incubation with blocking peptide. Anti-non-muscle myosin was used as a positive control for the immunoblot. Membranes were washed three times with the blocking buffer and then incubated with the appropriate horseradish peroxidase-conjugated secondary antibody at a final dilution of 1:10,000. After final washes with 0.1% Triton X-100 in PBS, signal was detected by enhanced chemiluminescence following
manufacturer’s instructions (Pierce, Rockford, IL) and exposed to autoradiographic film. All experiments were repeated three times to ensure consistent results.
2.5. RT-PCR of PTP-mu
For RT-PCR studies, total RNA was prepared from confluent culturesof rat corneal endothelial cells according to the manufacturer’sdirections (TRIzol; Gibco, Gaithersburg,MD). cDNA was prepared from 1 µgtotal RNA by reverse transcription in a volume of 20 µl using reagents froma commercially available kit (Promega, Pittsburgh, PA). Primersspecific for PTP-mu were designed using MacVector5.0
software (Oxford Molecular Group, Oxford, UK). Primer sequences were based on human PTP-mu sequences, since no sequence of rat PTP-mu was available in the GenBank database. In the region chosen for synthesis of the primers, mouse PTP-mu sequences only differed from the human sequence by one nucleotide, suggesting that these primers would recognize rat PTP-mu sequences.The primer pairs used were: upstream sequence, 5'-TCCTATTACCCAGATGAAACCCAC-3';downstream sequence, 5'-CCACTTCCACAAGATTGGTCACC-3'. PCR was performed in a reaction mixture containing 1 µgcDNA and 2.5 µM each of the upstream and downstream primers,plus reagents from a commercially available kit (Invitrogen). Specificityand yield of the PCR products were enhanced using the hot-startapproach (Erlich et al., 1991). PCR was performed for40 cycles in a thermal cycler. Cycle conditions included denaturationat 95°C for 1 min, annealing at 50°C for 1 min, and extension at 72°C for 2 mins. A 5-minextension was added at the end of the 40 cycles of PCR. PCR products and 100-bpDNA ladder molecular weight markers were electrophoresed in1.5% agarose gels containing 0.5 µg/ml ethidium bromideand photographed. Glyceraldehyde-3-phosphate dehydrogenase (G3PDH;Clontech, Palo Alto, CA) acted as a positive control for the PCR.Negative control samples consisted of the PCR reaction mixture, includingprimers, but without cDNA. To ensure that the total RNA sampleswere not contaminated with genomic DNA, a negative control using1 µg total RNA was substituted for cDNA in the PCR reaction mixture,along with 2.5 µM each of upstream and downstream G3PDH primers.All experiments were repeated three times to ensure consistent results.
3. Results
Endothelial Cells and in the Endothelium of Ex Vivo Corneas
The phosphatase inhibitor, SOV, disrupted cell-cell junctions and triggered cell cycle entry in contact-inhibited human umbilical vein endothelial cells (Suzuki et al., 2000). To test whether phosphatase inhibition could have similar effects on confluent rat corneal endothelial cells, we first examined the effect of SOV on cell-cell contacts. Cells were grown to confluence and maintained an additional week to ensure the formation of a mature, contact-inhibited monolayer. In a preliminary dose-dependent study, the concentration of 50 µM SOV was found to optimally release cell-cell
contacts without inducing cell loss or apoptosis. To investigate time-dependent effects, confluent cultures were incubated for 24, 48, or 72 hrs in 50 µM SOV plus
Medium-199 and antibiotics as described in Materials & Methods, then fixed and immunostained for ZO-1, a tight junction-associated protein (Siliciano and
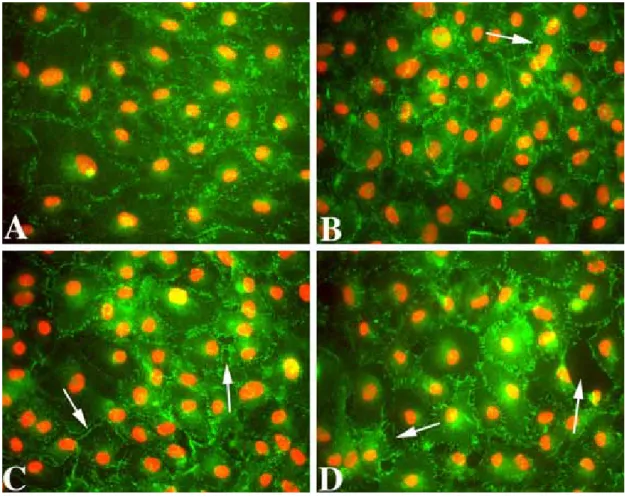
Goodenough, 1988), to help visualize cell borders. ZO-1 staining in control confluent cultures appeared as a single, discontinuous linear pattern at cell borders (Fig. 1A), indicating intact cell-cell contacts. Treatment of confluent cultures with SOV induced a time-dependent release of cell-cell contacts, resulting in the appearance of gaps between cells and a gradual loss of monolayer integrity (Fig. 1B-D). Secondary antibody controls were negative, indicating the specificity of the ZO-1 antibody staining (data not shown).
Figure 1. Representative images of immunolocalization of ZO-1 in fully confluent cultures of rat corneal endothelial cells. Control cultures were maintained minus FBS (A). Other cultures were incubated in 50 µM SOV without FBS for 24 hrs (B), 48 hrs (C), or 72 hrs (D) prior to immunostaining. Arrows in B-D indicate breaks in the confluent monolayer. Green: ZO-1. Red: PI. Original magnification = 40X.
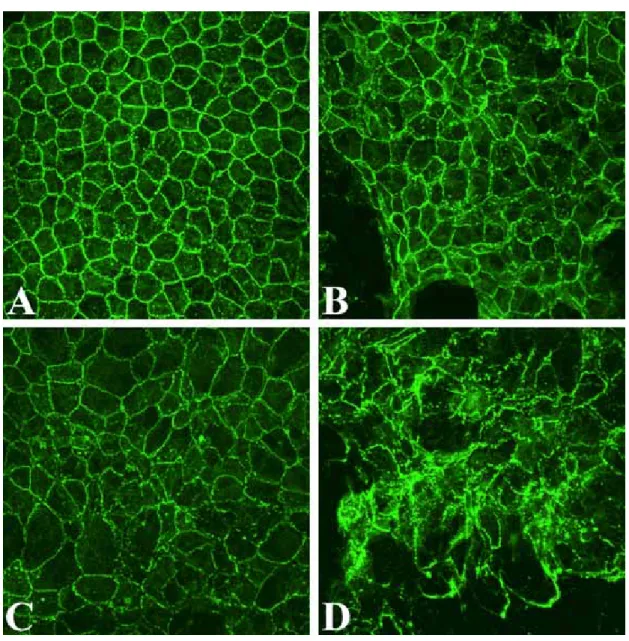
Similar immunolocalization studies were conducted in ex vivo rat corneas to determine whether SOV would disrupt the endothelial monolayer in situ, thus demonstrating relevance of the results obtained using the cultured cell model. Figure 2 presents representative confocal microscopic images of ZO-1 staining in rat corneal endothelium treated with 50 µM SOV for 24, 48, or 72 hrs. Untreated controls in Fig. 2A demonstrate the discontinuous linear pattern typical for the intact
endothelial monolayer. SOV treatment induced a time-dependent loss of cell-cell contacts, a general disruption of monolayer integrity, and loss of normal polygonal
cell shape (Fig. 2B-D), indicating that inhibition of phosphatase activity by SOV disrupts the endothelial monolayer in situ, as well as in culture.
Figure 2. Immunolocalization of ZO-1 in the endothelium of ex vivo corneas incubated in the absence of SOV (A) and in the presence of 50 µM SOV for 24 (B), 48 (C), or 72 hrs (D). PI staining is not shown in order to permit clear visualization of ZO-1. Original magnification: 80 X.
3.2. Phosphatase Inhibition Promotes LimitedCell Cycle Progression in Confluent Rat Corneal Endothelial Cells
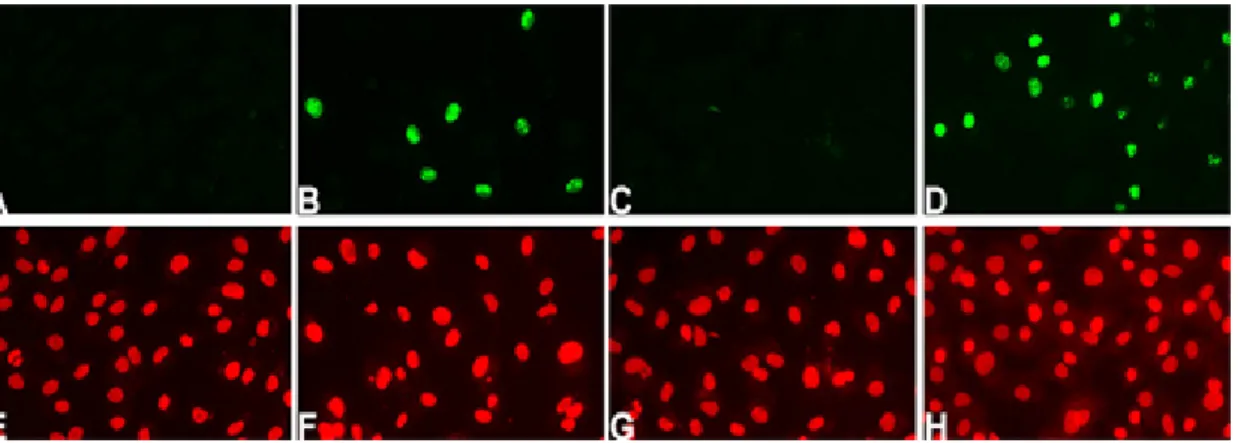
cultures of rat corneal endothelial cells. For this study, fully confluent cultures were treated and incubated for 48 hrs as indicated below, then fixed and immunostained for Ki67, a marker of actively cycling cells (Gerdes et al., 1991). Representative images of the results are presented in Figure 3. As expected, confluent cultures maintained for 48 hrs in Medium-199 and antibiotics without FBS did not reveal positive staining for Ki67 (Fig 3A). Positive Ki67 staining was detected in some cells within confluent cultures treated with 50 µM SOV (Fig. 3B). We also tested whether release of cell-cell contacts by itself was sufficient to induce cell cycle entry. To answer this question, we compared the effect of SOV and the calcium chelator, EGTA, since chelation of
calcium releases cell-cell contacts in corneal endothelium (Stern, 1981; Senoo, 2000b). Confluent cultures were pre-treated for 30 minutes with 5 mM EGTA and then
incubated for 48 hrs in the absence or presence of FBS. As seen in Fig. 3C, cultures treated with EGTA in the absence of FBS did not show Ki67-positive cells. As
expected, cells did enter the cell cycle when EGTA-treated cultures were incubated in the presence of 10% FBS (Fig. 3D). These results indicate that disruption of cell-cell contacts alone was not sufficient to induce cell cycle entry and that positive Ki67 staining, such as that seen in Fig. 3B, must result from SOV-induced phosphatase inhibition. Ki67 staining patterns change with cell cycle position (Starborg, 1996) and this change in pattern has been used to document cell cycle progression in human corneal endothelial cells in an ex vivo cornea wound model (Senoo, 2000a).
Interestingly, Ki67 staining patterns in SOV-treated cells (Fig. 3B) appeared to differ from those in cells treated with EGTA in the presence of FBS (Fig. 3D).
EGTA-treated cells showed a variety of patterns from G1-phase through mitosis, whereas, a single G1/S-phase pattern was observed in SOV-treated cells.
Figure 3. Immunolocalization of Ki67 in confluent cultures of rat corneal endothelial cells incubated minus FBS (A, E); in 50 µM SOV alone (B, F); in 5 mM EGTA alone (C, G); or in 5 mM EGTA plus 10% FBS (D, H). Ki67 staining (green) is shown in (A-D); PI (red) in (E-H) shows nuclear staining in the same microscopic field. Original magnification = 400X.
Studies were also conducted in which the endothelium of ex vivo corneas was treated with 50 µM SOV for 24, 48, and 72 hrs followed by immunostaining for Ki67 or BrdU. Positive Ki67 staining was observed in some endothelial cells in
SOV-treated corneas; however, there was no evidence of a time-dependent increase in the relative number of Ki67-positive cells (data not shown). No positive BrdU
staining was observed at any time-point. Together, results from both cultured cells and ex vivo corneas indicate that SOV is capable of inducing limited G1-phase progression in endothelial cells, but that phosphatase inhibition is not sufficient by itself to induce entry into S-phase of the cell cycle.
3.3. Detection of Candidate PTPs in Rat Corneal Tissue
Many PTP isoforms have been identified in mammalian cells, but not all tissues express all isoforms. Since phosphatase inhibition by SOV resulted in loss of cell-cell contacts and induced limited cell cycle entry in confluent rat corneal endothelial cells, we attempted to identify specific candidate PTPs known to mediate these functions.
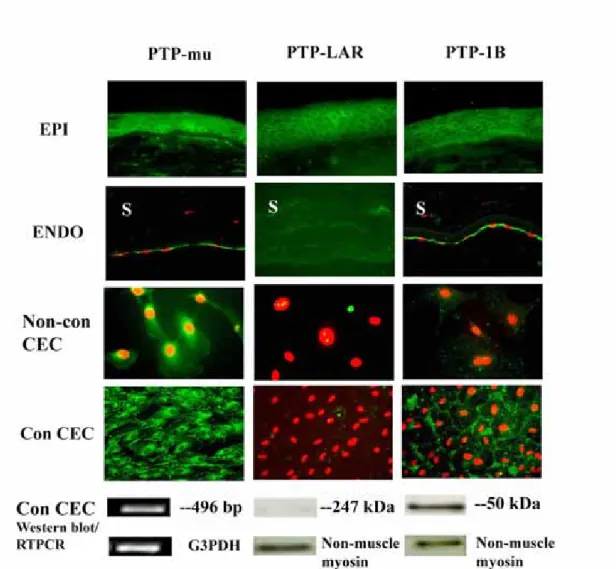
PTP-mu and PTP-LAR (leukocyte antigen-related PTP) are cell contact-associated PTPs. PTP-mu (Brady-Kalnay et al., 1993, 1998) and PTP-LAR (Aicher et al., 1997) are receptor-like transmembrane proteins. PTP1B is a non-receptor protein frequently associated with the endoplasmic reticulum (Haj, 2002) and vesicles subjacent to the plasma membrane. PTP1B appears to help mediate cell adhesion through regulation of cadherin-catenin complexes (Balsamo et al., 1996, 1998; Rhee, et al., 2001; Xu, 2002) and integrin signaling (Liu, 1998), as well as to regulate phosphorylation of the EGF, PDGF, and insulin receptors (Tonks, 2003). We used ICC to determine the relative expression of these PTPs in transverse sections of rat cornea. In Figure 4, we show that PTP-mu, PTP-LAR and PTP1B are expressed at the cell periphery in all layers of the corneal epithelium. Intense staining of PTP-mu was particularly evident in the lateral domains of basal epithelial cells. PTP-mu and PTP1B, but not PTP-LAR, are expressed in rat corneal endothelium. Although no nuclear staining was evident, it was not possible to determine in transverse corneal sections whether PTP-mu or PTP1B were present throughout the cytoplasm or at cell borders. For that reason, the staining patterns were also determined in cultured cells. In non-confluent cultured rat corneal endothelial cells, a predominantly perinuclear distribution of PTP-mu was found. There was no staining for PTP-LAR and little-to-no staining for PTP1B. When the cultured cells became confluent, PTP-mu was localized to both the perinuclear region and the cell periphery. Intense positive staining for PTP1B was observed mainly toward the cell periphery. Controls using secondary antibody alone yielded no positive staining (data not shown). As shown in Figure 4, Western blots confirmed the expression of PTP1B protein and lack of expression of PTP-LAR. Expression of non-muscle myosin was tested in the same blots as a positive control. The PTP-mu monoclonal antibody used in these studies specifically recognized the intracellular domain of PTP-mu (according to Dr. Susan Brady-Kalnay Case Western Reserve
University, Cleveland, OH). Previously this antibody functioned well in immunoblots of an LNCaP prostate carcinoma cell line (Hellberg, 2002); however, it did not work in Western blots of rat corneal endothelial cells. As such, we chose RT-PCR as a method to verify the expression of PTP-mu in rat corneal endothelial cells. As seen in Figure 4, a positive band of the expected size was obtained using primers specific for PTP-mu. No positive bands were observed for the PCR negative controls described in the Materials and Methods section (data not shown). G3PDH, the positive PCR control, yielded a band of the appropriate size.
ICC studies of rat corneal sections confirm the expression of PTP-mu, PTP-LAR and PTP1B (green) in corneal epithelium, and PTP-mu and PTP1B (green) in corneal endothelium. PI staining (red) indicates nuclei. RT-PCR confirmed the expression of PTP-mu mRNA, while Western blots confirmed the expression of PTP1B protein and lack of expression of PTP-LAR in cultured corneal endothelial cells. G3PDH acted as the positive control for the RT-PCR. Non-muscle myosin was the positive control for the Western blots. EPI=epithelium, ENDO=endothelium, Non-con
CEC=non-confluent rat corneal endothelial cells. Con CEC=confluent corneal
endothelial cells. S=stroma. Original magnification = 100X for EPI, 200X for ENDO, 600X for Non-con CEC, 400X for Con CEC.
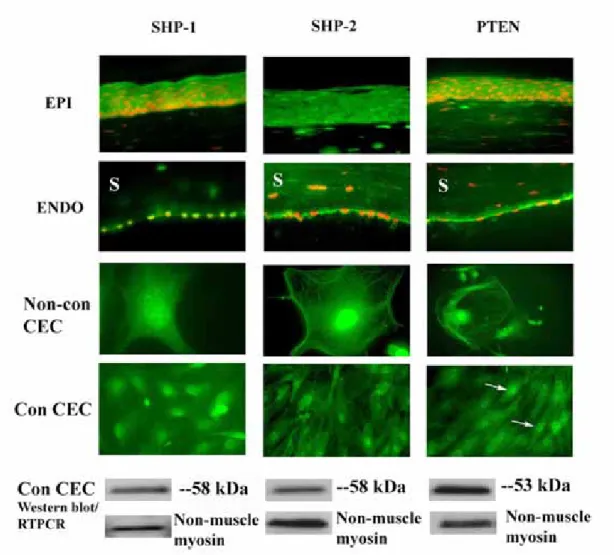
SHP-1, SHP-2, and PTEN, as well as PTP1B, are growth factor- and cell-cycle associated PTPs (Keilhack et al., 1998; Yu et al., 1998; Besson et al., 1999; Qi et al., 1999; Qu et al., 1999; Ramaswamy et al., 1999; Tonks, 2003). SHP-1 and SHP-2 are cytosolic PTPs that help regulate the function of receptor tyrosine kinases, such as the EGF and PDGF receptors. PTEN is a cytosolic dual-specificity phosphatase, i.e., it regulates serine and threonine, as well as tyrosine phosphorylation. It is also able to dephosphorylate lipids (Machama, 1998; Myers, 1998). Its function, in part, is to regulate signaling via the PI3K/Akt pathway. In Figure 5, we show that SHP-1, SHP-2 and PTEN were expressed in the cytoplasm within the entire rat corneal epithelium. Staining for SHP-1, SHP-2 and PTEN was also seen in the corneal endothelium. In cultured cells, SHP-1, SHP-2 and PTEN antibodies labeled subconfluent and confluent rat corneal endothelial cells, and the staining patterns were similar under both conditions. There was intense nuclear staining with moderate cytoplasmic
staining for all three PTPs. The staining was specific for SHP-1, SHP-2, and PTEN, as it was completely blocked when primary antibodies were pre-incubated with the corresponding antigen peptide (data not shown). Western blot analysis confirmed the expression of SHP-1, SHP-2 and PTEN protein in confluent rat corneal endothelial cells. For all three PTPs, positive reaction disappeared after pre-incubating the
primary antibodies with specific blocking peptides (data not shown). As previously, non-muscle myosin acted as a positive control for the Western blot.
Figure 5. Expression of SHP-1, SHP-2, and PTEN in rat corneal cells. ICC studies confirm the expression of SHP-1, SHP-2 and PTEN (green) in corneal epithelium, corneal endothelium, confluent and non-confluent rat corneal endothelial cells. PI staining (red) indicates nuclei. Arrows indicate punctate staining in the nucleus of PTEN in confluent corneal endothelial cells. Western blots confirmed the ICC results. Non-muscle myosin was used as a positive control for Western blots in all
experiments. EPI=epithelium, ENDO=endothelium, Non-con CEC=non-confluent corneal endothelial cells. Con-CEC=confluent corneal endothelial cells. S=stroma. Original magnification = 100X for EPI, 200X for ENDO, 800X for Non-con CEC, 400X for Con CEC.
4. Discussion
The present study provides evidence that SOV, a general phosphatase inhibitor, can both disrupt junctional integrity and promote limited cell cycle progression in confluent rat corneal endothelial cells both in culture and in ex vivo corneas. Results of our studies on cell-cell contact are consistent with those of others in which treatment with SOV interfered with normal cell-cell adhesion (Tsukita et al., 1991; Sorby and Ostman, 1996; Takahashi and Suzuki, 1996; Young et al., 2003). Since a phosphatase inhibitor was capable of disrupting cell-cell junctions, our results suggest that there must be a constitutive phosphatase activity in the confluent corneal
endothelial monolayer that helps maintain the integrity of these junctions. Exposure of confluent cells to SOV also promoted limited cell cycle progression in the absence of serum, but was not sufficient to induce S-phase entry. The fact that phosphatase inhibition promoted at least limited cell cycle entry in the absence of serum strongly suggests that constitutive phosphatase activity must help mediate the suppression of cell cycle entry in the endothelial monolayer. We previously demonstrated the importance of cell-cell contacts in mediating inhibition of proliferation in confluent cultures of rat corneal endothelial cells (Joyce, Harris and Mello, 2002) and in ex vivo human corneal endothelium (Senoo et al., 2000b). The fact that SOV was able to both alter the integrity of cell-cell contacts and promote limited cell cycle progression strongly suggests that phosphatases are involved in regulating contact-induced inhibition of proliferation in corneal endothelium; however, further study will be needed to confirm this.
It is well known that protein tyrosine phosphorylation and dephosphorylation are important physiological mechanisms that control diverse cellular behaviors, such as cell growth, differentiation, adhesion, and migration (Pallen and Tong, 1991;
and Sap, 2000). As such, our intriguing findings led us to initiate studies to identify PTPs expressed in rat corneal endothelial cells. This identification is an important first step in determining how the activity of specific phosphatases contributes to the regulation of contact inhibition in these cells. Our data indicates that PTP-mu, PTP1B, SHP-1, SHP-2, and PTEN are expressed in rat corneal endothelial cells both in culture and in situ.
The PTP family can be divided into receptor-like transmembrane proteins, such as PTP-mu, PTP-κ, PTP-λ, PTP-LAR, PTPα and CD45, and soluble cytosolic proteins, including SHP-1, SHP-2, and PTP1B (Barford et al., 1998). There are also dual-specificity phosphatases, such as PTEN, CDC25, and MAP kinase phosphatase-1, which catalyze the dephosphorylation of both protein serine/threonine and tyrosine kinases (Barford et al., 1998). We report here the identification of candidate PTPs that are members of each of these groups.
The receptor-like transmembrane PTP-mu and the non-receptor phosphatase, PTP1B can interact with proteins that comprise adhesion junctions, such as cadherins and catenins. Previous studies have demonstrated the expression of N-cadherin (Beebe and Coats, 2000; Ickes, et al. 2002) and α-, β-, and γ-catenin (plakoglobin) (Petroll, et al. 1999) in adhesion junctions of corneal endothelial cells, providing a rationale for examining the expression of these PTPs. PTP-mu interacts with E-, N- and R-cadherin (Brady-Kalnay et al., 1998). PTP1B has been shown to be essential for both N-cadherin and beta-1 integrin-mediated adhesion in some cell types (Balsamo et al., 1996; Balsamo et al., 1998; Pathre et al., 2001; Rhee et al., 2001). Immunolocalization of these PTPs in cross-sections of rat cornea indicated that PTP-mu and PTP1B are expressed in both the epithelium and endothelium, whereas, PTP-LAR was only expressed in the epithelium. Of interest was the observation of differences in the relative localization of PTP-mu and PTP1B in subconfluent and
confluent cells. Their peripheral localization in the intact endothelium suggests that both PTP-mu and PTP1B may be involved in the regulation of the integrity of junctional complexes.
The non-receptor PTPs, SHP-1 and SHP-2, as well as PTP1B, and the
dual-specificity phosphatase, PTEN are among phosphatases known to help regulate proliferation in other cell types. Several positive growth factors, such as EGF, FGF, and PDGF, can trigger proliferation in corneal endothelial cells (Gospodarowicz et al., 1977; Kamiyama et al., 1995). SHP-1, SHP-2, and PTP1B help regulate the function of various growth factor receptors (Keilhack et al., 1998; Yu et al., 1998; Qi et al., 1999; Qu et al., 1999; Tonks, 2003). SHP-1 and PTP1B are generally considered negative regulators of cell signaling, while SHP-2 is largely considered a positive signal transducer (Marrero et al., 1998; Haj, 2002). PTEN has been shown to negatively regulate cell cycle progression through the PI3K/Akt pathway (Li et al., 1997; Besson et al., 1999; Paramio et al., 1999; Ramaswamy et al., 1999). All four of these PTPs were expressed in rat corneal endothelium in situ and in culture. Unlike with PTP-mu and PTP1B, SHP-1, SHP-2, and PTEN did not demonstrate a significant change in localization based on the state of confluence of the endothelial monolayer. The specific function of these PTPs in corneal endothelial cells needs to be examined.
To the best of our knowledge, this is the first study regarding phosphatase function and PTP expression in corneal endothelium. In summary, the results of our studies suggest that phosphatases play an important role in regulating the integrity of cell-cell junctions and cell proliferation in this tissue. We have identified five
phosphatases, PTP-mu, PTP1B, SHP-1, SHP-2, and PTEN, that are expressed in rat corneal endothelium. These are most likely not the only phosphatases expressed in this tissue and their specific function in the endothelium remains to be elucidated. Regardless, this study has opened a fruitful avenue for future investigation and may
lead to a greater understanding of the specific mechanisms underlying regulation of contact-dependent inhibition in corneal endothelium.
ACKNOWLEDGEMENTS
The authors wish to thank Ms. Rebecca Ickes for excellent technical assistance in early phases of this project.
REFERENCES
Aicher, B., Lerch, M.M., Muller, T., Schilling, J., Ullrich, A. 1997. Cellular redistribution of protein tyrosine phosphatase LAR and PTPs by inducible proteolytic processing. J. Cell Biol. 138, 681-696.
Balsamo, J., Arregui, C., Leung, T., Lilien, J. 1998. The nonreceptor protein tyrosine phosphatase PTP1B binds to the cytoplasmic domain of N-cadherin and regulates the cadherin-actin linkage. J. Cell Biol.143, 523-532.
Balsamo, J., Leung, T., Ernst, H., Zanin, M.K., Hoffman, S., Lilien, J. 1996. Regulated binding of PTP1B-like phosphatase to N-cadherin: control of
cadherin-mediated adhesion by dephosphorylation of beta-catenin. J. Cell Biol. 134, 801-813.
Barford, D., Das, A.K., Egloff, M.P. 1998. The structure and mechanism of protein phosphatase: Insights into catalysis and regulation. Annu. Rev. Biophys. Biomol. Struct. 27, 133-164.
Beebe, D.C. and Coats, J.M. 2000. The lens organizes the anterior segment: Specification of neural crest cell differentiation in the avian eye. Dev. Biol. 220, 424-431.
Besson, A., Robbins, S.M., Yong, V.W. 1999. PTEN/MMAC1/TEP1 in signal transduction and tumorigenesis. Eur. J. Biochem. 263,605-611.
Brady-Kalnay, S.M., Flint, A.J., Tonks, N.K. 1993. Homophilic binding of PTP mu, a receptor-type protein tyrosine phosphatase, can mediate cell-cell aggregation. J. Cell Biol. 122,961-972.
Brady-Kalnay, S.M., Mourton, T., Nixon, J.P., Pietz, G.E., Kinch, M., Chen, H., Brackenbury, R., Rimm, D.L., Del Vecchio, R.L., Tonks, N.K. 1998. Dynamic interaction of PTP-mu with multiple cadherins in vivo. J. Cell Biol.141, 287-296.
Brady-Kalnay, S.M., Tonks, N.K. 1995. Protein tyrosine phosphatases as adhesion receptors. Curr. Opin. Cell Biol.7, 650-657.
Burden-Gulley, S.M., Brady-Kalnay, S.M. 1999. PTP-mu regulates N-cadherin-dependent neurite outgrowth. J. Cell Bio. 144,1323-1336.
Chen, K.H., Azar, D., Joyce, N.C. 2001. Transplantation of adult human corneal endothelium ex vivo: a morphologic study. Cornea. 20,731-737.
Chen, K.H.,Harris, D.L., Joyce, N.C. 1999.TGF-beta2 in aqueous humor suppresses S-phase entry in cultured corneal endothelial cells. Invest. Ophthalmol. Vis. Sci. 40,2513-2519.
Erlich, H.A., Gelfand, D., Sninsky, J.J. 1991. Recent advances in the polymerase chain reaction. Science. 252,1643-1651.
Gebbink, M.F., Zondag, G.C., Koningstein, G.M., Feiken, E., Wubbolts, R.W., Moolenaar, W.H. 1995. Cell surface expression of receptor protein tyrosine phosphatase RPTP mu is regulated by cell-cell contact. J. Cell Biol.131,251-260 Gerdes J., Li, L., Schlueter, C., Duchrow, M., Wohlenberg, C., Gerlach, C., Stahmer, I.,
Kloth, S., Brandt, E., Flad H.D. 1991. Immunobiochemical and molecular biologic characterization of the cell proliferation-associated nuclear antigen that is defined by monoclonal antibody Ki-67. Am. J. Pathol. 138,867-873.
Geroski, D.H., Edelhauser, H.F. 1984. Quantitation of Na/K ATPase pump sites in the rabbit corneal endothelium. Invest. Ophthalmol. Vis. Sci. 25,1056-1060.
Gospodarowicz, D., Mescher, A.L., Birdwell, C.R. 1977. Stimulation of
corneal .endothelial cell proliferation in vitro by fibroblast and epidermal growth factors. Exp Eye Res. 25,75-89.
Haj, F.G., Verveer P.J., Squire, A, Neel, B.G., Bastiaens, P.I.H. 2002. Imaging sites of receptor dephosphorylatin by PTP1B on the surface of the endoplasmic reticulum. Science. 295, 1708-1711.
Hellberg CB, Burden-Gulley SM, Pietz GE, Brady-Kalnay SM. 2002. Expression of the receptor protein-tyrosine phosphatase, PTPµ, restores E-cadherin-dependent adhesion in human prostate carcinoma cells. J. Biol. Chem. 277,11165-11173. Ickes, R., Harris, D.L. Joyce, N.C. 2002. “Classical” cadherin expression in corneal
endothelium” [abstract]. 2002 Annual Meeting Abstract and Program Planner accessed at www.arvo.org. Association for Research in Vision and Ophthalmology. Abstract #3190.
Iwamoto, T., Smelser, G.K. 1965. Electron microscopy of the human corneal endothelium with reference to transport mechanisms. Invest. Opthalmol. Vis. Sci. 72,270-284.
Jampel, H.D., Roche, N., Stark, W.J., Roberts A.B. 1990. Transforming growth factor-beta in human aqueous humor. Curr. Eye Res. 9,963-969.
Joyce, N.C. 2003. Proliferative capacity of the corneal endothelium. Prog. Retinal Eye Res. 22:359-389.
Joyce, N.C., Harris, D.L., Mello, D.M. 2002. Mechanisms of mitotic inhibition in corneal endothelium: Contact inhibition and TGF-β2. Invest. Ophthalmol. Vis. Sci. 43,2152-2159.
Joyce, N.C., Harris, D.L., Zieske, J.D. 1998. Mitotic inhibition of corneal endothelium in neonatal rats. Invest. Ophthalmol. Vis. Sci. 39,2572-2583.
Joyce, N.C., Meklir, B., Joyce, S.J., Zieske, J.D. 1996a. Cell cycle protein expression and proliferative status in human corneal cells. Inves.t Ophthalmol. Vis. Sci. 37,645-655.
Joyce, N.C., Meklir, B., Neufeld, A.H. 1990. In vitro pharmacologic separation of corneal endothelial migration and spreading responses. Invest. Ophthalmol. Vis. Sci.31,1816-1826.
Joyce, N.C., Navon, S.E., Roy, S., Zieske, J.D. 1996b. Expression of cell
cycle-associated proteins in human and rabbit corneal endothelium in situ. Invest. Ophthalmol. Vis. Sci 37,1566-1575.
Kamiyama, K., Iguchi, I., Wang, X., Kita, M., Imanishi, J., Yamaguchi, N., Hongo, M., Sotozono, C., Kinoshita, S. 1995. Enhancement of growth of rabbit corneal endothelial cells by PDGF. Cornea 14,187-195.
Kaufman, H.E., Katz, J.I. 1977. Pathology of the corneal endothelium. Invest. Ophthalmol. Vis. Sci. 16,265-268.
Keilhack, H., Tenev, T., Nyakatura, E., Godovac-Zimmermann, J., Nielsen, L., Seedorf, K., Bohmer, F.D. 1998. Phosphotyrosine 1173 mediates binding of the protein-tyrosine phosphatase SHP-1 to the epidermal growth factor receptor and attenuation of receptor signaling. J. Biol. Chem. 273,24839-24846.
Klarlund, J.K. 1985. Transformation of cells by an inhibitor of phosphatases acting on phosphotyrosine in proteins. Cell. 41,707-717.
Kreutziger, G.O. 1976. Lateral membrane morphology and gap junction structure in rabbit corneal endothelium. Exp. Eye Res.23, 285-293.
Kypta, R.M., Su, H., Reichardt, L.F. 1996. Association between a transmembrane protein tyrosine phosphatase and the cadherin-catenin complex. J. Cell Biol. 134,1519-1529.
Li, J., Yen, C., Liaw, D., Podsypanina, K., Bose, S., Wang, S.I., Puc, J., Miliaresis, C., Rodger, L., McCombie, R., Bigner, S.H., Giovanella, B.C., Ittmann, M., Tycko, B., Hibshoosh, H., Wigler, M.H., Parson, R. 1997. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science 275,1943-1947.
Liu, F., Sells, M.A., Chernoff, J. 1998. Protein tyrosine phosphatase 1B negatively regulates integrin signaling. Curr. Biol. 8, 173-176.
Machama T, Dixon JE. 1998. The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate. J. Biol. Chem. 273,13375-13378.
Marrero, M.B., Venema, V.J., Ju, H., Eaton, D.C., Venema, R.C. 1998. Regulation of angiotensin II-induced JAK2 tyrosine phosphorylation: role of SHP-1 and SHP-2. Am. J. Physiol. 275,1216-1223.
Maurice, D.M. 1972. The location of the fluid pump in the cornea. J. Physiol. 221,43-54.
Myers MP, Pass I, Batty IH, et al. 1998. The lipid phosphatase activity of PTEN is critical for its tumor suppressor function. Proc. Natl. Acad. Sci. USA.
95,13513-13518.
Pallen, C.J., Tong, P.H. 1991. Elevation of membrane tyrosine phosphatase activity in density-dependent growth-arrested fibroblasts. Proc. Natl. Acad. Sci. USA.
88,6996-7000.
Paramio, J.M., Navarro, M., Segrelles, C., Gomez-Casero, E, Jorcano, J.L. 1999. PTEN tumour suppressor is linked to the cell cycle control through the
retinoblastoma protein. Oncogene 18,7462-7468.
Pathre, P., Arregui, C., Wampler, T., Kue, I., Leung, T.C., Lilien, J., Balsamo, J. 2001. PTP1B regulates neurite extension mediated by cell-cell and cell-matrix adhesion molecules. J. Neurosci. Res. 63,143-150.
Petroll, W.M., Hsu, J.K., Bean, J., Cavanagh, H.D. and Jester, J.V. 1999. The spatial organization of apical junctional complex-associated proteins in feline and human corneal endothelium. Curr. Eye Res. 18, 10-19.
function; lifting fog or simple shifting? J. Cell Sci. 113,2345-2354.
Qi, J.H., Ito, N., Claesson-Welsh, L. 1999. Tyrosine phosphatase SHP-2 is involved in regulation of platelet-derived growth factor-induced migration. J. Biol. Chem. 274,14455-14463.
Qu, C.K., Yu, W.M., Azzarelli, B., Feng, G.S. 1999. Genetic evidence that SHP-2 tyrosine phosphatase is a signal enhancer of the epidermal growth factor receptor in mammals. Proc. Natl. Acad. Sci. USA. 96,8528-8533.
Ramaswamy, S., Nakamura, N., Vazquez, F., Batt, D.B. Perera, S., Roberts, T.M., Sellers, W.R. 1999. Regulation of G1 progression by the PTEN tumor suppressor protein is linked to inhibition of the phosphatidylinositol 3-kinase/Akt pathway. Proc. Natl. Acad. Sci. USA. 96,2110-2115.
Rhee, J., Lilien, J., Balsamo, J. 2001. Essential tyrosine residues for interaction of the non-receptor protein-tyrosine phosphatase PTP1B with N-cadherin. J. Biol. Chem. 276,6640-6644.
Senoo, T., Joyce, N.C. 2000a. Cell cycle kinetics in corneal endothelium from old and young donors. Invest. Ophtalmol. Vis. Sci. 41,660-667.
Senoo, T., Obara, Y., Joyce, N.C. 2000b. EDTA: a promoter of proliferation in human corneal endothelium. Invest. Ophthalmol. Vis. Sci. 41,2930-2935
Sherrard, E.S. 1976. The corneal endothelium in vivo: its response to mild trauma. Exp. Eye Res. 22,347-357.
Siliciano, J.D., Goodenough, D.A. 1988. Localization of the tight junction protein, ZO-1, is modulated by extracellular calcium and cell–cell contact in Madin–Darby canine kidney epithelial cells. J. Cell Biol. 107,2389-2399.
Sorby, M., Ostman, A. 1996. Protein-tyrosine phosphatase-mediated decrease of epidermal growth factor and platelet-derived growth factor receptor tyrosine phosphorylation in high cell density cultures. J. Biol. Chem. 271,10963-10966.
Starborg, M., Gell, K., Brundell, E., Hoog.C. 1996. The murine Ki-67 cell
proliferation antigen accumulates in the nucleolar and heterochromatic regions of interphase cells and at the periphery of the mitotic chromosomes in a process essential for cell cycle progression. J. Cell Sci. 109, 143-153.
Stern, M.E., Edelhauser, H.F., Pederson, H.J., Staatz, W.D. 1981. Effects of ionophores X537a and A23187 and calcium-free medium on corneal endothelial morphology. Invest. Ophthalmol. Vis. Sci. 20, 497-508.
Suzuki, E., Nagata, D., Yoshizumi, M., Kakoki, M., Goto, A., Omata, M., Hirata, Y. 2000. Reentry into the cell cycle of contact-inhibited vascular endothelial cells by a phosphatase inhibitor. J. Biol. Chem. 275,3637-3644.
Takahashi, K., Suzuki, K. 1996. Density-dependent inhibition of growth involved prevention of EGF receptor activation by E-cadherin-mediated cell-cell adhesion. Exp. Cell Res. 226,214-222.
Tonks, N.K. 2003. PTP1B: From the sidelines to the front lines! FEBS Lett. 546, 140-148.
Tsukita, S., Oishi, K., Akiyama, T., Yamanashi, Y., Yamamoto, T., Tsukita, S. 1991. Specific proto-oncogenic tyrosine kinases of src family are enriched in cell-to-cell adherens junctions where the level of tyrosine phosphorylation is elevated. J. Cell Biol. 113, 867-79.
Van Setten, G.B., Fagerholm, P., Philipson, B., Schultz, G. 1996. Growth factors and their receptors in the anterior chamber. Absence of epidermal growth factor and transforming growth factor alpha in human aqueous humor. Ophthalmic Res. 28,361-364.
Wilson, S.E., Lloyd, S.A., He, Y.G., McCash, C.S. 1993. Extended life of human corneal endothelial cells transfected with the SV40 large T antigen. Invest. Ophthalmol. Vis. Sci.34, 2112-2123.
Wilson, S.E., Weng, J., Blair, S., He, Y.G., Lloyd, S. 1995. Expression of E6/E7 or SV40 large T antigen-coding oncogenes in human corneal endothelial cells indicates regulated high-proliferative capacity. Invest. Ophthalmol. Vis. Sci. 36,32-40.
Xu, G., Arregui, C., Lilien, J., Balsamo, J. 2002. PTP1B modulates the association of beta-catenin with N-cadherin through binding to an adjacent and partially
Young, B.A., Sui, X., Kiser, T.D., Hyun, S.W., Wang, P., Sakarya, S., Angelini, D.J., Schaphorst, K.L., Hasday, J.D., Cross, A.S., Romer, L.H., Passaniti, A., Goldblum, SE. 2003. Protein tyrosine phosphatase activity regulates endothelial cell-cell interactions, the paracellular pathway, and capillary tube stability. Am. J. Physiol. Lung Cell Mol. Physiol. 285, 63-75.
Yu, Z., Su, L., Hoglinger, O., Jaramillo, M.L., Banville, D., Shen, S.H. 1998. SHP-1 associates with both platelet-derived growth-factor receptor and the p85 subunit of phosphatidylinositol 3-kinase. J. Biol. Chem. 273, 3687-3694.
Zhu, C.C., Joyce, N.C. 2004. Proliferative response of corneal endothelial cells from young and older donors. Invest. Ophthalmol. Vis. Sci. 45,1743-1751.