QUERCETIN AFFECTED LEUKEMIA WEHI-3 CELLS IN VIVO 163
Copyright © 2009 John Wiley & Sons, Ltd. Phytother. Res. 24: 163–168 (2010)
Phytother. Res. 24: 163–168 (2010)
Published online 18 May 2009 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/ptr.2841
Quercetin Inhibited Murine Leukemia WEHI-3
Cells In Vivo and Promoted Immune Response
Chun-Shu Yu1, Kuang-Chi Lai2,3, Jai-Sing Yang4, Jo-Hua Chiang5, Chi-Cheng Lu5,Chang-Lin Wu6, Jing-Pin Lin7, Ching-Lung Liao8, Nou-Ying Tang7, W. Gibson Wood9
and Jing-Gung Chung6,10*
1
Center of General Education, China Medical University, Taichung 404, Taiwan
2Department of Surgery, China Medical University Beigang Hospital, Beigang, Yunlin 651, Taiwan 3
School of Medicine, China Medical University, Taichung 404, Taiwan
4Department of Pharmacology, China Medical University, Taichung 404, Taiwan 5
Department of Life Sciences, National Chung Hsing University, Taichung 402, Taiwan
6Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan 7
School of Chinese Medicine, China Medical University, Taichung 404, Taiwan
8Graduate Institute of Chinese Medical Science, China Medical University, Taichung 404, Taiwan 9
Department of Pharmacology, University of Minnesota, School of Medicine, Geriatric Research, Education and Clinical Center, VA Medical Center, Minneapolis, MN 55455, USA
10
Department of Biotechnology, Asia University, Wufeng, Taichung 413, Taiwan
Enhanced flavonoid consumption is closely related with a reduced cancer incidence as shown in epidemiologi-cal studies. Quercetin (3,5,7,3′′′′′,4′′′′′-pentahydroxylflavone) is one of the active components of flavonoids which exist in natural plants, particularly in onions and fruits. It was reported that quercetin induced apoptosis in human cancer cell lines, including human leukemia HL-60 cells, but there is no available information as to its effects on leukemia cells in vivo. The purpose of the present studies was to focus on the in vivo effects of quercetin on leukemia WEHI-3 cells. The effects of quercetin on WEHI-3 cells injected into BALB/c mice were examined. Quercetin decreased the percentage of Mac-3 and CD11b markers, suggesting that the differ-entiation of the precursors of macrophages and T cells was inhibited. There was no effect on CD3 levels but increased CD19 levels. Quercetin decreased the weight of the spleen and liver compared with the olive oil treated animals. Quercetin stimulated macrophage phagocytosis of cells isolated from peritoneum. Quercetin also promoted natural killer cell activity. Based on pathological examination, an effect of quercetin was observed in the spleen of mice previously injected with 3 cells. Apparently, quercetin affects WEHI-3 cells in vivo. Copyright © 2009 John Wiley & Sons, Ltd.
Keywords: quercetin; WEHI-3 cells; BALB/c mice; in vivo; phagocytosis.
Received 1 December 2008 Revised 28 February 2009 * Correspondence to: Dr Jing-Gung Chung, Department of Biological
Science and Technology, China Medical University, No 91, Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C.
E-mail: [email protected]
Contract/grant sponsor: China Medical University, Taichung, Taiwan; contract/grant number: CMU95-330.
INTRODUCTION
In Taiwan about 2.1 per 100 000 people die each year of leukemia and it is the 13th most common malig-nancy based on reports of the ‘People Health Bureau of Taiwan’. The treatment of leukemia is not satisfac-tory and it was reported that an increased consumption of a plant-based diet led to a reduction of colon cancer (Mutoh et al., 2000; Wenzel et al., 2000). Herbal based dietary supplements contain many phytochemicals which might mediate physiological functions related to cancer suppression in vivo. Immune responses are involved in many kinds of leukocytes such as T cells, B cells, natural killer cells and macrophages etc. It is well known that CD3 is the marker for T cells, CD11b is the marker for monocytes or NK cells, CD19 is the marker for B cells and Mac-3 is the marker for macrophages. Macrophages perform phagocytosis then digest antigen
for helper T cells, then these T cells help B cells form plasma cells to release immunoglobulin (antibody) to bind the antigen. Those antigens will be phagocytosed by macrophages.
Quercetin, one of the active components of flavonoids, possesses various biological activities such as antioxidant, antiinflammation, antiatherosclerosis and antitumor prop-erties (Mutoh et al., 2000; Naderi et al., 2003; Wenzel
et al., 2000). One of the antitumor effects of quercetin
is to induce cytotoxic effects such as inhibition of cell proliferation and induction of apoptosis in human cancer cell lines. The molecular mechanisms of quercetin induced cytotoxic effects include cell cycle arrest and induction of caspase-mediated apoptosis in breast (Singhal
et al., 1995), colon (van Erk et al., 2005), leukemia (Lee et al., 2006; Mertens-Talcott et al., 2003; Shen et al.,
2003), lung (Nguyen et al., 2004), liver (Granado-Serrano
et al., 2006) and oral (Ong et al., 2004) cancer cells.
It was reported that quercetin inhibited the invasive potential of melanoma and prostate cancers (Kandaswami
et al., 2005; Zhang et al., 2000; Zhang et al., 2004) and
tube formation in human umbilical vascular endothelial cells (Tan et al., 2003). It was also reported that quercetin inhibited lipopolysaccharide- or 12-O-tetradecanoylphorbol-13-acetate-induced inflammation and matrix metallopro-teinase-9 (MMP-9) expression in mouse macrophages
Copyright © 2009 John Wiley & Sons, Ltd. Phytother. Res. 24: 163–168 (2010)
164 C-S. YU ET AL.
and glioma cells (Chen et al., 2004; Lin et al., 2003). Although various bioactivity studies of quercetin have been carried out the antileukemia capacity of quercetin has not yet been analysed. Therefore, the effects of quercetin on murine leukemia WEHI-3 cells in vivo were investigated.
MATERIALS AND METHODS
Materials and reagents. Quercetin and olive oil were obtained from Sigma Co. (St Louis, MO, USA). RPMI 1640, fetal bovine serum, penicillin–streptomycin and glutamine were obtained from Gibco BRL (Grand Island, NY, USA).
BALB/c mice. Fifty male BALB/c mice 8 weeks of age (approximately 22–28 g) were purchased from the Labo-ratory Animal Center, National Taiwan University, College of Medicine (Taipei, Taiwan).
Murine leukemia WEHI-3 cells. The murine myelomo-nocytic leukemia cell line (WEHI-3) was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan). The cells were plated onto 75 cm2 cell culture flasks and grown in RPMI 1640
medium containing 1% penicillin-streptomycin (100 U/ mL penicillin and 100μg/mL streptomycin), 1% glutamine and 10% fetal bovine serum, at 37 °C under a humidified 5% CO2 atmosphere.
In vivo studies. Fifty BALB/c mice were divided into five groups (10 animals per group) and maintained at 25 °C on a 12 h light/dark cycle. Quercetin (2 and 4 mg/kg) was administered by gauge in isotonic saline (1 mL of saline administered) and control animals received 1% olive oil only. Group I was the control and Group II was treated with olive oil only. Group III was injected i.p. with WEHI-3 cells only. Group IV was injected i.p.with WEHI-3 cells and then treated with quercetin (2 mg/kg) in olive oil. Group V was injected i.p. with WEHI-3 cells and then treated with quercetin (4 mg/kg) in olive oil. Mice were treated daily for 3 weeks before being weighed and killed (Su et al., 2008).
Blood collection and immunofluorescence staining. After 3 weeks, 1 mL of blood was collected from all animals. The blood sample was treated immediately with ammo-nium chloride to lyse the red blood cells and then cen-trifuged at 1500 rpm (1000 × g) at 4 °C for 15 min. Isolated white blood cells were examined for cell surface markers of T cells (CD3), B cells (CD19), monocytes and macrophages (CD11b and Mac-3) (WEHI-3 is a myelomonocytic leukemia cell line) using staining with anti-CD3, CD11b, CD19 and Mac-3 antibodies (BD Pharmingen, San Diego, CA, USA) then were stained with the second fluorescent antibody to determine the cell marker levels by flow cytometry (FACS Calibur™, Becton Dickinson, NJ, USA) as described (Loken et al., 2000).
Liver and spleen tissues. All mice were weighed before blood was drawn. The liver and spleen were isolated and also weighed for each animal (Su et al., 2008).
Phagocytic activity of macrophages. Macrophage phago-cytosis was measured by using the Phagotest kit (Orpegen Pharma, Heidelberg, Germany). The cells were isolated from PBMC and the peritoneum of the control and experimental animals. Isolated cells from each group were individually incubated for 4 h at 37 °C with opsonized fluorescein isothiocyanate (FITC)-labeled
E. coli (20μL) as per the manufacturer’s instruction.
An ice-cold quenching solution (100μL) was added to stop the reaction. After the completion of phagocytosis, the monocytes/macrophages were fixed, and cell cycle analysis for viable cells (dead cells were stained) was performed according to the manufacturer’s instructions. The cell preparations were analysed by a flow cytometer (FACSCalibur, Becton Dickinson). The fluorescence data were collected on 10 000 cells and analysed using Cellquest software (Stachowska et al., 2007).
Histopathology. Spleen samples from each group were fixed in 4% formaldehyde and embedded in paraffin. Sections of 5 mm were stained with hematoxylin and eosin according to standard procedures (Yang et al., 2006).
NK activity. Approximately 1 × 105 leukocytes from
mice spleen in 1 mL of medium were cultured in each well of a 96-well. About 2.5 × 107 cells of YAC-1 (NK
target cells) in 15 mL tubes with serum-free RPMI-1640 medium and the PKH-67/Dil.C buffer was added to the cells then mixed thoroughly for 2 min at 25 °C, then 2 mL PBS was added for 1 min. Then 4 mL RPMI-1640 was added for 10 min incubation, then centrifuged at 1200 rpm and 25 °C. About 2.5 × 106 cells of YAC-1
were placed onto 96-well plates for 100μL before the addition of the leukocytes to the well for 12 h and determination of the NK cell activation by flow cytometry as described previously (Yang et al., 2006). Statistical analysis. Data were expressed as mean ± SD and differences between the control and experimental groups were analysed by Student’s t-test. A value of
p < 0.05 was used as the level of significance.
RESULTS
Injection of WEHI-3 cells induces leukemia tumors in mice
Representative whole body images of the control and quercetin treated BALB/c mice after injection with WEHI-3 cells for 3 weeks and the presence of leukemia tumors are shown in Fig. 1A. The spleen and liver size was decreased compared with the WEHI-3 cells only treated mice.
The effects of quercetin on the weights of spleen and liver from BALB/c mice after injection with WEHI-3 cells
Spleen and liver tissues were isolated from animals and were weighed individually. Representative results are presented in Fig. 1B and C. The results indicate that quercetin affected the weights of the spleen and liver.
Copyright © 2009 John Wiley & Sons, Ltd. Phytother. Res. 24: 163–168 (2010) Figure 1. Representative pictures from dissected BALB/c mice after injection with WEHI-3 cells and/or treated with quercetin for 3
weeks. BALB/c mice were injected with WEHI-3 cells (1 × 105cells/100μL) in PBS for 3 weeks and/or co-treated with quercetin, blood
was collected and animals (A) were killed for examinations of spleen (B) and liver (C) tissues and were individually photographed and weighed (n = 10).
After exposure to 2 and 4 mg/kg quercetin, the spleen weights in the control and quercetin treated groups were decreased by 2% and 19%, respectively, the liver weights were decreased by 14% and 28%, respectively. These effects were dose-dependent.
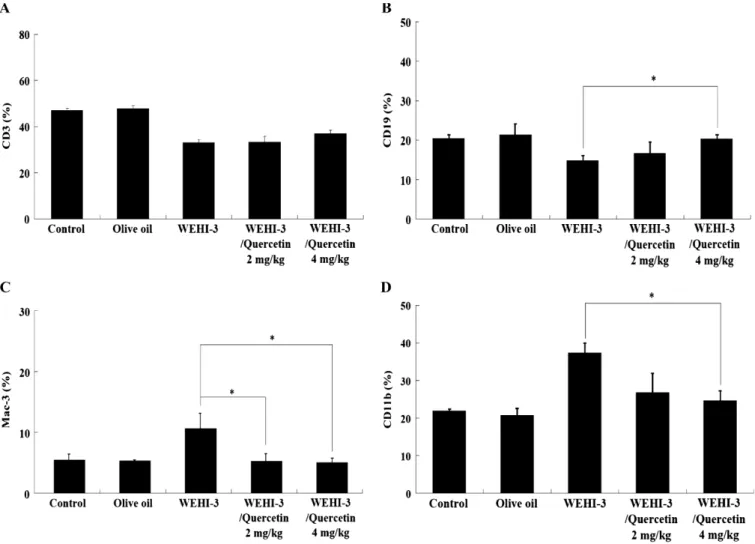
Effects of quercetin on whole blood cell surface markers of BALB/c mice after injection with WEHI-3 cells
The percentage changes in cell markers of white blood cells from BALB/c mice after treatment with quercetin in olive oil or olive oil treatment are shown in Fig. 2A, B, C and D. Quercetin reduced the levels of Mac-3 (Fig. 2C) and CD11b (Fig. 2D) but increased the levels of CD19 (Fig. 2B) and had little, if any, effect on CD3 (Fig. 2A).
Effects of quercetin on macrophage activity of BALB/c mice after injection with WEHI-3 cells The percentage changes of macrophages with phagocy-tosed green fluorescent particles from PBMC and peri-toneum of the control and quercetin treated groups are presented in Fig. 3. Quercetin did not induce signifi-cant differences in macrophage activity in cells isolated from PBMC (Fig. 3A). However, quercetin stimulated macrophage activity in cells from the peritoneum (Fig. 3B). These effects were dose-dependent manners.
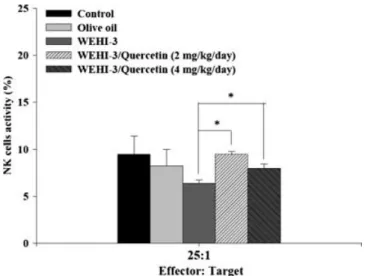
Effects of quercetin on activity of natural killer cells from BALB/c mice after injection with WEHI-3 cells The YAC-1 target cells were killed by NK cells isolated from the spleen of mice after being treated with quercetin in a target cell ratio of 25:1, both doses of quercetin showed a significant difference between the control and tested agents treatment in a target cell ratio of 25:1 (Fig. 4).
Morphological effects of quercetin on spleen of BALB/c mice after injection with WEHI-3 cells There was a marked expansion in the red pulp, but the white pulp showed little change in spleen tissue (Fig. 5). Neoplastic cells contained large irregular nuclei accom-panied by clumped chromatin and prominent nucleoli, abundant clear and light eosinophilic cytoplasm. Often mitotic figures were also noted.
DISCUSSION
Quercetin is a component of flavonoids which has been reported to have anticancer activity such as: (1) apoptosis in different human cancer cell lines (Granado-Serrano
et al., 2006; Lee et al., 2006; Mutoh et al., 2000; Nguyen et al., 2004; Singhal et al., 1995; Vijayababu et al., 2006;
Copyright © 2009 John Wiley & Sons, Ltd. Phytother. Res. 24: 163–168 (2010)
166 C-S. YU ET AL.
Figure 2. The effects of quercetin on cell markers of white blood cells from BALB/c mice. BALB/c mice were injected with WEHI-3
cells (1 × 105cells/100μL) in PBS for 3 weeks and treated without or with quercetin for 3 weeks. Blood was collected from individual
animals and analysed for cell markers (A) CD3, (B) CD19, (C) Mac-3 and (D) CD11b by flow cytometry as described in Materials and Methods. Each point is mean ± SD. * p < 0.05 (n = 10).
growth factor-induced invasiveness (Huang et al., 1999; Lee et al., 2004) and; (3) inhibition of metastasis of melanoma and prostate cancer (Piantelli et al., 2006; Vijayababu et al., 2006). It was also reported that quercetin induced apoptosis in human leukemia HL-60 cells in vitro (Hibasami et al., 2005). However, the role of quercetin on leukemia cells in vivo has not been determined. Primary experiments also demonstrated that quercetin induced cytotoxicity in human leukemia HL-60 cells and murine leukemia WEHI-3 cells there-fore raising the possibility that quercetin could affect mouse WEHI-3 leukemia cells in vivo. The present study showed that quercetin effectively suppressed leukemia WEHI-3 cells which had been injected into BALB/c mice in vivo. The study used WEHI-3 cells for i.p. in-jection into BALB/c mice because leukemic animals have been used to test for anticancer agents and also it is a low cost, easy method that takes a short time to develop leukemia in vivo.
The data indicated that quercetin significantly decreased the average size and weight of the liver and spleen of BALB/c mice injected with leukemia cells. Quercetin also decreased the percentage of Mac-3 and CD11b
cells, promoted CD19 cells but did not alter CD3 mar-kers in the blood. The stimulating effects of quercetin on the CD19 marker indicated it may promote B cell numbers. Quercetin increased macrophage phagocytosis from isolated PBMC and peritoneum and promoted natural killer cell activity. This is in agreement with regard to agents that promote immune responses are designed to increase the phagocytosis of macrophages and the activities of natural killer cells. The results demonstrated that quercetin inhibited leukemia-related spleen growth. A notable characteristic of the leukemia model used in this study is the elevation of peripheral monocytes and granulocytes with immature morphology, as well as enlarged and infiltrated spleens compared with the normal counterpart (He and Na, 2001). Based on these observations it is indicated that quercetin also promoted immune responses in BALB/c mice in vivo.
Acknowledgement
This work was supported by Grant CMU95-330 from China Medical University, Taichung, Taiwan.
Copyright © 2009 John Wiley & Sons, Ltd. Phytother. Res. 24: 163–168 (2010) Figure 3. The effects of quercetin on macrophage phagocytosis
from BALB/c mice. BALB/c mice were injected with WEHI-3 cells (1 × 105cells/100μL) in PBS for 3 weeks and treated without or
with quercetin for 3 weeks. Cells were collected from (A) PBMC and (B) peritoneum of animals and analysed for macrophage phagocytosis by flow cytometry as described in Materials and Methods. Each point is mean ± SD. * p < 0.05 (n = 10).
Figure 4. The effects of quercetin on NK cell activity from BALB/
c mice. BALB/c mice were injected with WEHI-3 cells (1 × 105cells/100μL) in PBS for 3 weeks and treated without or with
quercetin for 3 weeks. Blood was collected from peritoneum of animal and analysed for NK cell activity by flow cytometry as described in Materials and Methods. Each point is mean ± SD. * p < 0.05 (n = 10).
Figure 5. The effects of quercetin on the histopathology of
the spleen from BALB/c mice. BALB/c mice were injected with WEHI-3 cells after treatment with quercetin. Spleens from each animal of each group were excised for histopathological examination as described in Materials and Methods.
REFERENCES
Chen TJ, Shen SC, Lin HY, Chien LL, Chen YC. 2004. Lipopolysaccharide enhancement of 12-o-tetradecanoyl-phorbol 13-acetate-mediated transformation in rat glioma C6, accompanied by induction of inducible nitric oxide synthase. Toxicol Lett 147: 1–13.
Granado-Serrano AB, Martin MA, Bravo L, Goya L, Ramos S. 2006. Quercetin induces apoptosis via caspase activation, regulation of Bcl-2, and inhibition of PI-3-kinase/Akt and ERK pathways in a human hepatoma cell line (HepG2). J Nutr 136: 2715–2721.
He Q, Na X. 2001. The effects and mechanisms of a novel 2-aminosteroid on murine WEHI-3B leukemia cells in vitro and in vivo. Leuk Res 25: 455–461.
Hibasami H, Mitani A, Katsuzaki H, Imai K, Yoshioka K, Komiya T. 2005. Isolation of five types of flavonol from seabuckthorn (Hippophae rhamnoides) and induction of apoptosis by some of the flavonols in human promyelotic leukemia HL-60 cells. Int J Mol Med 15: 805–809.
Huang YT, Hwang JJ, Lee PP et al. 1999. Effects of luteolin and quercetin, inhibitors of tyrosine kinase, on cell growth and metastasis-associated properties in A431 cells
overexpress-ing epidermal growth factor receptor. Br J Pharmacol 128: 999–1010.
Kandaswami C, Lee LT, Lee PP et al. 2005. The antitumor activities of flavonoids. In Vivo 19: 895–909.
Lee LT, Huang YT, Hwang JJ et al. 2004. Transinactivation of the epidermal growth factor receptor tyrosine kinase and focal adhesion kinase phosphorylation by dietary flavonoids: effect on invasive potential of human carcinoma cells. Biochem Pharmacol 67: 2103–2114.
Lee TJ, Kim OH, Kim YH et al. 2006. Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Cancer Lett 240: 234–242.
Lin HY, Juan SH, Shen SC, Hsu FL, Chen YC. 2003. Inhibition of lipopolysaccharide-induced nitric oxide production by flavonoids in RAW264.7 macrophages involves heme oxygenase-1. Biochem Pharmacol 66: 1821–1832.
Loken MR, Green CI, Wells DA. 2000. Immunofluorescence of surface markers. In Flow Cytometry: A Practical Approach, Ormerod MG (ed.). Oxford University Press: Oxford, 61– 82.
Copyright © 2009 John Wiley & Sons, Ltd. Phytother. Res. 24: 163–168 (2010)
168 C-S. YU ET AL.
concentrations of quercetin and ellagic acid synergistically influence proliferation, cytotoxicity and apoptosis in MOLT-4 human leukemia cells. J Nutr 133: 2669–267MOLT-4.
Mutoh M, Takahashi M, Fukuda K et al. 2000. Suppression by flavonoids of cyclooxygenase-2 promoter-dependent tran-scriptional activity in colon cancer cells: structure-activity relationship. Jpn J Cancer Res 91: 686–691.
Naderi GA, Asgary S, Sarraf-Zadegan N, Shirvany H. 2003. Anti-oxidant effect of flavonoids on the susceptibility of LDL oxidation. Mol Cell Biochem 246: 193–196.
Nguyen TT, Tran E, Nguyen TH, Do PT, Huynh TH, Huynh H. 2004. The role of activated MEK-ERK pathway in quercetin-induced growth inhibition and apoptosis in A549 lung cancer cells. Carcinogenesis 25: 647–659.
Ong CS, Tran E, Nguyen TT et al. 2004. Quercetin-induced growth inhibition and cell death in nasopharyngeal carcinoma cells are associated with increase in Bad and hypophos-phorylated retinoblastoma expressions. Oncol Rep 11: 727– 733.
Piantelli M, Rossi C, Iezzi M et al. 2006. Flavonoids inhibit melanoma lung metastasis by impairing tumor cells endothelium interactions. J Cell Physiol 207: 23–29. Shen SC, Chen YC, Hsu FL, Lee WR. 2003. Differential
apoptosis-inducing effect of quercetin and its glycosides in human promyeloleukemic HL-60 cells by alternative activation of the caspase 3 cascade. J Cell Biochem 89: 1044–1055. Singhal RL, Yeh YA, Praja N, Olah E, Sledge GW, Jr, Weber G.
1995. Quercetin down-regulates signal transduction in human breast carcinoma cells. Biochem Biophys Res Commun 208: 425–431.
Stachowska E, Baskiewicz-Masiuk M, Dziedziejko VB et al. 2007. Conjugated linoleic acids can change phagocytosis of human
monocytes/macrophages by reduction in Cox-2 expression. Lipids 42: 707–716.
Su CC, Yang JS, Lin SY et al. 2008. Curcumin inhibits WEHI-3 leukemia cells in BALB/c mice in vivo. In Vivo 22: 63–68. Tan WF, Lin LP, Li MH et al. 2003. Quercetin, a dietary-derived
flavonoid, possesses antiangiogenic potential. Eur J Pharmacol
459: 255–262.
van Erk MJ, Roepman P, van der Lende TR et al. 2005. Inte-grated assessment by multiple gene expression analysis of quercetin bioactivity on anticancer-related mechanisms in colon cancer cells in vitro. Eur J Nutr 44: 143–156. Vijayababu MR, Arunkumar A, Kanagaraj P, Venkataraman P,
Krishnamoorthy G, Arunakaran J. 2006. Quercetin down-regulates matrix metalloproteinases 2 and 9 proteins expression in prostate cancer cells (PC-3). Mol Cell Biochem
287: 109–116.
Wenzel U, Kuntz S, Brendel MD, Daniel H. 2000. Dietary flavone is a potent apoptosis inducer in human colon carcinoma cells. Cancer Res 60: 3823–3831.
Yang JS, Kok LF, Lin YH et al. 2006. Diallyl disulfide inhibits WEHI-3 leukemia cells in vivo. Anticancer Res 26: 219–225. Yu FS, Yu CS, Chan JK et al. 2006. The effects of emodin on the expression of cytokines and functions of leukocytes from Sprague-Dawley rats. In Vivo 20: 147–151.
Zhang X, Xu Q, Saiki I. 2000. Quercetin inhibits the invasion and mobility of murine melanoma B16-BL6 cells through inducing apoptosis via decreasing Bcl-2 expression. Clin Exp Metastasis 18: 415–421.
Zhang XM, Huang SP, Xu Q. 2004. Quercetin inhibits the invasion of murine melanoma B16-BL6 cells by decreasing pro-MMP-9 via the PKC pathway. Cancer Chemother Pharmacol 53: 82–88.