http://ict.sagepub.com/
Integrative Cancer Therapies
http://ict.sagepub.com/content/9/3/298
The online version of this article can be found at: DOI: 10.1177/1534735410378661
2010 9: 298 originally published online 16 August 2010 Integr Cancer Ther
Kuan-Hung Lu, Yuh-Fang Chang, Pen-Hui Yin, Ting-Ting Chen, Yu-Ling Ho, Yuan-Shiun Chang and Chin-Wen Chi
HL-60 Human Leukemia Cells
In Vitro and In Vivo Apoptosis-Inducing Antileukemic Effects of Mucuna macrocarpa Stem Extract on
Published by:
http://www.sagepublications.com
can be found at: Integrative Cancer Therapies
Additional services and information for
http://ict.sagepub.com/cgi/alerts Email Alerts: http://ict.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://ict.sagepub.com/content/9/3/298.refs.html Citations:
© The Author(s) 2010
Reprints and permission: http://www. sagepub.com/journalsPermissions.nav DOI: 10.1177/1534735410378661 http://ict.sagepub.com
In Vitro and In Vivo Apoptosis-
Inducing Antileukemic Effects
of Mucuna macrocarpa Stem Extract
on HL-60 Human Leukemia Cells
Kuan-Hung Lu, MS
1, Yuh-Fang Chang, BS
1, Pen-Hui Yin, PhD
2,
Ting-Ting Chen, MS
3, Yu-Ling Ho, PhD
4, Yuan-Shiun Chang, PhD
1,
and Chin-Wen Chi, PhD
2,5Abstract
Mucuna macrocarpa Wallich (Leguminosae) is believed to hold blood circulation activating effects, and has been used as a folk remedy in Southeast Asia for the treatment of various hematologic and circulatory-related ailments. The objective of this study was to investigate whether crude methanolic extract of M macrocarpa (CMEMM) possessed antileukemic effects on HL-60, human leukemia cells. CMEMM was prepared from dried stems of this plant, and its apoptosis-inducing effects were investigated using HL-60 cells in vitro and in vivo. With treatment of 25 to 75 mg/mL CMEMM, the in vitro antiproliferative effect on HL-60 cells increased in a dose- and time-dependent manner during the 72-hour treatment period. The concentration of CMEMM that exhibited a 50% growth inhibition (IC50) for 72-hour exposure was 36.4 mg/mL. Apoptosis triggered by CMEMM in HL-60 cells was confirmed by the following observations: (a) characteristic apoptotic nuclear fragmentation, (b) dose-dependent accumulation of sub-G1 phase in cell cycle analyses, (c) increased percentages of annexin V-positive apoptotic cells, and (d) dose-dependent elevation of active caspase-3. Furthermore, an in vivo tumor growth suppression effect by CMEMM (500 mg/kg/d intraperitoneally) was observed in mouse xenografts. The results suggest that CMEMM exerts antileukemic effects via an apoptotic pathway in HL-60 cells, and could be a candidate for developing antileukemic agents in the future.
Keywords
Mucuna macrocarpa, antileukemia, apoptosis, HL-60, xenograft, caspase-3
Introduction
Mucuna macrocarpa Wallich belongs to the Leguminosae family. It is a large woody climber found in forests at alti-tudes of 100 to 1500 meters, and is distributed throughout Taiwan and Southeast Asia.1 Dried stems of this plant have been used in folk medicine to activate blood circulation for various hematologic and circulatory-related ailments, such as anemia, hemoptysis, dysmenorrheal, pain in waist and knees, numbness of hands and feet.2 Additionally, crude drug of this medicinal plant has been used as a remedy for the treatment of diabetes mellitus.3 In phytochemical investigations, amino acids, lipids, and triterpenoids have been isolated.4,5 More-over, it has also been demonstrated to possess diuretic and spasmolytic activities in rat models;6 however, there has been no studies on the anticancer effect of this plant.
Leukemia, a cancer of the blood or bone marrow, is clini-cally and pathologiclini-cally subdivided into 2 different forms,
myeloid and lymphoid. Acute lymphoblastic leukemia (ALL) more frequently occurs in children, whereas the risk of acute myeloid leukemia (AML) is higher in adults. Acute promyelocytic leukemia (APL) is a distinct subtype of AML characterized by a specific genetic alteration, affecting the retinoic acid receptor-a, and leading to a blockage in the dif-ferentiation of granulocytic cells.7 APL is unique from other forms of AML in its striking response to anthracyclines and
1China Medical University, Taichung, Taiwan 2Taipei Veterans General Hospital, Taipei, Taiwan 3National Taiwan Sport University, Taoyuan, Taiwan 4Huangkuang University, Taichung, Taiwan 5National Yang-Ming University, Taipei, Taiwan
Corresponding Author:
Chin-Wen Chi, Department of Medical Research and Education, Taipei Veterans General Hospital, No. 201, Sec. 2, Shih-Pai Road, Taipei 112, Taiwan
differentiating agents such as all-trans retinoic acid (ATRA) or arsenic trioxide; however, the relapse rate has been reported to be about 30% to 40%.8 Despite significant progress in the treatment of these diseases, therapy is still unsuccessful in many patients. Prognosis is particularly poor in adult AML;9 moreover, ATRA is known to have more fre-quent adverse effects in pediatric patients.10 Therefore, developing new therapeutic drugs for refractory or recurrent leukemia is a worthwhile task.
Apoptosis, a programmed cell death, plays an important role in the development of various diseases, including cancer.11 In recent years, interest has been focused on the manipulation of apoptotic processes in the treatment and prevention of cancers.12 Much effort has been directed toward the search for herbs that influence apoptosis as alter-native cancer therapies because of their low toxicity and costs.13,14 In addition, recent reports have demonstrated that extracts from herbal medicines exert antileukemic effects by inducing apoptosis in human leukemia cells.15,16
In the present study, the in vitro apoptosis-inducing abil-ity of crude methanolic extract of M macrocarpa (CMEMM) was investigated on human leukemia cell line, HL-60. Anti-proliferative activity was examined by trypan blue exclusion assay. CMEMM-induced apoptosis was determined by typ-ical apoptotic morphologic changes, cell cycle analysis, annexin V binding assay, and caspase-3 activation analysis. Furthermore, the in vivo antitumor effect was examined using human leukemia xenograft model.
Materials and Methods
Reagents
RPMI-1640 medium, fetal bovine serum (FBS), gentamy-cin, trypan blue dye solution, and phosphate-buffered saline (PBS; pH 7.4) were purchased from Gibco (Grand Island, NY). Dimethyl sulfoxide (DMSO) and ethidium bromide were purchased from Sigma (St. Louis, MO). Acetic acid and methanol were purchased from Merck (Darmstadt, Germany). Cycle TEST PLUS DNA Reagent Kit was pur-chased from Becton Dickinson (San Jose, CA). ANNEX100F Kit (annexin V: FITC assay kit) was purchased from AbD Serotec (Kidlington, UK). Active Caspase-3 FITC Mab Apoptosis Kit was purchased from BD Pharmingen (San Diego, CA).
Plant Material and Extraction
Mucuna macrocarpa Wallich (Leguminosae) was collected in Nantou County, Taiwan in July 2006. The material was identified by Professor Yuan-Shiun Chang in the Graduate Institute of Chinese Pharmaceutical Sciences, China Medi-cal University; a voucher specimen (MM 950729) has been
deposited in the same institute. Crude methanolic extract of the stems of M macrocarpa (CMEMM, yield = 13.9% of dry weight) was prepared as described below. The stems were cut into small pieces, air-dried for 1 week, and extracted with methanol for 48 hours at room temperature 3 times. All extracts were mixed, and the mixture was fil-tered and concentrated using a rotary evaporator until dry.
Cell Culture and CMEMM Treatment
Human promyelocytic leukemia HL-60 cell line was obtained from the American Type Culture Collection (ATCC, Rockville, MD). HL-60 cells were cultured in RPMI-1640 medium containing 10% FBS and 0.01 mg/mL gentamycin and incubated in a humidified atmosphere of 5% CO2 at 37°C. CMEMM was dissolved in DMSO and diluted in culture medium to obtain final concentrations of 25, 50, and 75 mg/mL. Cells (1 × 105 cells/mL) were seeded into 60-mm dishes and exposed to vehicle or various con-centrations of CMEMM for 24 to 72 hours. The concentration of DMSO (0.1%) did not affect the cell viability. Cells were harvested and counted by hemocytometer at 24, 48, and 72 hours after treatment with CMEMM and stored for fur-ther examinations.
Cell Proliferation Assay
Cell growth was determined by trypan blue exclusion assay. Cells were collected at indicated times following CMEMM exposure. After centrifugation, cells were resuspended in culture medium and stained with 0.4% trypan blue solu-tion. Viable cells were counted using a hemocytometer. The percentages of growth inhibition were calculated by comparing the cell numbers of CMEMM group with that of the control.
Cytological Examination
Ethidium bromide fluorescence staining method was used to observe apoptotic morphology of individual cells. Briefly, cells (1 × 106 cells/mL) were washed twice with PBS and collected on microscope slides by cytospin (Shandon Cyto-spin Cytocentrifuge). This was followed by fixation with 25% (v/v) acetic acid in methanol for 10 minutes and stain-ing with ethidium bromide solution (20 mg/mL) at room temperature. Photographs were taken under an inverted fluorescence microscopy (Olympus IX-70). Fragmented nuclei were suggestive of apoptosis.
Cell Cycle Analysis
DNA staining was carried out using Cycle TEST PLUS DNA Reagent Kit. Briefly, cells (1 × 106 cells/mL) were
washed and stained for DNA content according to the kit pro-tocol. Fluorescence intensity of propidium iodide (PI) was determined using a FACScan flow cytometer and analyzed by CellQuest software (Becton Dickinson, San Jose, CA).
Annexin V-FITC/PI Analysis
Annexin V-FITC/PI staining method was used to identify and quantify apoptotic cells. Cells (2 × 105 to 5 × 105 cells/mL) were treated according to the manufacturer’s instruction of ANNEX100F Kit. FITC/PI fluorescence intensity was measured using a FACScan flow cytometer to differentiate between viable (annexin V-negative and PI-negative), early apoptotic (annexin V-positive, PI-negative), and necrotic cells (annexin V-positive and PI-positive). The extent of apoptosis was quantified according to the percentage of annexin V-positive cells.
Active Caspase-3 Analysis
The amount of active caspase-3, a marker for cells undergo-ing apoptosis, was measured usundergo-ing Active Caspase-3 FITC Mab Apoptosis Kit. The FITC-conjugated monoclonal rabbit anti-active caspase-3 antibody specifically recognizes the active form of caspase-3 in human or mouse cells. Briefly, cells (1 × 106 cells/0.5 mL) were permeabilized, fixed, and stained for active caspase-3 according to the Active Caspase-3 FITC Staining Protocol. The cells were then analyzed by flow cytometry to determine FITC fluorescence intensity.
Human Leukemia Xenograft Model
In vivo testing of antileukemic effect of CMEMM was per-formed in xenograft models and carried out with ethics committee approval (No. 97-148) from Taipei Veterans General Hospital. A total of 22 male athymic nude mice (BALB/c nude, with body weight of 20 to 24 grams and aged 5 weeks) were obtained from the National Laboratory Animal Center (Taipei, Taiwan) and kept in an autoclaved cage with polyester fiber filters to avoid contact with patho-gens. The mice were housed under sterilized conditions with a constant temperature of 25 ± 2°C and 12-hour light:12-hour dark cycles. All animal diet and water were autoclaved before feeding to the mice ad libitum. The mice were acclimatized to the housing condition at least 1 week before the experi-ment. Both sides of lateral thighs of the nude mice were injected subcutaneously with 1 × 106 HL-60 cells. When palpable tumors (4 to 6 mm in diameter) arose 3 weeks after injection, the mice were randomly divided into 3 groups (7 to 8 mice per group). The experimental groups were treated with CMEMM extracts (100 or 500 mg/kg/d in auto-claved water), and the control was treated with autoauto-claved water by daily intraperitoneal (i.p.) injection. Tumor size
and body weight were monitored daily throughout the exper-iment. Tumor volumes were measured by a digital caliper and calculated according to the following formula: tumor volume (mm3) = 0.4 × L × W2, where L and W were the major and minor dimensions of the tumor, respectively.17 After 21 days of treatment, the mice were euthanized and tumors were removed. The final body and tumor weights were recorded for the calculation of percentage ratio final tumor-to-body weight. Tumors, livers, kidneys, and spleens were collected, fixed, embedded, and stained with hema-toxylin and eosin (H&E) for pathological analyses.
Phytochemical Analysis
Phytochemical components of CMEMM were analyzed by chromatographic methods. Briefly, CMEMM was suspended in distilled water and partitioned successively with different poloarities of organic solvents such as n-hexane, chloroform, ethyl acetate, and n-butanol. Chemical components were isolated from these partitioned fractions using column chro-matography. By means of infrared (Nicolet Impact 400 FT-IR spectrophotometer), EI-MS (VG Platform II Mass Spec-trometer), and 1H NMR and 13C NMR (Bruker DPX-200 FT-NMR), the structures of chemical components were iden-tified on the basis of spectral evidence. The major isolated components from CMEMM were further determined by the application of HPLC (Waters 2695 separation module). HPLC chromatogram was carried out on XBridge RP-18 end-capped column (5 mm pore size, 250 × 4.6 mm, inner diameter) by an isocratic elution of 2% aqueous acetic acid and acetoni-trile (63:37, v/v) solvent. The peaks were recorded with a photodiode array detector (Waters 996 PDA detector) at 254 nm, and the solvent flow rate was kept at 0.8 mL/min.
Statistical Analysis
To assess statistical significant differences between the control and CMEMM-treated groups, the data were ana-lyzed by one-way analysis of variance (ANOVA) followed by Student–Newman–Keuls test using the SPSS for Win-dows 10.0 version software. Differences were considered significant when P < .05.
Results
Antiproliferative Effect of CMEMM
on HL-60 cells
As illustrated in Figure 1, CMEMM reduced the prolifera-tion of HL-60 cells in a dose- and time-dependent manner. Significant growth inhibitions (P < .05) were observed for cells treated with 50 and 75 mg/mL of CMEMM. Similar response profiles were observed in the 48- and 72-hour
treatment groups. Growth inhibitions were most obvious after 72 hours of treatment; the percentage of growth inhi-bition increased from 22.7% to 81.0% as the dosage of CMEMM increased from 25 to 75 mg/mL. Time response data demonstrated that 50% growth inhibition (IC50) was observed at 36.4 mg/mL for 72 hours.
Nuclear Morphological
Changes Induced by CMEMM
After 72 hours incubation with CMEMM, nuclear morpho-logical alterations in HL-60 cells were determined by ethidium bromide staining. As shown in Figure 2, 25 mg/mL CMEMM-treated cells had similar nuclear morphology as that of the control. However, the nuclei of 50 and 75 mg/mL CMEMM-treated cells presented with condensed, fragmented chromatin and formation of apoptotic bodies, which were in clear con-trast to the spherical and intact nuclei of the control.
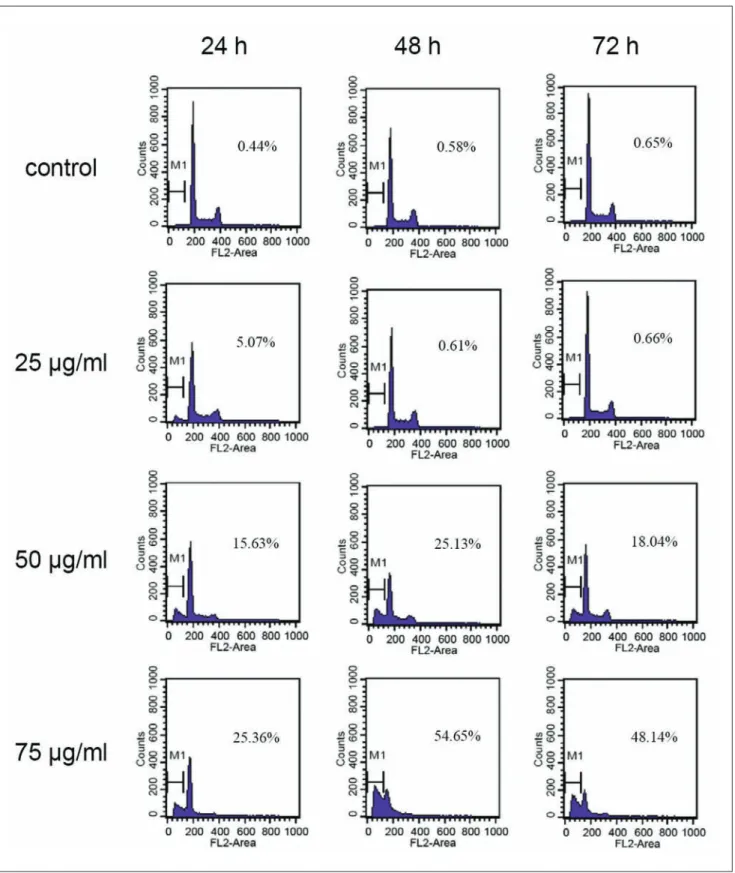
Cell Cycle Progression in CMEMM-treated cells
After exposure to 25, 50, or 75 mg/mL of CMEMM for 24 to 72 hours, the cell cycle of HL-60 cells were analyzed using flow cytometry. As shown in Figure 3, CMEMM-treated HL-60 cells at sub-G1 phase (or hypodiploid apoptotic cells) increased in a dose-dependent manner. After 72 hours of treatment, the percentage of 75 mg/mL CMEMM-treatedFigure 1. A ntiproliferative effects of crude methanolic extract
of Mucuna macrocarpa (CMEMM) on HL-60 cells
Cells (1 × 105cells/mL) were seeded onto 60-mm dishes and exposed
to dosages of CMEMM ranging from 25 to 75 mg/mL for up to 72 hours. Control cells were treated with 0.1% dimethyl sulfoxide (DMSO) in medium. The percentages of growth inhibition were measured by trypan blue exclusion assay and calculated by comparing the cell numbers with that of the controls. Each value represents the mean ± standard error (mean ± SE) of 6 replicate cultures from 3 independent experiments. *P < .05 and **P < .01 indicate significant difference from the control value.
Figure 2. Nuclear morphological changes induced by crude
methanolic extract of Mucuna macrocarpa (CMEMM)
Nuclear morphology of HL-60 cells treated with (A) 0.1% dimethyl sulfoxide (DMSO; control), (B) 25 mg/mL CMEMM, (C) 50 mg/mL CMEMM, and (D) 75 mg/mL CMEMM for 72 hours. The nuclei were stained with ethidium bromide. Arrows indicate apoptotic bodies of nuclear fragmentation. Magnification ×200.
Figure 3. Cell cycle progression in crude methanolic extract of Mucuna macrocarpa (CMEMM)-treated cells
Cell cycle analyses of HL-60 cells treated with 0.1% dimethyl sulfoxide (DMSO; control), 25 mg/mL CMEMM, 50 mg/mL CMEMM, and 75 mg/mL CMEMM for 24 to 72 hours. After treatment with CMEMM, cells were collected and stained with propidium iodide, and analyzed for DNA content by flow cytometry. The percentages of sub-G1 or hypodiploid cells were analyzed by CellQuest software. The representative cell cycle progressions in CMEMM-treated or vehicle control cells were from 1 of 3 independent experiments.
cells at sub-G1 phase increased from 25.4% to 54.7%. As for the control, no significant changes were observed (0.4% to 0.7%).
Annexin V Binding in CMEMM-Treated Cells
Annexin V staining can identify apoptosis at an early stage. To further confirm and quantify apoptosis in HL-60 cells triggered by CMEMM, cells were stained with annexin
V-FITC/PI, and then analyzed by flow cytometry. After treatment with 25, 50, or 75 mg/mL CMEMM for 24 to 72 hours, the percentage of annexin V-positive apoptotic cells increased in a dose- and time-dependent manner (Figure 4). During the 72-hour treatment period, the percentage of early apoptotic cells in the 75 mg/mL CMEMM-treatment group increased from 8.5% to 64.8%. As for the control group, no significant changes were observed (1.3% to 2.5%). Further-more, the percentage of necrotic cells increased from
Figure 4. Annexin V-FITC/PI analyses of crude methanolic extract of Mucuna macrocarpa (CMEMM)-treated cells
HL-60 cells were treated with 0.1% dimethyl sulfoxide (DMSO; control), 25 mg/mL CMEMM, 50 mg/mL CMEMM, and 75 mg/mL CMEMM for 24 to 72 hours. Quantitative percentages of apoptotic and necrotic cells of CMEMM-treated cells were measured using flow cytometry. Data represent the result from 1 of 3 independent experiments.
0.6% to 1.8% and from 7.1% to 23.4% for the control group and 75 mg/mL CMEMM treatment group, respectively.
Involvement of Active Caspase-3
in CMEMM-Treated Cells
Because caspase-3 is a key member in the caspase family of cysteine proteases that cleave protein substrates and give rise to characteristic apoptotic morphology, the activation of cas-pase-3 has been used as an index of apoptosis.18 To investigate whether CMEMM-induced apoptosis in HL-60 cells involved the activation of caspase-3, cells were analyzed using flow cytometry with active caspase-3-specific antibody. As shown in Figure 5, caspase-3 activity in HL-60 increased after treat-ment with 25, 50, or 75 mg/mL of CMEMM for 24 hours in a dose-dependent manner. The percentage of cells containing active caspase-3 increased from 2.8% as in the control to 42.5% in cells treated with 75 mg/mL of CMEMM.
In Vivo Antiproliferation Effect of CMEMM
on HL-60 in Mouse Xenograft Models
To further examine whether CMEMM can effectively sup-press growth in vivo, HL-60 cells were transplanted into athymic nude mice. After tumor formation, the mice were treated either with CMEMM (100 or 500 mg/kg/d, i.p.) or with vehicle PBS as control. CMEMM at dosage of 500 mg/ kg/d significantly inhibited the growth of subcutaneously transplanted HL-60 cells in athymic mice after 21 days of treatment, as supported by both the tumor volume (Figure 6A) and the final tumor weight measurements (Figure 6B). The relative tumor volume (ratio of tumor volume on each test day to that of day 1) of 500 mg/kg/d CMEMM-treated animals were significantly (P < .05) reduced when compared with the control from days 5 to 21. At the end of the study, the relative tumor volume of 100 mg/kg/d CMEMM-treated and control animals had average relative tumor volumes of 16.8 and 16.6, respectively. In contrast, the 500 mg/kg/d CMEMM-treated animals had an average relative tumor volume of 9.4. Moreover, the final tumor-to-body weight ratio for the 500 mg/kg/d CMEMM-treated animals was sig-nificantly (P < .05) reduced when compared with the controls. No obvious acute toxicity of CMEMM was observed during the treatment period, and there was no significant difference between the body weight of the treated and control animals. Also, no tissue damage was observed in the livers, kidneys, and spleens following examination of the tissue slices stained with H&E (data not shown).
Chemical Components and HPLC
Chromatogram of CMEMM
As shown in Figure 7A, nine chemical components were isolated from CMEMM. Based on the NMR, IR, and MS
spectroscopic data, these compounds were identified by comparison of spectroscopic data with literature reports. These included 1 lipid (tetracosanoic acid19), 1 sterol Figure 5. Activation of caspase-3 in crude methanolic extract of
Mucuna macrocarpa (CMEMM)-treated cells
Active caspase-3 analyses were performed on HL-60 cells treated with (A) 0.1% dimethyl sulfoxide (DMSO; control), (B) 25 mg/mL CMEMM, (C) 50 mg/mL CMEMM, and (D) 75 mg/mL CMEMM for 24 hours. After treatment with CMEMM, cells were permeabilized, fixed and stained, and analyzed for active caspase-3 by flow cytometry. Data represent the result from 1 of 3 independent experiments.
(mixture of b-sitosterol and stigmasterol20), 2 carbohy-drates (sucrose21 and d-pinitol22),1 triterpenoid (friedelin23), and 4 isoflavones (medicarpin24,calycosin25, afrormosin,26 and genistein27). As shown in Figures 7B and 7C, calycosin, afrormosin, and genistein were used as marker compo-nents for the standardization of flavonoid ingredients of CMEMM by high-performance liquid chromatography (HPLC) chromatograms.
Discussion and Conclusions
In the theory of traditional Chinese medicine, blood-activating and stasis-resolving herbs have been used for the treatment of various hematologic and circulatory related ailments since ancient times. Since these medicinal herbs can relieve symptoms related to leukemia, they have been employed as clinical dialectical therapy for patients with myeloproliferative disorders by the Chinese in recent years.28 Additionally, recent reports have demonstrated that antileu-kemic effects of these herbs were exerted by induction of apoptosis in human leukemia cells.29,30 In the present study, we have for the first time examined the antileuke-mic activity of M macrocarpa, seeking scientific support for its traditional use.
The current study demonstrated that CMEMM was effec-tive against the proliferation of human promyelocytic leukemia cells, HL-60 (Figure 1). However, 2 lymphoid leukemia cell lines, Jurkat T and Molt-3 cells, showed sim-ilar responses but were less sensitive to CMEMM-induced
Figure 6. In vivo antiproliferative effect of crude methanolic extract of Mucuna macrocarpa (CMEMM) on human leukemia HL-60
xenografts
When the tumors were palpable (4 to 6 mm), BALB/c nude mice were treated either with vehicle control or with CMEMM by daily intraperitoneal injection. The animals (7 to 8 mice per group) were treated with 100 mg/kg/day CMEMM (CME100), 500 mg/kg/day CMEMM (CME500), or with phosphate-buffered saline (PBS) as vehicle control for 21 days. A, For relative tumor volume, results were expressed as mean ± standard error (mean phosphate-buffered saline (PBS) as vehicle control for 21 days. A, For relative tumor volume, results were expressed as mean ± standard error (mean ± SE) calculated from the ratio of tumor volume on each test day to that of day 1. B, For percentage ratio of final tumor-to-body weight, results were expressed as the mean ± SE. *P < .05 and **P < .01 indicate significant difference from the control value.
growth inhibition in comparison to HL-60 in our prelimi-nary study (data not shown). Similarly, Chang et al 31 had also reported differences in sensitivity between promyelo-cytic and lymphoid leukemia cells when exposed to the same agents in cell proliferation assays. APL cells (HL-60) were more sensitive to drug-induced growth inhibition than ALL cells (Molt-3). An important finding is that apoptosis in HL-60 cells triggered by CMEMM was confirmed not only by the characteristic apoptotic morphology (Figure 2) and accumulation at sub-G1 phase (Figure 3) but also by an increase of annexin V-FITC/PI staining (Figure 4). More-over, active caspase-3 elevation was found to be a key mediator for CMEMM-induced apoptosis (Figure 5). These results together suggest that CMEMM exerts antileukemic effects via an apoptotic pathway in HL-60 cells.
Furthermore, the present in vivo results confirmed the growth-inhibitory effect of CMEMM on human leukemia xenografts in athymic nude mice. Throughout the 21 days of treatment by i.p. injection, CMEMM at 500 mg/kg/d was efficacious in tumor suppression, and the ratio of final tumor-to-body weight was significantly reduced in comparison to that of the control (Figure 6). However, in our pilot study, treating mice with 500 mg/kg/d of CMEMM in their daily gavage did not result in observable tumor suppressing effect on HL-60 xenografts. This implied that the extent of bio-availability in CMEMM-treated animals through gavage was not as high as i.p. injection probably because of pharmacoki-netic behavior of the different administration routes. To improve bioavailability of CMEMM, pharmacological
studies such as dosage form design and formulation develop-ment will be worthwhile tasks in the future.
Using column chromatography and spectroscopy, 9 com-ponents from the stem of M macrocarpa were isolated and identified as 1 lipid, 1 sterol, 2 carbohydrates, 1 triterpe-noid, and 4 isoflavones (Figure 7A). In previous studies of the bioactive isoflavones isolated in this experiment, medi-carpin and genistein had been reported to exert antileukemic effects by inducing apoptosis in HL-60 cells;32,33 calycosin had been reported to exert antiproliferative effects in U937 lymphoma cells;34 and afrormosin had been reported to exhibit inhibitory effects on TPA-induced tumor promo-tion in mouse.35 Since medicarpin was absent from the HPLC chromatogram because of its relatively low content, the other bioactive isoflavones were identified in CMEMM
with pure marker components (Figure 7B). Although not shown in the present study, the HPLC quantitative analy-sis showed the content of genistein was 0.001% in air-dry stems. It has been reported recently that IC50 value of genis-tein for 48-hour exposure was 18.2 mM in HL-60 cells.36 It was reasonable to assume that genistein was not the main component in CMEMM that induced growth inhibition on HL-60 cells because of the low content of genistein in air-dried stems of this plant. This implied that the antileukemic effect of M macrocarpa may have resulted from the interac-tion of different flavonoids and other constituents.
In conclusion, we presented evidence that treating human leukemia HL-60 cells with CMEMM led to the induction of apoptosis. Therefore, our results suggested that CMEMM could be a candidate for developing antileukemic
Figure 7. Phytochemical analyses of crude methanolic extract of Mucuna macrocarpa (CMEMM)
A, Chemical components were isolated from CMEMM using chromatographic methods: tetracosanoic acid (H-1), friedelin (H-2), mixture of
b-sitosterol and stigmasterol (C-1), medicarpin (C-2), calycosin (E-1), afrormosin (E-2), genistein (E-3), sucrose (B-1), and d-pinitol (B-2). B and C,
high-performance liquid chromatography (HPLC) chromatograms show a standard mixture and CMEMM: (1) calycosin; (2) afrormosin; (3) genistein. (For HPLC chromatographic conditions, please see the Materials and Methods section.)
agents. For future therapeutic application, further studies of CMEMM in comedications with synthetic drugs are now in progress in our laboratory.
Acknowledgments
We thank Dr Anna F.-Y. Li for pathological analyses. Authors’ Note
Dr Yuan-Shiun Chang and Dr Chin-Wen Chi contributed equally to this work. Correspondence may also be addressed to Dr Yuan-Shiun Chang at Graduate Institute of Chinese Pharmaceutical Sciences, China Medical University, No. 91, Hsueh-Shih Road, Taichung 404, Taiwan. E-mail address: [email protected] Declaration of Conflicting Interests
The author(s) declared no conflicts of interest with respect to the authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article:
This study was supported in part by grants from the National Science Council (NSC 97-2320-B-075-006-MY3; NSC 94-2320- B-039-023; NSC 92-2320-B-039-041), Taipei Veterans General Hospital (V97E2-011; V96E2-007), Department of Health (DOH99-TD-C-111-007), and China Medical University (CMC91-CPS-02), Taiwan.
References
1. Editorial Committee of the Flora of Taiwan. Flora of Taiwan. 2nd ed. Vol. 3. Taipei, Taiwan: Tah Jinn; 1993.
2. Editorial Board of Zhong Hua Ben Cao, State Administration of Traditional Chinese Medicine. Zhong Hua Ben Cao (China Herbal). Vol. 4. Shanghai, China: Shanghai Scientific Tech-nology Press; 1999.
3. Lin CC. Crude drugs used for the treatment of diabetes mel-litus in Taiwan. Am J Chin Med. 1992;20:269-279.
4. Chen Y, Zhen HS, Xu XJ, Pan Y, Tang J. Determination oflevodopa in Maodou (Mucuna pruriens) and Lidou (M. mac-rocarpa) by TLC scanning [in Chinese]. Zhong Cao Yao. 1993; 24:294-295.
5. Hu WY, Luo SD, Cai JX. Studies on the chemical components of Daguoyoumateng (Mucuna macrocarpa) [in Chinese]. Zhong Cao Yao. 1994;25:59-60.
6. Bhakuni DS, Goel AK, Jain S, Mehrotra BN, Patnaik GK, Prakash V. Screening of Indian plants for biological activity: Part XIII. Indian J Exp Biol. 1988;26:883-904.
7. Lo-Coco F, Ammatuna E, Montesinos P, Sanz MA. Acute pro-myelocytic leukemia: recent advances in diagnosis and man-agement. Semin Oncol. 2008;35:401-409.
8. Burry LD, Seki JT. CNS relapses of acute promyelocytic leu-kemia after all-trans retinoic acid. Ann Pharmacother. 2002; 36:1900-1906.
9. Ansari M, Krajinovic M. Pharmacogenomics of acute leuke-mia. Pharmacogenomics. 2007;8:817-834.
10. Lin CH, Hung GY, Chang CY, Chien JC. Subdural hemor-rhage in a child with acute promyelocytic leukemia presenting as subtle headache. J Chin Med Assoc. 2005;68:437-440. 11. Thompson CB. Apoptosis in the pathogenesis and treatment
of disease. Science. 1995;267:1456-1462.
12. Fleischer A, Ghadiri A, Dessauge F, et al. Modulating apop-tosis as a target for effective therapy. Mol Immunol. 2006; 43:1065-1079.
13. Guruvayoorappan C, Kuttan G. Apoptotic effect of Biophy-tum sensitivum on B16F-10 cells and its regulatory effects on nitric oxide and cytokine production on tumor-associated macrophages. Integr Cancer Ther. 2007;6:373-380.
14. Kang SA, Park HJ, Kim MJ, Lee SY, Han SW, Leem KH. Citri Reticulatae Viride Pericarpium extract induced apopto-sis in SNU-C4, human colon cancer cells. J Ethnopharmacol. 2005;97:231-235.
15. Kumagai T, Muller CI, Desmond JC, Imai Y, Heber D, Koeffler HP. Scutellaria baicalensis, a herbal medicine: anti-proliferative and apoptotic activity against acute lympho-cytic leukemia, lymphoma and myeloma cell lines. Leuk Res. 2007;31:523-530.
16. Jesse P, Mottke G, Eberle J, Seifert G, Henze G, Prokop A. Apoptosis-inducing activity of Helleborus niger in ALL and AML. Pediatr Blood Cancer. 2009;52:464-469.
17. Attia MA, Weiss DW. Immunology of spontaneous mam-mary carcinomas in mice. V. Acquired tumor resistance and enhancement in strain A mice infected with mammary tumor virus. Cancer Res. 1966;26:1787-1800.
18. Kohler C, Orrenius S, Zhivotovsky B. Evaluation of caspase activity in apoptotic cells. J Immunol Methods. 2002;265:97-110. 19. Lin YT, Kuo YH, Chang BH. Studies on the extractive con-stituents of the bark of Libocedrus formosana Florin. II. J Chin Chem Soc. 1975;22:331-334.
20. Kao KC, Ho YL, Ho LK, Chang YS. 2-Benzoxazolinone, 2-hydroxy-1,4-benzoxazin-3-one form the root of Strobilan-thes cusia [in Chinese]. J Chin Med. 2001;12:41-49. 21. Park NY, Baek NI, Cha J, Lee SB, Auhe JH, Park CS.
Pro-duction of a new sucrose derivative by transglycosylation of recombinant Sulfolobus shibatae b-glycosidase. Carbohydr Res. 2005;340:1089-1096.
22. Breimaier E, Voelter W. Carbon-13 NMR Spectroscopy: High-Resolution Methods and Applications in Organic Chem-istry and BiochemChem-istry. 3rd completely rev. ed. New York, NY: VCH; 1987.
23. Akihisa T, Yamamoto K, Tamura T, et al. Triterpenoid ketones from Lingnania chungii McClure: arborinone, friedelin and glutinone. Chem Pharm Bull. 1992;40:789-791.
24. Herath HMTB, Dassanayake RS, Priyadarshani AMA, de Silva S, Wannigama GP, Jamie J. Isoflavonoids and a ptero-carpan from Gliricidia sepium. Phytochemistry. 1998;47: 117-119.
25. Kamnaing P, Fanso Free SNY, Nkengfack AE, Folefoc G, Fomum ZT. An isoflavan-quinone and a flavonol from Mil-lettia laurentii. Phytochemistry. 1999;51:829-832.
26. Caballero P, Smith CM, Fronczek FR, Fischer NH. Isofla-vones from an insect-resistant variety of soybean and the molecular structure of afrormosin. J Nat Prod. 1986;49: 1126-1129.
27. Kanakubo A, Koga K, Isobe M, et al. First finding of daidzein 7-O-phosphate and genistein 7-O-phosphate that are hydro-lyzed by sulfatase. Tetrahedron. 2001;57:8801-8805. 28. Wu SJ, Yan MX. Preliminary analysis about blood-activating
and stasis-eliminating in clinical dialectic of chronic myelo-cytic leukemia [in Chinese]. Study J Tradit Chin Med. 2001; 19:343-344.
29. Dai XZ. Mechanisim of Yiqi Yangyin and Huoxue Huayu on apoptosis in treating minimal residual leukemia in rats [in Chinese]. J Tianjin Univ Tradit Chin Med. 2004;23:177-178. 30. Zheng J, Liu Q, Wang ZR, et al. Blood-activating and stasis-eliminating compound induce K562 cells apoptosis [in Chinese]. Di 4 Jun Yi Da Xue Xue Bao. 2003;24:427-429.
31. Chang YF, Chi CW, Chern YT, Wang JJ. Effects of 1, 6-bis[4-(4-amino-3-hydroxyphenoxy)phenyl]diamantane
(DPD), a reactive oxygen species and apoptosis inducing agent, on human leukemia cells in vitro and in vivo. Toxicol Appl Pharmacol. 2005;202:1-12.
32. Militao GC, Dantas IN, Pessoa C, et al. Induction of apopto-sis by pterocarpans from Platymiscium floribundum in HL-60 human leukemia cells. Life Sci. 2006;78:2409-2417.
33. Traganos F, Ardelt B, Halko N, Bruno S, Darzynkiewicz Z. Effects of genistein on the growth and cell cycle progression of normal human lymphocytes and human leukemic MOLT-4 and HL-60 cells. Cancer Res. 1992;52:6200-6208.
34. Matsuda H, Yoshida K, Miyagawa K, et al. Rotenoids and flavonoids with anti-invasion of HT1080, anti-prolif-eration of U937, and differentiation-inducing activity in HL-60 from Erycibe expansa. Bioorg Med Chem. 2007; 15:1539-1546.
35. Konoshima T, Kokumai M, Kozuka M, Tokuda H, Nishino H, Iwashima A. Anti-tumor-promoting activities of afromosin and soyasaponin I isolated from Wistaria brachybotrys. J Nat Prod. 1992;55:1776-1778.
36. Raynal NJ, Momparler L, Charbonneau M, Momparler RL. Antileukemic activity of genistein, a major isoflavone present in soy products. J Nat Prod. 2008;71:3-7.