1)行政院農業委員會林業試驗所森林利用組,10066台北市南海路53號 Division of Forest Utilization, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
2)國 立 台 灣 大 學 生 命 科 學 系 ,10617台北市羅斯福路四段1號 Department of Life Science, National Taiwan University, 1 Roosevelt Rd., Sec. 4, Taipei 10617, Taiwan.
3)通訊作者 Corresponding author, e-mail:sschen@tfri.gov.tw
2006年1月送審 2006年3月通過 Received January 2006, Accepted March 2006.
紅檜造林木樹幹與枝條之生長應變探討
陳欣欣
1,3)黃玲瓏
2)李金梅
1)黃彥三
1)摘 要
本研究係利用應變規以刻溝法測定紅檜(Chamaecyparis formosensis Matsum.)造林木根曲樹幹之生 長應變,探討樹高及圓周位置對解放表面生長應變之影響,同時亦測定樹幹內部殘留生長應變,木質 部細胞壁S2層之微纖維傾斜角(microfibril angle, MFA)與生長應變之關係亦被探討。試驗結果顯示生長
應變與樹高間無明顯相關性,根曲樹幹下側有很大的壓縮生長應力,抗壓材之MFA遠大於正常材者。 樹枝承受來自自重之彎曲應力,此一應力與生長應力重疊。樹枝上下側之生長應變較樹幹者為 大。枝條下側之生長應變有隨反彈應變之增大而增大之趨勢,且前者大於後者。吾人認為樹枝下側抗 壓材所產生之壓縮生長應力係受到由自重所引發之重力彎曲應力之影響。因此,抗壓材所引起之高壓 縮應力係與重力刺激有關。樹枝形態受樹枝自重所引起之彎距與生長應力所引起之彎距二者間之交互 作用的影響。樹皮之生長應變測定結果顯示不論樹幹或樹枝均為收縮生長應變,顯示有引張生長應力 之存在。 關鍵詞:抗壓材、微纖維傾斜角、生長應變、反彈應變、內部殘留應變。 陳欣欣、黃玲瓏、李金梅、黃彥三。2006。紅檜造林木樹幹與枝條之生長應變探討。台灣林業科學 21(2):263-72。 Research paper
Growth Strains in Trunks and Branches of
Planted Chamaecyparis formosensis Matsum.
Shin-Shin Chen,
1,3)Ling-Long Kuo-Huang,
2)Chin-Mei Lee,
1)Yan-San Huang
1)【
Summary】
The distribution of growth strain in basal sweeping trunks of Chamaecyparis formosensis
Matsum. was measured by the kerf method using strain gauges. Effects of both tree height and
pe-ripheral positions on the trunk on the released surface strains were examined. The residual internal
growth strains were also measured. The microfibril angles (MFAs) of the S
2layer of cell wall were
measured using the iodine deposition method to examine their influences on growth strain. The
results showed that there was no obvious relationship between tree height and growth strain. We
found large compressive stresses on the lower side of sweeping trunks that differed greatly from
the tensile stresses in normal erect trunks. Regarding the residual internal stresses within erect
trunks, contract strain or tensile stress was present toward the outside, whereas extension strain or
compressive stress was induced toward the inside of the trunk. The MFA of compression wood was
larger than that of normal wood.
Branches suffered bending stresses due to their own weight. This stress was then superposed
on the growth stress. It was found that growth strains on the upper side or lower side of branches
were larger than those in the stems. It was shown that the growth strain increased with an increase
in the spring-back strain on the lower side of branches, and the former was obviously larger than
the latter. This suggests that the generation of growth stresses on the lower side of branches
con-taining highly developed compression wood is affected by the gravitational bending stress due to
their own weight. It was concluded that the branch form is affected by the interaction between the
bending moment due to its own weight and that due to growth stresses.
Key words: compression wood, microfibril angle, growth strain, residual internal stress, spring back
strain.
Chen SS, Kuo-Huang LL, Lee CM, Huang YS. 2006. Growth strains in trunks and branches of
planted Chamaecyparis formosensis matsum. Taiwan J For Sci 21(2):263-72.
緒 言
樹 幹 於 連 年 生 長 之 同 時 累 積 生 長 應 力 (growth stress),此一現象與人工材料於加工 過 程 中 所 產 生 的 殘 留 應 力 頗 為 類 似 。 樹 幹 外 周 之 形 成 層 在 進 行 細 胞 分 裂 之 同 時 , 對 內 產 生木質部,對外產生韌皮部,而於木質部形成 之過程中累積生長應力(Boyd 1972, Yamamoto et al. 1991, Guitard et al. 1999),樹木於肥大 生長時樹幹表面會發生異向性應力分佈,此一 應力經連年累積,樹幹內部為達成力學平衡, 而在樹幹內縱向、徑向及弦向產生特定形式之 殘留應力分佈(Archer and Bynes 1974, Kubler 1987, Fournier et al. 1990),此一應力累積可 說是樹木為適應環境之壓力所產生之不可避免 的生理反應(Niklas 1992, Mattheck and Kubler 1995)。傾斜樹幹或枝條之特定部位常發生較大 之生長應力,企圖使樹幹恢復直立生長或使枝 條朝上方生長,而不致於傾倒或折斷。例如, 針 葉 樹 在 傾 斜 樹 幹 或 樹 枝 之 下 側 產 生 抗 壓 材 (compression wood),闊葉樹則在上側產生抗 張材(tension wood),即使直立通直的樹幹,也 必然在樹幹內部產生生長應力。雖然是直立的 樹木,因環境因素的影響,邊材部有發生壓縮 破壞的危險性,故實際上,不論針、闊葉樹, 均 在 樹 幹 周 邊 部 產 生 纖 維 方 向 的 引 張 應 力 , 藉以削弱可能發生的壓縮應力而維持樹幹的安 全。因此可以認為樹木是藉生長應力來巧妙地 適應環境而維持樹體之生理需要(Mattheck and Kubler 1995)。然而,人類在砍伐樹木並做木 材加工利用之際,此生長應力卻造成利用上的 困擾,在立木狀態下恆續保持平衡狀態之樹幹 內部之殘留應力分佈,在各加工過程中被解放 出來,伐倒及截斷時可能發生心裂,製材時會 發生反翹,造成利用率下降。因此從木材利用 的立場,有必要就生長應力之發生機制加以探 究,並尋求解除生長應力之方法(Okuyama and Sasaki 1979, Okuyama et al. 1990)。樹木在地球生態體系中扮演重要的環境保 護角色後,再度被以木材利用之形式作各種不 同的有效利用,為人類的生活提供極為有用的 資材。唯由於人口不斷增加,林產工業迅速發 展,使森林資源日漸枯竭,全球性的環保意識 因而日益增強,為因應大環境的變化,林產研
究的方向亦應有所調整,俾與林學研究相互配 合,相輔相成,相得益彰。
紅檜(Chamaecyparis formosensis Matsum.) 分布於本省中央山脈海拔1,000~2,800 m之地 區 , 與 台 灣 扁 柏 共 稱 檜 木 , 為 台 灣 最 貴 重 之 針 葉 樹 , 其 材 質 極 為 均 勻 優 良 , 主 供 建 築 、 傢具、器物、彫刻、造船及化粧合板之用,極 具經濟價值。惟自實施保育政策以來,紅檜天 然林已遭禁伐,並積極進行造林工作。目前棲 蘭山林區已有生育狀況極佳的天然更新林,溪 頭林區亦有生長良好的造林木,惟此等生長快 速之紅檜次生林,其樹幹生長應力發生情況尚 不明瞭,實有加以探究之必要。惟以往生長應 力的研究大多以樹幹或原木為探討對象,僅少 數著墨於枝條之問題(Yoshida et al. 1992a, b, Ohsako et al. 1968)。本計劃以棲蘭山紅檜次生 林為對象,探討直立及根曲樹幹之表面生長應 變(growth strain)及內部殘留生長應變。此外並 就樹枝生長應變與反彈應變之關係加以探究。 為 了 解 木 材 組 織 細 微 構 造 與 生 長 應 力 之 關 連 性,並就樹幹表面木質部細胞壁S2層之微纖維
傾斜角(microfibril angle, MFA)加以測定,探討 兩者間之相互關係。
材 料 與 方 法
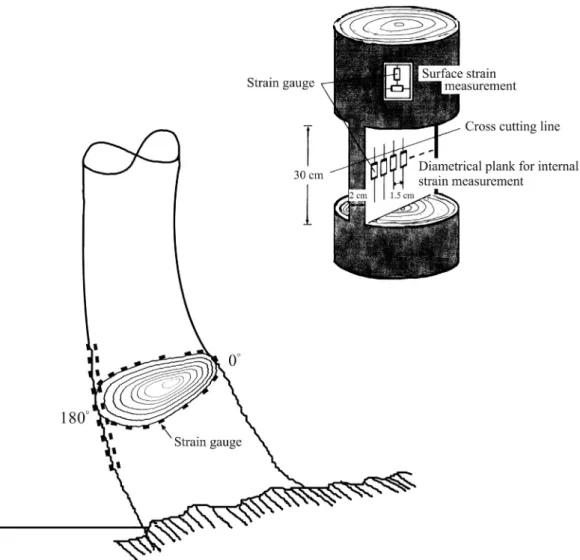
於 森 林 保 育 處 棲 蘭 山 事 業 區 紅 檜 次 生 林 選 取30~35年生立木六株(其中四株為根曲樹 幹)作為供試木,並加以伐採。分別編號為A、 B、C、D、E及F,其胸徑分別為17、27、23、 22、34.5及40 cm,樹高則分別為15、17、16、 16、18及18.5 m。A、B、C三棵樹在樹高1 m 以下略有根曲,1 m以上則通直,因屬造林地經 過修枝作業,A號樹,枝下高約為7~8 m,B、 C號樹,枝下高約為9~10 m。表面生長應變之 測定係就纖維方向與圓周方向同時進行。測定 時先在選定位置剝去樹幹表面之樹皮,然後將 十字型應變規沿木質部之縱向與橫向利用瞬間 膠予以黏貼。生長應變之測定採用附有40頻道 掃描器(USB-11A)之手提式數字顯示應變測定 儀(Model UCAM-A, Kyowa, Japan)。首先將所有貼妥之應變規予以歸零,然後以手鋸於應變 規上下距離約5 mm處鋸切深度約10~15 mm之 橫向切溝,以解除縱向生長應變,然後以鏈鋸 在應變規左右兩側鋸切縱向溝槽,以解除橫向 應變,隨即讀取應變測定儀之顯示數值,此一 刻溝法(Kerf method)之解放應變測定工作便告 完成(Sasaki et al. 1978, Huang et al. 2002)。為 瞭解樹高對生長應變之影響,先將伐倒之樹木 截斷成2.0~2.5 m長之原木,然後於各段原木之 近中央處測定縱向與橫向表面生長應變。至於 樹幹縱斷面上之殘留應變分佈測定,係就E號 樹在樹高5.3 m之直立部分,將原木中央加以鋸 切,留取通過樹心之徑面版(厚約2 cm),並保 留上下兩端完整,然後在近板中央位置每隔1.5 cm沿縱向貼一片應變規,於歸零後,以手鋸於 應變規上方5 mm處予以橫斷,再沿應變規之左 右間隔加以鋸解分隔,隨即讀取儀器之掃描數 值,測定工作便告完成。傾斜樹幹圓周方向生 長應變之分佈測定,係就D號樹之根曲部分,以 上側為0°,下側為180°,順時針方向每隔22.5° ~45°,測定縱向與橫向表面生長應變。因為抗 壓材係發生在傾斜樹幹下側,故沿下側作密集 的生長應變測定,藉以獲取更多資料(Fig. 1)。 樹枝生長應變之測定係就枝條上距主幹不 同距離之部位,先剝去樹皮,然後在上側及下 側分別貼上縱向應變規,於歸零後,將枝條鋸 下,去除側枝及樹葉,隨即讀取因枝條解除自 重後所產生的反彈應變,此後於應變規前後約5 mm處鋸斷枝條,便可測得生長應變。 木質部細胞壁S2層微纖維傾斜角之測定係 採用碘素沈澱法,利用由各樹幹表面生長應變 測定部位所取下之試材,切取20 µm弦面顯微 切片,於顯微鏡下就春材與秋材分別測定30個 管胞之MFA,求取平均值,以便與各該部位所 測得之生長應變互作比較。此外並觀察正常材 與抗壓材之橫切面與徑切面之顯微構造,抗壓 材之存在可由S2層之加厚,圓形細胞壁橫斷面 及其細胞間隙之存在,以及管胞內壁之螺旋狀 開裂(helical fissures)加以確認(Yamamoto et al. 1991)。此外並就樹幹、樹枝之樹皮部分,先剝 去外樹皮,然後將應變規貼在內樹皮上,同樣
以切割法切至木質部,測定解放應變。
結 果 與 討 論
一、樹高對表面生長應變之影響 Figure 2顯示樹高與A、B、C樹號之生長 應變間並無明顯關係。A號於樹高4 m以下部位 之縱向表面生長應變值平均在-340~-350 µε間, 並無太大變化,顯示引張生長應力之存在,惟 於6.2 m樹高部位則因輕微抗壓材之存在,使 生 長 應 變 呈 正 負 互 見 。B號樹於樹高6.2 m以Fig. 1. Measurements of the trunk and of the peripheral positions of a basal sweeping
trunk.
Fig. 2. Effects of tree height on released
surface growth strain (trees A, B, and C).
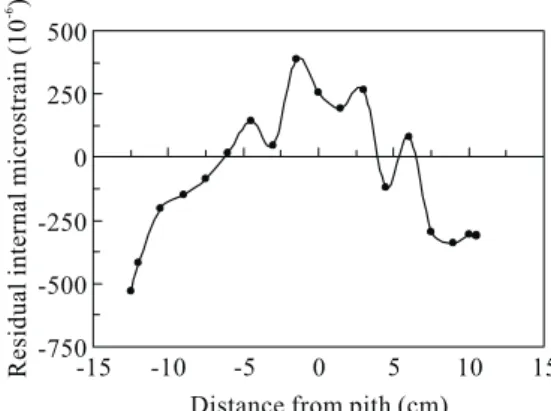
下部位,生長應變值平均在-185~-215 µε間, 亦無太大變化,惟在樹高8.7 m部位,因輕微 抗壓材之存在而呈現平均值+45 µε之較小伸張 應變。C號樹於樹高8.7 m以下部位,生長應變 平均值在-195~-485 µε間呈較大的起伏,且未 發現抗壓材之存在。Huang et al. (1998)就柳 杉直立正常木所作的試驗,顯示生長應力與樹 高間並無明顯關連。另Huang et al. (2003)就 台灣杉所作之測試,顯示不論修枝與否,縱向 與橫向表面生長應變與樹高間無明顯關係。而 Sasaki et al. (1978)就日本柳杉所作之測定,亦 顯示生長應力與樹高無明顯關連。Yao (1979) 選擇易於發生端裂(end split)之胡桃木(Carya ovata)、櫟木(Quercus nigra)與光腊樹(Fraxinus americana),探討生長應力之樹高變異性,顯 示平均在樹高7.7 m處呈現最大應力,平均應力 為97 kgf cm-2,並認定此一應力值足以造成製材 上之問題及原木端裂。Chafe (1981)亦就5株39 年生光腊樹作類似研究,得知彈性係數,比重 及生長應力均隨樹高之增高而增加,且因應力 之增加而使原木易於發生端裂,此點與現場所 見一致。顯然生育地對樹高與生長應力之關係 之效應尚待進一步之探討。 二、樹幹內部之殘留應變分佈 Figure 3顯示E號樹於樹高5.3 m處之樹幹內 部縱向殘留應變分佈。靠樹幹外周為收縮應變 或引張應力,樹幹內側則為伸張應變或壓縮應 力。靠近樹幹外周之最大殘留應變為-529 με, 靠近中心則為+388 με,以上之殘留應變分佈係 表面生長應變累積於樹幹內部之結果。表面生 長應變係每年產生在樹幹之最外側,而樹幹內 部則產生新的應變分佈以因應此一表面生長應 變。換言之,新的應變重疊在原有的應變分佈 上,形成應變的重新分佈,隨著樹幹的生長, 以上過程連年重複進行,因而形成有規則的應 變分佈(Okuyama et al. 1986)。 三、根曲樹幹之生長應變圓周分佈 F i g u r e 4 顯 示 根 曲 樹 幹 ( 樹 號 D ) 之 下 側 (180°)具有頗大的縱向表面生長應變,其範圍在 +1036~+1884 με間。正值表示解放應變為伸張 應變,顯示有較大的壓縮生長應力存在。180° 左右兩側(191°與169°)之生長應變呈現正、負互 見,而其餘各角度則均呈現負值,顯示根曲樹 幹之側面及上側均為引張生長應力,因正常直 立樹幹之外周均呈現引張生長應力(Huang et al. 1998),根曲樹幹下側之壓縮生長應力應來自抗 壓材之存在。Boyd (1980)亦在Pinus radiata D. Don之傾斜樹幹下側測得較大的伸張解放應變 (+1800 με)。抗壓材所產生的壓縮生長應力對傾 斜樹幹之恢復直立具有一定之力學功能,與闊 葉樹相對應,針葉樹樹幹通常一方面由傾斜樹
Fig. 3. Distribution of residual internal
strain in the erect portion of the trunk
(tree E).
Fig. 4 Peripheral distribution of released
surface strain in the longitudinal direction
in a basal sweeping trunk (tree D).
幹之上側產生拉力另ㄧ方面又由下側產生推力 俾使樹幹恢復直立(Archer 1986)。
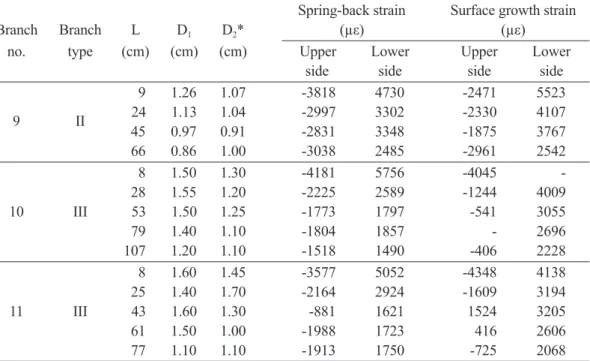
傾 斜 樹 幹 恢 復 直 立 之 機 制 曾 被 解 釋 為 抗 壓 材 於 縱 向 發 生 膨 脹 而 推 動 樹 幹 至 直 立 位 置(Timell 1986b, Niklas 1992, Mattheck and Kubler 1995),惟抗壓材沿樹幹縱向膨脹之機制 仍尚未被完全明瞭。吾等認為傾斜樹幹下側抗 壓材所產生的壓縮應力會在橫斷面上引起一個 平衡彎距(balancing moment),傾斜樹幹必須朝 上彎曲,藉以抵消內部彎距來達成力學平衡, 樹 幹 於 向 上 彎 曲 之 際 , 樹 幹 下 側 產 生 引 張 應 力,上側則產生壓縮應力,此等應力則與原有 內部生長應力相重疊(superpose),因此所測得 之內部應力分佈仍原有內部殘留應力與彎曲應 力之和,且因連年生長之應力累積而更形複雜 (Huang et al. 2001)。吾人有必要作更進一步之 研究,藉以釐清生長應力之生理意義。 四、傾斜樹幹表面微纖維傾斜角之圓周分佈 Figure 5顯示根曲樹幹(樹號D)表面木質部 細胞壁S2層微纖維傾斜角(MFA)之圓周分佈。靠 近下側(180°)之部位因抗壓材之存在,其MFA 有偏高之明顯趨勢,其最大值為31.9°,左右兩 側及上側之MFA範圍在15°~23.5°之間,值得注 意的是左右兩側之MFA並不對稱,右側之MFA 顯然較左側者為高,其理由尚難判斷。MFA與 生長應力具有密切相關性(Okuyama et al. 1986, 1994, Yamamoto et al. 1995),依據統一假說 (unified hypothesis),Guitard et al. (1999)運用 力學模式來探討MFA與生長應力的關係,彼等 計算出生長應力由引張應力轉變為壓縮應力之 臨界MFA約為20°~30°,與Fig. 4相對照,可知傾 斜樹幹下側之高壓縮應力與高MFA,與抗壓材 之存在息息相關,而抗壓材之所以產生壓縮應力 被認為與木質素之膨潤作用有關,抗壓材常較 正常材含較多之木質素與較大的MFA。壓縮生 長應力之大小可用來評估抗壓材之發達程度。 五、樹枝縱向表面生長應變分佈 針葉樹常在傾斜樹幹或樹枝下側產生抗壓 材(Panshin et al. 1964, Timell 1986a),由此特殊 組織所產生的應力不同於由正常材所產生者, 抗壓材產生壓縮應力藉以拘束木材之伸張,並 迫使傾斜樹幹或樹枝朝上方生長或使樹枝與主 幹保持一定的角度。Yoshida et al. (1992a)曾 研究日本扁柏樹枝之生長應力,並指出其生長 應力與物理性質間之關係異於樹幹者,彼等認 為枝條所產生的生長應力有異於樹幹中之正常 材 或 反 應 材 。Table 1顯示樹枝上距離主幹不 同位置之反彈應變與生長應變,樹枝斷面均呈 卵圓形並在下側產生抗壓材,由表可知,樹枝 上側之反彈應變均為收縮應變(負值),而下側 者皆為伸張應變(正值)。樹枝下側之生長應變 皆為正值,而上側者則正負互見,樹枝下側之 生長應變均遠大於樹幹者。樹枝下側之生長應 變,在距主幹距離較近者有隨反彈應變之增加 而增加之明顯趨勢,且前者之數值大於後者, 兩者之平均比值為0.51。Figure 6顯示生長應 變與反彈應變成高度直線相關性(R2 = 0.478)。 另一可見的傾向是樹枝下側之生長應變有隨距 主幹距離之減小而增大之現象,從樹枝斷面之 觀察可知,距離主幹越近則髓心之偏心越大, 靠 近 主 幹 處 之 抗 壓 材 顯 然 較 為 發 達 , 故 能 產 生較大的壓縮應力。我們強調,含有高度發達 的抗壓材之樹枝下側所產生的生長應力係受到 枝條自重引起的重力彎距之影響,而此一彎距 係經由反彈應變表現出來。換言之,重力刺激
Fig. 5. Effects of peripheral positions in
a basal sweeping trunk on the microfibril
angle (tree D).
Table 1. Branch shape, growth strains, and spring-back strains at different distances from
the trunk
Spring-back strain
Surface growth strain
Branch Branch
L
D
1D
2*
(µε)
(µε)
no.
type
(cm) (cm) (cm)
Upper
Lower
Upper
Lower
side
side
side
side
6
3.00
2.65
-3121
4133
1280
6895
28
2.80
2.70
-2139
2495
450
4936
1
I
52
2.40 2.70
-1510
2263
-211
3154
75
2.20
2.90
-323
685
-
609
96
2.20
2.30
-1695
365
-667
119
6
2.90
2.10
-2433
2517
2141
5719
17
2.50
2.20
-1525
2335
629
4950
2
I
30
2.20 2.20
-2093
2197
486
4796
44
2.00
2.20
-401
1992
105
3036
64
1.60
2.05
-1594
1545
-
1650
10
5.30
4.85
-1552
1403
1310
3397
31
4.60
5.00
-973
1247
396
3219
3
I
52
4.25
4.70
-946
853
252
75
4.00
4.10
-393
731
247
72
100
3.75
3.80
-194
845
172
-30
12
2.55
2.40
-2218
1509
436
2905
37
2.30
2.20
-2247
1602
417
2869
4
I
62
2.10
2.00
-1275
1256
-677
2109
89
1.80
1.80
-1254
1273
-206
1495
113
1.50
1.50
-1674
713
-1017
391
16
7.20
5.50
-2063
3230
783
6615
5
I
46
7.20 5.00
-2676
3053
-50
5776
72
6.50
4.30
-1409
2887
490
5893
94
5.90
4.70
-2127
2791
-306
4260
25
6.80
4.70
-
3879
-
5937
55
6.50
4.40
-515
1005
2820
5249
6
I
91
5.90 4.50
-1170
973
982
3524
124
4.90
3.80
-945
983
700
2975
150
4.30
3.80
-1062
1049
27
1606
184
3.90
3.60
-1121
1166
-174
1768
15
3.50
3.30
-1456
2062
-1063
2754
7
II
27
3.30 3.00
-1202
1452
-407
2416
31
3.00
2.20
-848
1014
-34
1284
10
1.13
1.02
-4196
3536
-2789
4032
8
II
33
1.05 1.10
-1117
2537
-911
3186
59
0.83
0.89
-1179
1281
-1674
1588
77
0.78
0.73
-847
1112
-848
1636
(gravity stimulus)可用來說明抗壓材所引起的 高壓縮應力。樹枝形態與枝條自重所引起的彎 曲應力以及生長應力間之平衡有關。當因枝條 之自重所引起之朝下彎距大於生長應力所引起 之朝上彎距時,樹枝便會下垂。反之,樹枝便 朝上彎曲,這便是廣義上的負屈地性(negative gravitropism)現象(Mattheck 1991),其結論是樹 形因反應材(reaction wood)所產生的高生長應力 而受到調整。Table 1顯示水平伸展(或斜傾)枝 條(Type II)及朝下彎曲枝條(Type III)之上側生 長應變為負值,下側者為正值。惟在朝上彎曲 之枝條(Type I),則不論上側或下側,在靠近樹 幹之彎曲部份皆為正值,在靠近末端之直線部 分,則上側為負值,下側為正值,枝條之形態 與生長應變分布之關係圖可歸納如Fig. 7。 六、樹皮之生長應變 就直立樹幹、根曲樹幹之上側、側面及下 側,以及樹枝之上側、側面及下側所測得解放 生長應變值如Table 2。由表可知不論測定部位 如何,樹皮之縱向生長應變皆為負值,顯示有 引張生長應力之存在。有關樹皮生長應力之研 究,僅見於Okuyama et al. (1981)就日本柳杉所
continue
Spring-back strain
Surface growth strain
Branch Branch
L
D
1D
2*
(µε)
(µε)
no.
type
(cm) (cm) (cm)
Upper
Lower
Upper
Lower
side
side
side
side
9
1.26
1.07
-3818
4730
-2471
5523
9
II
24
1.13 1.04
-2997
3302
-2330
4107
45
0.97
0.91
-2831
3348
-1875
3767
66
0.86
1.00
-3038
2485
-2961
2542
8
1.50
1.30
-4181
5756
-4045
28
1.55
1.20
-2225
2589
-1244
4009
10
III
53
1.50
1.25
-1773
1797
-541
3055
79
1.40
1.10
-1804
1857
-
2696
107
1.20
1.10
-1518
1490
-406
2228
8
1.60
1.45
-3577
5052
-4348
4138
25
1.40
1.70
-2164
2924
-1609
3194
11
III
43
1.60
1.30
-881
1621
1524
3205
61
1.50
1.00
-1988
1723
416
2606
77
1.10
1.10
-1913
1750
-725
2068
* L, distance from the trunk; D
1, vertical axial diameter of a branch cross-section; D
2, horizontal axial
diameter of a branch cross-section; Type I, bending upward; Type II, horizontal; Type III, bending
downward.
Fig. 6. Relationship between surface
growth strains and spring-back strains in
a branch.
作之試驗,其結果顯示不論縱向或弦向之測定 皆為負值,顯示有引張應力之存在,與本試驗 之結果相似。