中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/32559
58
0
0
全文
(2) . Index Index. .................................................................................................... I. Figure Index ........................................................................................... III Appendix Index ...................................................................................... IV 致謝…………………………………………………………………………………………………. V Abstract ................................................................................................VII 摘要. .............................................................................................. VIII. Introduction ...............................................................................................1 Viral infection ...........................................................................................1 Immune system and Macrophage .............................................................3 Interferons (IFNs) and Toll-like receptor 3 (TLR3) .................................5 Src kinase ..................................................................................................8 iNOS (inducible nitric oxide synthase) ..................................................10 Aim. ..................................................................................................12. Materials and Methods ...........................................................................13 Cell Culture .............................................................................................13 Preparation of mouse peritoneal macrophages (PEMs) .........................13 Generation of RAW264.7 mcrophages expressing src siRNA ..............14 Reagents and Antibodies ........................................................................14 ELISA for mouse IFN-β .........................................................................15 I .
(3) . RT-PCR for IFN-β ..................................................................................17 Western blotting ......................................................................................19 Nuclear Extraction ..................................................................................20 Statistical analysis ...................................................................................20 Result. ..................................................................................................21. 1.Poly(I:C) induced interferon-β (IFN-β) production in murine RAW264.7 macrophages. .......................................................................21 2.Src family kinase was required for IFN-β production in macrophages in response to poly(I:C). .........................................................................21 3.Enhanced Src expression in RAW264.7 macrophages and peritoneal macrophages (PEMs) in response to poly(I:C).......................................22 4.Src attenuation reduced poly(I:C)-elicited IFN-β production in macrophages. ..........................................................................................23 5.NO/cGMP mediated IFN-β production. ..............................................23 6. Src participated in SNAP-mediated IFN-β production in macrophages. .................................................................................................................24 7.Src-mediated IRF7 nuclear translocation was required for IFN-β production in poly(I:C)-treated macrophages.........................................25 Discussion.................................................................................................27 Reference .................................................................................................31 Figure. ..................................................................................................37. Appendix ..................................................................................................45 II .
(4) . Figure Index Figure 1. Poly(I:C) treatment induces IFN-β production in murine RAW264.7 macrophages. .......................................................37 Figure 2. Poly(I:C)-induced IFN-β was PP2 sensitive. ........................38 Figure 3. Poly(I:C)-mediated IFN-β secretion was inhibited by src-specific siRNA, which could be reversed by ectopic Src. ..................................................................................................39 Figure 4. Poly(I:C)-induced ifn-β transcript was sensitive to inhibitors of inducible nitric oxide synthase (iNOS) and soluble guanylyl cyclase (sGC). .............................................40 Figure 5.iNOS was involved in poly(I:C)-mediated IFN-β production. ..................................................................................................41 Figure 6. SNAP-mediated IFN-β secretion was inhibited by src-specific siRNA, which could be reversed by ectopic Src. ..................................................................................................42 Figure 7. Poly(I:C)-mediated IRF7 nuclear translocation was inhibited by src-specific siRNA, which could be reversed by ectopic Src................................................................................43 Figure 8. The signaling pathway for IFN-β secretion in poly(I:C)-stimulated macrophages. ......................................44 . III .
(5) . Appendix Index Supplementary Figure 1. The inhibitory effect of PP2 on poly(I:C)-induced protein tyrosine phosphorylation. .......................45 Supplementary Figure 2. iNOS is involved in poly(I:C)-mediated macrophage migration. ........................................................................46 Supplementary Figure 3. Poly(I:C) induced Src expression in murine RAW264.7 macrophages and rat peritoneal macrophages (PEMs). .................................................................................................................47 Supplementary Figure 4. Poly(I:C) induced Src expression is dependent on iNOS. ..............................................................................48. . IV .
(6) . 致謝. 碩士生涯的兩年很快就到了,在這兩年在馬老師以及學長姐的指導下, 順利學習一位碩士生該學會的技巧與觀念。對於我以後準備往博士那 不確定的道路,給予我最好的武器與裝備。馬老師常說,她實驗室出 去的學生都能在各種環境下存活,雖然這話有些玩笑意味,不過卻也 透露著在老師實驗室內做任何事都要抱著嚴謹的態度,不論何事都要 抱著謹慎的態度去面對,永遠提早準備完成,才不會在快到時間時慌 亂不已。對於一開始面對老師的"嚴格”的態度,老實講有點不太適應, 雖然早知道做研究,早就該把自己的內心做調整,好在有學長姐的陪 伴下,能將我所面對的壓力給稀釋,也能從他們身上得到非常好的建 議,能夠讓我不自於單獨面對,妙瑩學姐,做實驗的態度及技巧,是 讓我作好實驗做好的範本,青昭學姐,不會忘了你對於我的美工逐一 搖頭的表情,銘育學長,我跟你沒甚麼好說的,我所有學習到的東西 都是從你的身上得到的,不論好的還是壞的(開玩笑)。春霖學長及阿 ken 學長,也給予我不少好方法來面對實驗上所碰到的難題。均晏, 佐駿,婉竹,你們這三個學弟妹,也陪伴我一年,雖然我會常常會罵 你們,東管西管,不過也很感激你們能陪在我身邊,新來的學弟妹也 同樣謝謝你們讓我認識你們。也很感謝我的同學們,在一起求學及做 V .
(7) . 實驗的壓力下,互相給彼此好的意見。最後,我的父母,也是我最想 感謝的人,給予我充足的補充,讓我能夠完整的放心在碩士的路途 上。. VI .
(8) . Abstract. By recognizing double stranded RNA (dsRNA), a product of viral infection, TLR3 can produce type I interferons such interferon alpha and beta (i.e. IFNα/β) that possess anti-viral and immune modulating functions.. As. the. synthetic. analogue. of. dsRNA,. polyinosinic-polycytidylic acid (poly(I:C)) can effectively induce the production of IFNα/β in macrophages. In this study, we investigated the signal transduction pathways in IFN-β secretion elicited by poly(I:C). We observed that poly(I:C)-mediated IFN-β generation was PP2 (an inhibitor for Src family kinases(SFKs)) sensitive in murine RAW264.7 cells. Pharmacological blockade of AG (an iNOS inhibitor) and ODQ (a sGC inhibitor) suppressed poly(I:C)-evoked IFN-β mRNA expression. Either SNAP (a NO donor) or 8-br-cGMP (a cGMP analogue) could rescue these defects. We found that the increment of IFN-β could be attributable to their increased transcripts. Interestingly, knockout of iNOS suppressed poly(I:C)-evoked IFN-β secretion and Src induction. Notably, attenuation of Src by src-specific siRNA reduced poly(I:C)-, and SNAP-enhanced IFN-β production in macrophages, and reintroduction of avain Src could rescue this defect. With these results, we concluded that iNOS was critical for poly(I:C)-mediated IFN-β production and Src played a pivotal role in this process.. Key words: poly(I:C); TLR3; Src; macrophages; interferon-β; iNOS VII .
(9) . 摘要. Toll-like receptor 3 (TLR3)藉著辨識病毒產物雙股 RNA (dsRNA),會 產生干擾素 α 和 β,而兩者皆屬於第一型干擾素。而第一型干擾素具 有對抗病毒入侵和調控免疫反應的功能。現在已知人工合成的 polyinosinic-polycytidylic acid (poly(I:C))能夠有效的刺激巨噬細胞,產 生 type I interferons。在本篇的研究,我們將探討有什麼訊息傳遞路徑 參與在 poly(I:C)刺激巨噬細胞所產生的 IFN-β 的過程中。我們發現 PP2 (Src family kinases 的抑制劑)可以抑制 poly(I:C)刺激老鼠的巨噬 細胞 RAW264.7 所產生的 IFN-β。PP2 會抑制 poly(I:C)刺激下所產生 的 IFN-β 且其 mRNA 也顯著降低。另外一方面,SNAP (a NO donor) 或 8-br-cGMP (a cGMP analogue)皆增加 IFN-β mRNA。故 IFN-β 釋放 量的增加,可由其 mRNA 量的增加來反應。有趣的是,基因剃除 iNOS 的表現確實可以使的 poly(I:C) 刺激巨噬細胞產生 IFN-β 的釋放量及 Src 表現量降低。使用了弱化 Src 的 src-specific siRNA 細胞會使的 poly(I:C)和 SNAP 所刺激的 IFN-β 釋放量下降,不過使用了放回 ectopic Src 的細胞則又能恢復 IFN-β 的產生。根據這些結果,我們可 以得到一個結論,那就是 iNOS 參與 poly(I:C)-induced IFN-β production,. VIII .
(10) . 而且 Src 在這個過程中也扮演重要的角色。. 關鍵字: poly(I:C); TLR3; Src; 巨噬細胞; 干擾素-β; iNOS. IX .
(11) . Introduction Viral infection. Viruses are the smallest organisms that cause infection in our body, much smaller even than other microbes. Viruses consist of one or more genetic materials of DNA or RNA, which contain their genes in the protein coat. In order to survive, virus must attach and invade to living cells. When viruses invade into the cell, they may kill or alter the cell’s functions by replicating their genomes to generate new viruses. Newly formed viruses will result in further infection. Most viruses do cause diseases when they invade normal, living cells, through different ways like inhalation, swallowing, touching or unsafe sex. Culture and eating habits also increase the risk of viral infections.. Various human viruses can attack specific cells in particular parts of our body’s such as respiratory system, digestive system, liver or blood. The main types of viral infection include common cold and influenza (flu). These diseases are frequently caused by viral infection of nose, throat and have some unpleasant symptoms such as stuffy nose, sneezing, sore throat, coughing, body chills, fever, headache, and muscle ache. The skin can also be infected by viral attack. The prominent example is chickenpox. Chickenpox is an infectious disease and spreads very easily in highly populated areas. Furthermore, many serious lethal diseases such 1 .
(12) . as ebola, avian influenza, SARS and AIDS are also transmitted by viral attack and have symptoms like hemorrhagic fever, dehydration, acute respiratory syndrome. Until 2009, an estimated 1.1 million persons are currently living with HIV/AIDS in the US [1]. Recently, new influenza A is a global epidemiological disease by H1N1 virus infection. WHO raised the level from phase 5 to phase 6, 74 countries in 5 WHO regions have been affected [2].. These highly lethal viral pathogens are the world’s. worst epidemic threat.. 2 .
(13) . Immune system and Macrophage. Encounter to viral infections, the immune system offers the main defense in our body. The immune system is composed of organs, tissues and cells that interact to protect the body from external invaders. Several levels are in the two subdivisions of the immune system. Mechanical barriers include skin and mucous membrane. Physical and chemical barriers include coughing, sneezing, vomiting, diarrhea, tears, saliva, and perspiration. Mechanical, physical and chemical barriers are the first lines of defense that belong to non-specific immunity. When they are invalid the second line of defense in innate (non-specific) immunity comes into play. The defensive measures include phagocytosis by granulocytes (the white blood cells called neutrophils, eosinophils and basophils) and monocytes/macrophages, secretion of protective proteins like cytokines and complement proteins. The last lines of defense are cell-mediated (T lymphocytes) and humoral (B lymphocytes) immunity that are pathogen-specific.. The innate immune responses are rapid than the adaptive responses, when viral infection occur macrophages and dentritic cells (DCs) can start the innate immune responses. The myeloid cells in innate immunity are derived from pluripotent hematoietic stem cells in the bone marrow that perform various immune tasks. Macrophages are derived from monocytes that circulate in blood and migrate to tissues where they become resident. Macrophages play an essential role for in innate immunity and have 3 .
(14) . different names dependent on the tissues they reside. For example, Kupper cells in the liver, Microglia in the central nerve system. Macrophages are important in fighting viruses and bacteria, with digestive enzymes to destroy invading microorganisms. Macrophages detect microorganisms through pattern-recognition receptors (PRRs) like Toll-like receptors (TLRs), retinoic acid-inducible gene-I-like receptors (RLRs) and nucleotide-binding oligomerization domain-like receptors (NLRs) [3]. Through these receptors, macrophages detect and phagocytose invaders to present antigens and releas several cytokines (i.e. tumor necrosis factor α, interleukins and interferons) to activate adaptive immune responses.. 4 .
(15) . Interferons (IFNs) and Toll-like receptor 3 (TLR3). Interferons (IFNs) are widely expressed cytokines, was the first to be described by Isaacs and Lindenmann in 1957. It was designated due to its ability “ interfere ” viral replication activities [4]. IFNs have several biological functions. Among them, antiviral and immunomodulatory, are the major IFNs-exerted effects [5]. These cytokines are first line of defense in the innate immunity against viruses, bacteria infections and have important roles to activate antigen-presenting cells and natural killer (NK) cells in the innate immune system. The IFN family has two types: type I interferons and type II interferons. In human, there are many type I interferons include IFN-α and its subtypes [6], IFN-β, IFN-ε, IFN-κ, and IFN-ω [7]. By contrast, there is only one type II interferon, IFN-γ. In the antiviral host immune response, protein kinase R (PKR), 2’-5’oligoadenylate synthases and RNaseL are important antiviral mediators. These IFNs-induced proteins are believed to affect viral multiplication through inhibition of the translation initiation and degradation of viral RNA [8]. IFNs also link the innate responses and the adaptive immune responses. Viral recognition by PRRs in the innate immunity, initiates the activation of the signaling pathway that leads to type I interferons and inflammatory cytokines production that eliminate viruses.. Type I interferons (IFN-β) induction can be triggered by TLRs in immune cell such as macrophages and DCs. In 1985, toll was initially identified to play role in Drosophila development. Later in 1996, the important of toll 5 .
(16) . in Drosophila immunity was described. One year later, a human homologue of the drosophila toll protein (TLR4) was identified that can induce expression of some inflammatory genes [9, 10]. Mammalian TLRs family consists 13 members with different cellular localization that can be recognize by specific pathogen-associated molecular patterns (PAMPs). As TLR1, TLR2, TLR4, TLR5, TLR6 localize to the cell surface and recognize lipid and protein PAMPs, TLR3, TLR7, TLR9 are expressed in intracellular compartments such as endosome to detect viral nucleic acid PAMPs. Each TLR has its own signaling pathway and induces specific biological responses. By utilizing different adaptors, TLRs can trigger several signaling pathways. While most TLRs utilize MyD88-dependent pathway like TLR4, TLR2 can utilize myeloid differentiation factor 88 (MyD88), TLR3 triggers signaling pathways in a MyD88-independent manner.. TLR3 detects viral dsRNAs, that can be mimicked by the synthetic analog polyinosinic acid-cytidylic acid (Poly(I:C)). TLR3-deficient mice showed reduced production type I IFNs and proinflammatory cytokines [11]. TLR3 signals only via the TIR domain-containing adapter inducing interferon-β (TRIF) that is also called TIR domain–containing adapter molecule-1 (TICAM-1) [12]. Generation of TRIF-deficient mice hampered TLR3-mediated activation of IFN-β and IFN-β inducible genes production [13]. TRIF interacts with TRAF3, TBK1 and inducible IκB kinase. (IKK-i).. TBK1. and. IKKi. mediate. phosphorylation. of. interferon-regulatory factors, IRF3 and IRF7. Phosphorylated IRF3 and 6 .
(17) . IRF7 dimerizes and translocated to the nucleus to activate the expression of type I IFN and IFN-inducible genes [14].. 7 .
(18) . Src kinase. The viral src (v-src) was the first indentified oncogene in Rous sarcoma virus (RSV) by Peyton Rous in 1911. Until 1976, this oncogene genetic and physical was indentified. As a counterpart of v-src, cellular src (c-src) was identified as a proto-oncogene in normal cells. Mounting evidence indicates that c-src (the protein encoded by c-src) play an important role in normal cellular functions [15].. Src is a non-receptor tyrosine kinase that contains several structural features. There are several domains in this Src protein. Form N to C terminus, there are Src Homology 4 (SH4), SH3, SH2 and catalytic/regulatory domains. The N-terminus of c-Src is myristoylated at Gly2, and becomes membrane-associated. As SH3 domain can interact with. proline-rich. phosphorylated. sequences.. tyrosine.. SH2. Therefore,. domain both. can. domains. interact can. with. mediate. protein-protein interaction. The catalytic and regulatory domains are important to regulate Src activity. Tyr-416 and Tyr-527 are two major sites whose phosphorylation can modulate Src activity. While phosphorylation of Tyr-416 resultes in Src activation, phosphorylation of Tyr-527 decrease its activity. This is become phosphorylated Tyr-527 (PiTyr527) will interact with SH2 domain, renders Src in an inactive conformation. However, dephosphorylated Tyr-527 and Pi-Tyr416 will render in an active Src conformation. In this report, we use the level of Pi-Tyr416 as an indicator Src activation. 8 .
(19) . Since the discovery of Src, Yes, Fgr, Fyn, Lck, Lyn, Blk, and Hck have been identified as membranes of Src family kinases (SFKs) [15]. Despite Src family members are expressed in a wide variety of tissues, Lyn, Fgr, Hck, Lck, and Blk are predominantly expressed in hematopoietic cells. Accumulated evidence indicates the participation of tyrosine kinases in LPS-stimulated macrophages. Through Lyn, Fgr and Hck are largely expressed in monocytes and macrophages, macrophages derived from lyn−. / −. fgr−. / −. hck−. / −. triple knockout mice still retained full LPS. responsiveness [16]. This implicates the possible involvement of an Src family kinase in LPS-induced macrophage activation. Interestingly, we previously reported, found Src up-regulation in LPS-stimulated macrophages both in vitro and in vivo [17]. In this report we present evidences demonstrating the induction of c-Src in RAW264.7 and peritoneal macrophages in response to poly(I:C)-mediated IFN-β production.. 9 .
(20) . iNOS (inducible nitric oxide synthase). Nitric oxide (NO) has been identified as an important signaling molecular involved in many biological functions, immune, vasodilatation. NO synthesis has been found in a variety of different cell types. With the enzyme nitric oxide synthase (NOS), NO direct formation and release. There are three distinct isoforms of NOSs, neuronal and endothelial NOS that are constitutively expressed whereas inducible NOS induced by cell-mediated immunity and inflammation. Unlike nNOS and eNOS, iNOS is independent of Ca+2 /calmodulin regulation.. In the immune system, NO and its derivatives are important to the antimicrobial. activity. especially. when. macrophages. activation.. Macrophages generate NO from L-arginine through the inducible isoform of nitric oxide synthase (iNOS) by LPS stimuli. iNOS expressing in several cells including endothelium, hepatocytes, monocytes, mast cells, macrophages and smooth muscle cells participates in many signaling pathway. Depending on the cytokines or TLRs ligands stimulus in the immune cells (macrophages and DCs) increase iNOS expressing causes NO production, than following activation of sGC results cGMP production. NO activates NF-κB and up-regulates iNOS (positive feedback). NO and cGMP are second messenger molecules allow smooth muscle. to. relax. and. immune. cells. activation,. enhance. anti-microorganisms activity. Furthermore, in 2008 we demonstrated lipopolysaccharide-mediated Src 10 .
(21) . induction and macrophages migration was inducible nitric-oxide synthase (iNOS) [18]. Later, similar study indicated Src kinase-mediated phosphorylation stabilizes iNOS in normal cells and cancer cells [19]. In this. study,. we. demonstrated. that. iNOS. was. required. in. poly(I:C)-mediated IFN-β production which was PP2-sensitive. And attenuation of Src by src-specific siRNA reduced poly(I:C)-, SNAP-, enhanced. IFN-β. production. in. macrophages.. Interestingly,. pharmacological blockade like AG (a iNOS inhibitor), ODQ (a sGC inhibitor) or knockout of iNOS suppressed poly(I:C)-evoked type I interferons (IFN-β) secretion. By RT-PCR, we found that the increment of IFN-β could be attributable to their increased transcripts. In doing this we have evaluated the role of IFN-β is correlated with the activation of Src and IRF proteins that depends on iNOS.. 11 .
(22) . Aim. Macrophages play a critical role in inflammatory responses. Macrophages have antigen presentation, phagocytosis, and immunomodulation through production of various cytokines and growth factors. Previously, we demonstrated poly(I:C)-mediated macrophage migration. Poly(I:C) used to mimic viral dsRNA, is recognized by TLR3. Poly(I:C), a byproduct of viral infection can promote macrophage migration and anti-viral cytokines production in inflammatory processes. TLR3 mediates a more potent antiviral response than other TLRs [20]. This is why we study TLR3 pathway investigated downstream signaling activation mediated anti-viral cytokines IFN-β expression. Previous research indicates c-Src tyrosine kinase associates with initiate TLR3 antiviral signaling in human monocyte-derived dendritic cell [21]. In this report we want to know the induction of c-Src in RAW264.7 and peritoneal macrophages in response to poly(I:C)-mediated IFN-β production.. 12 .
(23) . Materials and Methods Cell Culture. RAW264.7 (American Type Culture Collection, murine macrophage cell line ), was cultured and propagated in RPMI 1640 medium (Invitrogen) supplemented with 10% fetal bovine serum (JRH Biosource), 100unit/ml-100μg/ml. Penicillin-streptomycin. (Sigma). and. 2mM. L-glutamine(Sigma) at 37 °C in humidified atmosphere of 5% CO2 and air.. Preparation of mouse peritoneal macrophages (PEMs). C57BL/6JNarl male mice were purchased from National Laboratory Animal Center in Taiwan and maintained in the animal center of China Medical University at Taichung. The animal room was maintained on 12 hour light and 12 hour dark cycle with a standard temperature and humidity. All mice were grown at 8-10 weeks old, sacrificed under anesthesia, and used to obtain cells from peritoneal exudates. All procedures adhered to the “Guide for Care and Use of Laboratory Animals” (National Research Council, USA) and were approved by the animal experiment committee of China Medical University. Mouse peritoneal macrophages (PEMs) were obtained from wild-type and iNOS-null mice by lavage with 10ml of cold PBS per mouse at 3 days after intraperitoneal injection of 3% 1ml thioglycollate (MERCK) in PBS. 13 .
(24) . The PEMs were seeded in culture plates and incubated at 37 °C in humidified 5% CO2 and 95% air to allow macrophages adherence. After 6 hours, the non-adherent cells were removed by washing with warm PBS and remaining cells were used for experiment.. Generation of RAW264.7 mcrophages expressing src siRNA. To generating the cells expressing nonspecific siRNA (Ctrl-1, Ctrl-2) or src siRNA (siRNA), RAW264.7 mcrophages were transfected with plasmid. DNA. pLKO.1-msrc. (puro). (targeted. sequence:. 5’-. -CGAGCCGCCAATATCCTAGTA-3’ or the negative control pSilencer plasmid (not present in mouse genome and provided by Ambion Inc.) by the Lipofectamine followed by puromycin or hygromycin selection. RAW264.7 mcrophages expressing src siRNA were cotransfected with pBabe (hygro) DNA and a plasmid DNA encoding. avian. c-Src [22]. to generate Src-attenuated cells expressing ectopic Src (siRNA-1/Src6, siRNA-1/Src15).. Reagents and Antibodies. Polyinosinic-polycytidylic acid (poly(I:C)), aminoguanidine hemisulfate (AG),. 1H-[1,2,4]oxadiazolo[4,3-α]quinoxalin-1-one. S-nitroso-N-acetylpenicillamine. (SNAP),. 8-bromo-cyclic. (ODQ), guanosine. monophosphate (8-Br-cGMP) were obtained from Sigma. Thioglycollate and. 4-Amino-5-(4-chlorophenyl)-7-(t-butyl)pyrazolo-[3,4-d]pyrimidine 14 . .
(25) . (PP2) were obtained from MERCK. The primary antibodies used were actin (Sigma); FAK and iNOS (Upstate); Pi-Y861 FAK and Pi-Y416 Src (BIOSOURCE International); Lyn, Fgr and Hck (Santa Cruz); IRF3, IRF7 and PCNA (Santa Cruz); HRP-goat anti-mouse antibody and HRP-goat anti-rabbit antibody (Jackson). Src-specific mouse monoclonal antibody are provided by Dr. Leu in National Cheng Kung University, Tainan.. ELISA for mouse IFN-β. RAW264.7 and its based, Ctrl-1, Ctrl-2, siRNA, siRNA-1/Src6, and siRNA-1/Src15 cells were induced with poly(I:C) for 48 hours. The culture supernatants were examined by enzyme-linked immunosorbent assay (ELISA). The IFN-β ELISA kit was purchased from PBL Biomedical Laboratories. Murine IFN-β production was detected by ELISA according to the manufacturer’s procedure (PBL Biomedical Laboratories) [23]. Produces are following:. Prepare Wash Buffer as follows: Dilute the 50 ml of 10x wash concentrate with 450 ml of distilled or deionized water. The Final wash solution should be stored at 2-8°C and mixed thoroughly before use. All the wash steps should be performed at room temperature (22-25°C). Wash steps can be performed manually with a squirt bottle or by using an automated plate washer. Prepare a 1:10 working stock of the Mouse IFN-β standard by pipetting 15 .
(26) . 10 μl of the Mouse IFN-β Standard in 90 μl of sample diluent. Prepare fresh dilutions of the standard curve for each assay run. Determine the number of microplate wells required to test the desired number of samples and standard curve. Place precisely 100 μl of the interferon samples and standard curve prepared in Step 2 in individual wells of the microtiter plate, at least in duplicate. Remaining strips should be resealed with desiccant in the foil pouch and stored at 2-8°C for future use. Cover the microtiter plate with one of the enclosed plastic plate sealers and incubate for 60 minutes in a closed chamber at 22-25°C. Optionally, the incubations can be carried out at room temperature, keeping the plate away from drafts and other temperature fluctuations. After the 60 minutes incubation, wash three times (3x) with prepared wash solution (at least 250 μl/well). Washes can be performed manually with a squirt bottle or by utilizing an automated plate washer. During the incubation period, prepare the Antibody Solution for use in step 6. Dilute the Antibody Concentrate with Concentrate Diluent. Refer to the lot specific Certificate of Analysis (COA) for the correct amounts of Antibody Solution to prepare. Store used undiluted antibody concentrate at 2-8°C for future use. Add 100 μl of the Antibody Solution prepared using the COA, to each well. Cover the microtiter plate with one of the enclosed plastic plate sealers and incubate for 60 minutes in a closed chamber at 22-25°C. After the 60 minutes incubation, wash three times (3x) with prepared wash solution (at least 250 μl /well). 16 .
(27) . During the incubation period, prepare the HRP solution for use in step 8. Dilute the HRP Conjugate Concentrate with HRP Conjugate Diluent. Refer to the lot specific Certificate of Analysis (COA) for the correct amounts of HRP Solution to prepare. Add 100 μl of the HRP solution prepared using the COA, to each well. Cover the microtiter plate with one of the enclosed plastic plate sealers and incubate for 60 minutes in a closed chamber at 22-25°C. After the 60 minutes incubation, wash three times (3x) with prepared wash solution (at least 250 μl/well). Add 100 μl of the TMB Substrate Solution to each well. Incubate for 15 minutes in a closed chamber at 22-25°C. Add 100 μl of Stop Solution to each well. Mix gently. Using a microplate reader, determine the absorbance at 450 nm within 5 minutes after the addition of the stop solution.. RT-PCR for IFN-β. Total RNA was isolated from RAW264.7 mcrophages, Ctrl-1, siRNA, siRNA-1/Src6, and siRNA-1/Src15 cells by utilizing the Trizol reagent (REzol, Protech, Taiwan) according to the manufacturer's protocols. Transfer these cell lysate to a microfuge tube, and incubate the homogenate for 5 minutes at room temperature. Add 0.2 mL of chloroform (Sigma) per 1 mL of REzolTM C & T. Cover the samples tights and shake vigorously for 15 seconds and incubate them at room temperature for 2 minutes. Centrifuge the samples at 14000 xg for 15 17 .
(28) . minutes at 4°C. After centrifugation, the homogenate form two phase: the lower red phenol-chloroform phase and the colorless upper aqueous phase. The RNA remains exclusively in the aqueous phase (with a volume of approximately 0.6 mL), whereas the DNA and proteins remain in the interphase and organic phase. Carefully transfer the aqueous phase (0.5 mL) to a new microfuge tube without disturbing or touching the interphase. Add an equal volume (0.5 mL) of isopropanol (Sigma) and mix gently. Incubate the samples for 10 minutes at room temperature and centrifuge them at 14000 xg for 10 minutes at 4°C. The RNA precipitate (often invisible before centrifugation) forms a white pellet at the bottom of the tube. Carefully remove the supernatant, wash the RNA pellet in 1 mL of 75% ethanol (in DEPC-treated water) by vortex-mixing and subsequence centrifugation at 14000 xg for 5 minutes at 4°C. Carefully remove the ethanol and dry the pellet by air or under vacuum. Do not dry the RNA by vacuum. It is important not to over-dry the pellet because this makes it harder to dissolve the RNA. Dissolve the RNA pellet in 50 μl of DEPC-treated water. The RNA was quantified and checked for purity and condition by spectrophotometry at a wavelength of 260 nm. RNA concentration. (μg/ml). =. Dilution. factor. ×. RNA (OD260). ×. (40μg/ml/OD260). The RNA samples of 1 μg were then subjected to reverse ranscription-polymerase chain reaction (RT-PCR) by one tube reaction system-Fast Run™ HotStart RT-PCR kit (Protech technology, Taiwan) to determine the mRNA level of IFN-β. The sequences of primers. used. for. murine. ifnβ. CCACAGCCCTCTCCATCAACTATAAGC 18 . transcript (sense). were and.
(29) . AGCTCTTCAACTGGAGAGCAGTTGAGG. (antisense).. The. amplification cycles included 94 °C for 60 s, 60 °C for 60 s, and 72 °C for 120 s. After 30 cycles of amplification, the PCR products were subjected to electrophoresis on a 0.5% agarose gel and then visualized by ethidium bromide staining. The mRNA of gapdh served as control for the sample integrity and loading.. Western blotting. Cells were washed three times with PBS and re-suspended in lysis buffer (4.38 g NaCl, 3.0285 g Tris-base, 1.25 g deoxycholate, 5 ml IGEPAL CA-630), with proteinase inhibitor (sodium orthovannadate, EGTA, aprotinin, and PMSF). The protein concentration was determined by colorimetric assay (Bio-Rad). Sample containing equal protein (50 μg) were separated by 8% SDS-PAGE and transferred onto polyvinylidene difluoride membranes (Millipore, Belford, MA). The membranes were blocked with blotting buffer containing 0.05% Tween 20 and 3% BSA in a shaker for 1 hour and then incubated overnight with first antibodies at 4°C or incubated with first antibodies 3 hours at room temperature. The membranes were washed with TBST three times and then incubated with Horseradish peroxidase conjugated secondary antibody (Jackson). The membranes were washed with TBST three times. The proteins of interest were detected by exposure the autoradiograph to X-ray films (Kodak).. 19 .
(30) . Nuclear Extraction. Cells were washed three times with PBS and re-suspended with 200 μl hypotonic buffer containing 10 mM HEPES, pH 7.9; 10 mM KCL; 0.1 mM EDTA; 0.1 Mm EGTA; DDH2O and proteinase inhibitor 10X (10 mM DTT, 10 mM PMSF, 10 mM NaF, 10 mM Na3VO4, DDH20) and incubated 5 minutes on ice to allow cells to swell. Centrifuge the homogenate at 13200 g at 4°C for 5 minutes. The cytoplasmic fraction was carefully aspirated. The pellet was washed three times with PBS and re-suspended with extraction buffer containing 20 mM HEPES, pH 7.9; 400 mM NaCl; 1 mM EDTA; 1 mM EGTA; DDH2O and proteinase inhibitor 10X and vigorously votexed. Centrifuge the homogenate at 13200 g at 4°C for 15 minutes. Collect the supernatants (nuclear extracts). The protein concentration was determined by colorimetric assay (Bio-Rad).. Statistical analysis. All experiments were repeated at least three times using independent culture preparations. Values are shown as meanേSD. Student’s t test was used to calculate the statistical significance of the experiments. A value of P<0.05 was considered statistically significant.. 20 .
(31) . Result 1. Poly(I:C) induced interferon-β (IFN-β) production in murine RAW264.7 macrophages.. Each TLR has its own signaling pathway and induces specific biological responses. TLR3, which localizes to endosomes can recognize viral dsRNA. Unlike the other endosomal TLRs, TLR3 induces its signals through MyD88-independent pathway. The TLR3-TRIF signaling pathway can activate subsequent IRFs (interferon regulatory factors), MAPKs and NF-κB to increase production of type I interferons and pro-inflammatory cytokines. In this study, we use polyinosinic acid-cytidylic acid (Poly(I:C)) to mimic viral dsRNA. Murine RAW264.7 macrophages were incubated with poly(I:C) (20 μg/ml) for 24, 48 and 72 hours. Conditioned medium were harvested from each time point and the amount of IFN-β was examined by ELISA. As demonstrated in Figure 1, increased IFN-β was detected in poly(I:C)-treated macrophages as compared to non-treated ones. 2. Src family kinase was required for IFN-β production in macrophages in response to poly(I:C).. Previous study indicated LPS stimulates murine macrophages activation can increase protein tyrosine phosphorylation [24]. Two years later, others demonstrated Src family kinases Lyn, Fgr, Hck play an important role in LPS stimulates macrophages activation [25, 26]. We want to know 21 .
(32) . whether poly(I:C) (20 μg/ml) treatment will increase protein tyrosine phosphorylation, Previously, we demonstrated that show poly(I:C) (20 μg/ml) increased the whole protein tyrosine phosphorylation (pTyr) in macrophages. And PP2 treatment greatly reduced this poly(I:C)–mediated phosphotyrosine enhancement, suggesting the involvement of SFKs in this process (Supplementary Figure 1).. To confirm this point, RAW264.7 cells were pretreated with PP2 (10 μM), and then stimulated with poly(I:C) (20 μg/ml) for 48 hours. As demonstrated in Figure 2A, PP2 (a SFK inhibitor) greatly suppressed the production of IFN-β in poly(I:C)-stimulated macrophages. Consistently, PP2 almost wiped out ifn-β transcript (Figure 2B). These findings pointed out Src family kinases were involved in poly(I:C)-elicited increased of the level of transcript and protein expression of ifn-β gene.. 3. Enhanced Src expression in RAW264.7 macrophages and peritoneal macrophages (PEMs) in response to poly(I:C).. Given that SFKs were crucial in poly(I:C)-induced IFN-β production (Figure 2), therefore , we examined the expression Src and its myeloid relatives (i.e. Lyn, Fgr, and Hck) in poly(I:C)-treated RAW264.7. As exhibited in Supplementary Figure 3, unlike the most constant expression of Lyn, Fgr, and Hck, the amount of Src was greatly induced in poly(I:C)-treated RAW264.7. Notably, similar phenomenon was also observed in PEMs in response to poly(I:C). 22 .
(33) . 4. Src attenuation reduced poly(I:C)-elicited IFN-β production in macrophages.. Since Src is the only SFK whose expression can be induced by poly(I:C), therefore its involvement in poly(I:C)-mediated IFN-β production in macrophages was attempt. To answer this question, nonspecific and src-specific siRNA–bearing plasmids were transfected into RAW264.7 macrophages to obtain the control (Ctrl-1, Ctrl-2) and Src-attenuated (siRNA-1) cells. As shown in Figure 3, Src induction was observed in RAW264.7 and control (Ctrl-1, Ctrl-2) cells but not in Src attenuated (siRNA-1) cells following poly(I:C) stimulation. Interestingly, the expression of Lyn was not significantly altered in the presence or absence of poly(I:C) in RAW264.7, control and siRNA cells. Further analysis of IFN-β production in these cells revealed that poly(I:C)-induced IFN-β production was significantly suppressed in Src attenuated (siRNA-1) cells as compared to RAW264.7 and control cells following poly(I:C) treatment. To confirm the contribution of Src, we checked whether Src-attenuated cells ectopic Src (siRNA-1/Src6, siRNA-1/Src15) could rescue this event. As shown in Figure3, suppressed IFN-β production by attenuated Src could be restored by ectopic Src. Thus, the pivotal role of Src in poly(I:C)-induced IFN-β production was corroborated.. 5. NO/cGMP mediated IFN-β production.. Our previous study demonstrated that poly(I:C)-mediated macrophage 23 .
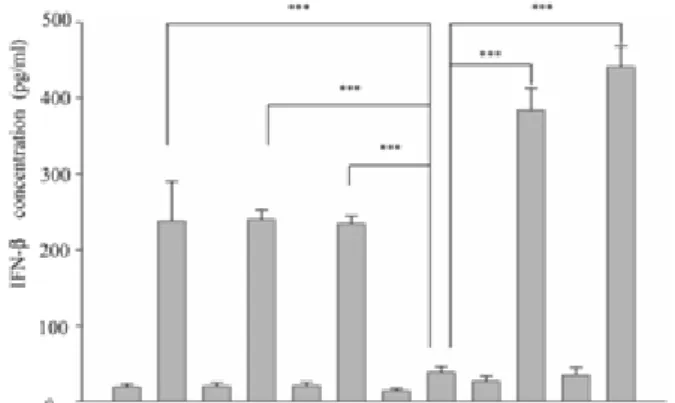
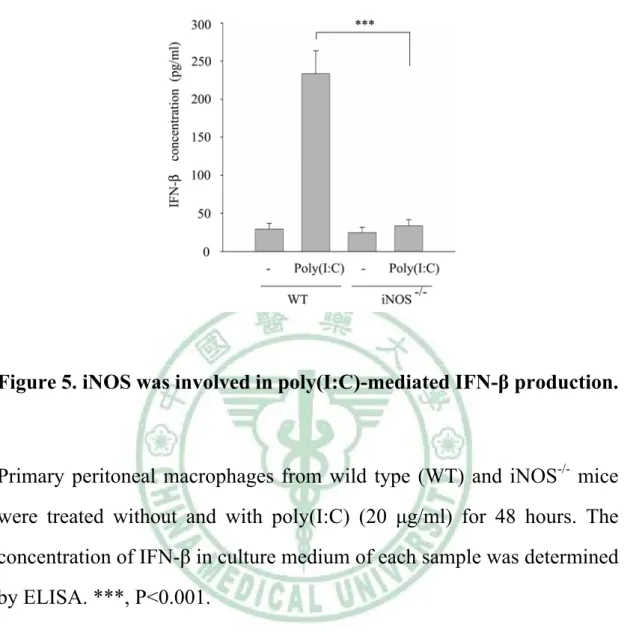
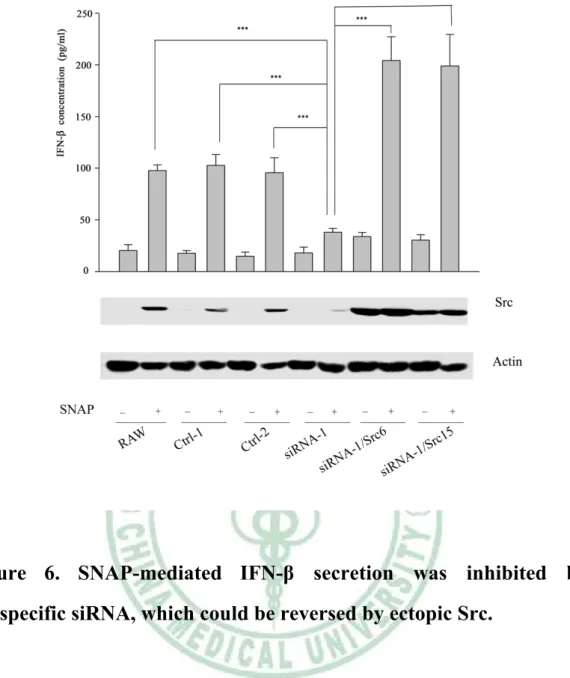
(34) . migration was iNOS-dependent. Due to Src expression was NO-inducible, therefore we wonder the involvement of iNOS in poly(I:C)-induced IFN-β production in macrophages. To answer this question, we pretreated RAW264.7 with AG (an iNOS inhibitor) (2mM) or ODQ (a sGC inhibitor) (100 μM) for 30 minutes, then cells were stimulated without and with poly(I:C) (20 μg/ml). Total RNAs extracted from each sample were analyzed by reverse transcription polymerase chain reaction (RT-PCR). As demonstrated Figure 4A, AG and ODQ abrogated poly(I:C)-elicited increase of ifn-β transcript.. Notably, SNAP (100 μM) and 8-Br-cGMP. (cGMP analogue, 100 μM) significantly increased the amount of ifn-β transcript (Figure 4B).. To confirm the importance of iNOS in IFN-β production in macrophages by poly(I:C), the IFN-β expression of PEMs derived from WT and iNOS-/- mice treated with or without poly(I:C) was analyzed. As shown in Figure 5, compared with WT PEMs, reduced IFN-β production by poly(I:C) treatment was observed in iNOS null PEMs. These findings indicate. the. participation. of. the. NO/cGMP. pathway. in. poly(I:C)-mediated IFN-β production.. 6. Src participated in SNAP-mediated IFN-β production in macrophages.. As shown in Figure 5, we demonstrated that iNOS played an important role. in. poly(I:C)-mediated. IFN-β 24 . . production. in. macrophages..
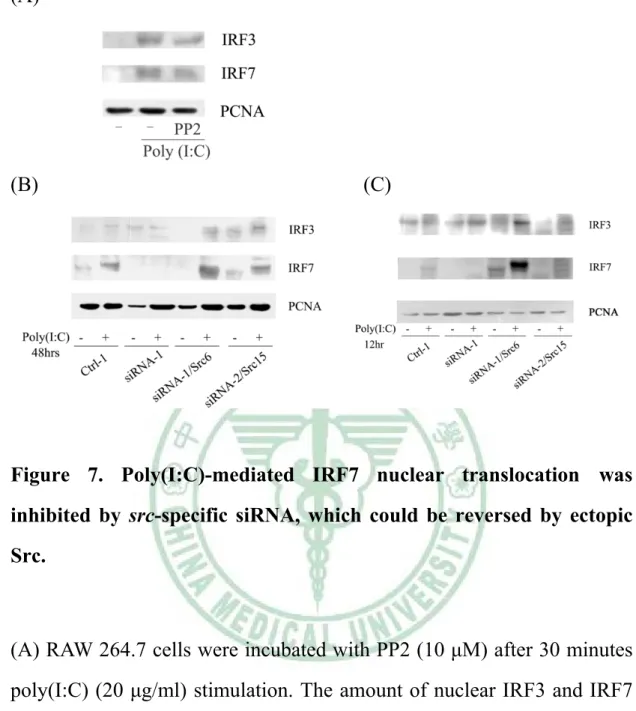
(35) . Interestingly, we also found that poly(I:C)-induced Src expression was abrogated in iNOS knockout macrophages (Supplementary Figure 4). Since Src expression was NO-inducible, therefore we wondered that Src was critical in NO-elicited IFN-β production in macrophages. To confirm this speculation, RAW264.7 and its derived control (Ctrl-1, Ctrl-2) and siRNA (siRNA-1) cells were stimulated without or with SNAP (NO donor) (100 μM) for 48 hours and the secretion of IFN-β in these cells was determined. As shown in Figure 6, we observed that SNAP-mediated Src expression and IFN-β production were significantly suppressed in Src attenuated (siRNA-1) cells as compared to RAW264.7 and Ctrl-1 and -2 cells following SNAP treatment. Interestingly, ectopic src could rescue these events. From these results, we confirmed that by induction of Src, iNOS was critical in IFN-β production.. 7. Src-mediated IRF7 nuclear translocation was required for IFN-β production in poly(I:C)-treated macrophages.. Poly(I:C)-mediated IFN-β production required the participation of Src in macrophages (Figure 3). Since IRF3 and IRF7 were reported to be pivotal in ifn-b gene transcription, thereby we wondered whether Src affected nuclear translocation of IRF3 and IRF7. As exhibited in Figure 7A, poly(I:C)-mediated nuclear translocation of IRF3 and IRF7 was PP2-sensitive. To study the role of Src in poly(I:C)-mediated nuclear translocation of IRF3 and IRF7, nuclear extracts derived from RAW264.7, Ctrl-1, siRNA-1, siRNA-1/Src6 and siRNA-1/Src15 cells following 25 .
(36) . poly(I:C) stimulation were analyzed. As shown in Figure 7B, that poly(I:C)-mediated increase of nuclear IRF7 was significantly suppressed in Src attenuated (siRNA-1) cells as compared to Ctrl-1 cells following 48 hours poly(I:C) treatment. Notably, ectopic Src rescue these events. Due to the amount of poly(I:C)-induced nuclear IRF3 was not suppressed by Src attenuation. Since, IRF3 was preceded of IRF7, so we chose 12 hour instead of 48 hour poly(I:C) treated cells to analyze the translocation of IRF3 and IRF7 to nucleus. As demonstrated in Figure 7C, the amount of poly(I:C)-enhanced nuclear IRF7 was suppressed in Src attenuated (siRNA-1) cells, but the amount of nuclear IRF3 was not changed in siRNA-1 cells. These results indicated that by augmenting the amount of IRF7, Src was required for poly(I:C)-mediated IFN-β production in macrophages.. 26 .
(37) . Discussion In this study, we found that iNOS and Src play critical roles in poly(I:C)-mediated IFN-β production in macrophages in vitro (Figure 6) and in vivo (Figure 5, Supplementary Figure 4). The function of type I IFNs is well characterized and they are known to be essential for a powerful host response against viral infection. There are four effector pathways of the IFN-mediated antiviral responses: the ISG15 ubiquitin-like. pathway,. the. Mx. GTPase. pathway,. the. 2’,. 5’-oligoadenylate-synthetase-directed ribonuclease L pathway, and the protein kinase R pathway. These pathways degrade viral RNA, block viral transcription, inhibit translation and modify protein function to control viral replication.. ISG15 functions to prevent virus-mediated degradation of IRF3, and increases the induction of IFN-β to against viral infection [27]. ISG15 target antiviral effector proteins such as Mx, PKR and RNaseL [28]. Following viral infection, Mx proteins produce from an IFN-stimulated response element (ISRE) to block viral gene transcription [29]. Following activation. by. viral. double-stranded. RNA. (dsRNA),. 2’,. 5’-oligoadenylate-synthetase expression is upregulated by type I IFNs [30], and activated the constitutively expressed inactive ribonuclease L (RNaseL). The binding of 2’, 5’-oligoadenylate-synthetase to RNaseL enables RNaseL activation and be capable to cleave virals RNA. Protein kinase R (PKR) expression is also upregulated by type I IFNs. PKR is 27 .
(38) . activated directly by viral RNA, and can suppress viral proteins translation ability [31] by phosphorylation of eukaryotic translation initiation factor 2α.. IFN-β production is one of the means of innate immune system to against viral infection. Innate immune cells include dendritic cells (DCs), macrophages, neutrophils, and others. In host defense, the innate immune cells recognize microbial pathogens through distinct pattern recognition receptors (PRRs). Through induction of costimulatory molecules and cytokines to activation and differentiation of T lymphocytes, TLRs can trigger adaptive immune responses. TLRs detect microbial nucleic acids, such as double-stranded RNA (dsRNA) (TLR3), single-stranded RNA (ssRNA) (TLR7), and dsDNA (TLR9). TLR3 mainly detect viruses like reovirus, rotavirus in the gastrointestianl system, and Influenza A virus from infected cells but not in the direct viruses themselves [32]. TLR3 can activate its downstream signaling pathways to produce type I IFNs.. To mimic viral infection, we use poly(I:C), which is a synthetic analog of double-stranded RNA (dsRNA), a molecular pattern associated with viral infection and virus replication by-product recognized by TLR3 [33]. TLR3 is activated by extracellular dsRNA and endogenous mRNA [34], which escaping from damaged tissue or in endocytosed cells. TLR3 can not only detect exogenous danger signals, but also sense distress from damaged or stressed tissue. TLR3-mediated type I IFN signaling is only one of TLR3-mediated signaling pathway. All of TLR family membranes, 28 .
(39) . only. TLR3. use. MyD88-independent. adaptor. protein. TRIF/. Toll-interleukin 1 receptor domain (TIR)-containing adaptor molecule TICAM-1 [13, 35]. Through TRIF/ TICAM-1-mediated activation of transcription factors (i.e. NF-κB, AP-1 and IRF3/7), transcription of ifn-b and proinflammatory cytokine encoding genes can be induced [36]. In this study, we demonstrated the non-recetpor tyrosine kinase Src participated in TLR3-mediated IFN-β production in macrophage. Interestingly, c-Src is required for TLR3-mediated activation in dentritic cells [21]. Previously, Src Family Kinases (SFKs) were documented to be involved in inflammatory cytokines production in human macrophages response to HIV-1 envelope protein SFKs were involved [37]. Recently mounting evidence indicated encephalomyocarditis virus-mediated COX2 expression in macrophages is regulated by SFKs [38]. The ability of PP2 and Src attenuation to reduce poly(I:C)-induced IFN-β expression (Figure 2B, Figure 3) and nuclear translocation of IRF7 (Figure 7), suggests that Src participates in poly(I:C)-mediated downstream signaling in the innate immune cells. Furthermore, we also found Src was involved in poly(I:C)-induced proinflammtory cytokines TNFα production in macrophages (data not shown).. Previously, we have demonstrated that LPS-induced Src induction and macrophage migration are required for NO/cGMP [18]. Expression of iNOS and NO production is well known to be an important effector in macrophages in response to viral infection [39, 40]. It is known that iNOS and NO inhibit viral replication and play a role in the host defense against 29 .
(40) . viruses [41, 42]. Using poly(I:C) to mimic viral infection, we also demonstrated suppressed reduced migration in iNOS null PEMs as compared to WT PEMs (Appendix 2), indicating iNOS expression plays a critical role in macrophages activation including migration, cytokines production. In this study, we use pharmacological blockade, AG (iNOS inhibitor) and ODQ (sGC inhibitor), suppressed poly(I:C)-evoked increment of ifn-b mRNA (Figure 4A), and SNAP (a NO donor) or 8-br-cGMP (a cGMP analogue) could increase the abundance of ifn-b transcript (Figure 4B). With these findings, we conclude that iNOS also plays a critical role in antiviral cytokines production. Since exogenous IFN-β can accelerate the induction of iNOS [43], and iNOS is involved in IFN-β production, therefore there might exist a loop to amplify poly(I:C)-mediated effects. At the same time, we also found Src induction is dependent on iNOS (Supplementary Figure 4).. According to our results, we demonstrate that poly(I:C)-mediated IFN-β production, which is the requirement of iNOS in the innate immune cells macrophages. We suggest that Src seemed to be one of the downstream targets in this process and induction by iNOS. In the immune system, macrophages are important for our body to against infection agents. We clear up iNOS and Src playing important roles in poly(I:C)-mediated IFN-β production in macrophages .. 30 .
(41) . Reference 1.. Heffelfinger, J.D., et al., Pandemic influenza: implications for programs controlling for HIV infection, tuberculosis, and chronic viral hepatitis. Am J Public Health, 2009. 99 Suppl 2: p. S333-9.. 2.. New influenza A (H1N1) virus: global epidemiological situation, June 2009. Wkly Epidemiol Rec, 2009. 84(25): p. 249-57.. 3.. Kawai, T. and S. Akira, The roles of TLRs, RLRs and NLRs in pathogen recognition. Int Immunol, 2009. 21(4): p. 317-37.. 4.. Isaacs, A. and J. Lindenmann, Virus interference. I. The interferon. Proc R Soc Lond B Biol Sci, 1957. 147(927): p. 258-67.. 5.. Takaoka, A. and H. Yanai, Interferon signalling network in innate defence. Cellular Microbiology, 2006. 8(6): p. 907-922.. 6.. Pestka, S., The human interferon-alpha species and hybrid proteins. Semin Oncol, 1997. 24(3 Suppl 9): p. S9-4-S9-17.. 7.. Platanias,. L.C.,. Mechanisms. of. type-I-. and. type-II-interferon-mediated signalling. Nat Rev Immunol, 2005. 5(5): p. 375-386. 8.. Gantier, M.P. and B.R. Williams, The response of mammalian cells to double-stranded RNA. Cytokine Growth Factor Rev, 2007. 18(5-6): p. 363-71.. 9.. Arancibia, S.A., et al., Toll-like receptors are key participants in innate immune responses. Biol Res, 2007. 40(2): p. 97-112.. 10.. Beutler, B.A., TLRs and innate immunity. Blood, 2009. 113(7): p. 31 . .
(42) . 1399-407. 11.. Alexopoulou, L., et al., Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature, 2001. 413(6857): p. 732-8.. 12.. Yamamoto, M., et al., Cutting Edge: A Novel Toll/IL-1 Receptor Domain-Containing Adapter That Preferentially Activates the IFN-{beta} Promoter in the Toll-Like Receptor Signaling. J Immunol, 2002. 169(12): p. 6668-6672.. 13.. Yamamoto,. M.,. et. al.,. Role. of. adaptor. TRIF. in. the. MyD88-independent toll-like receptor signaling pathway. Science, 2003. 301(5633): p. 640-3. 14.. Uematsu, S. and S. Akira, Toll-like receptors and Type I interferons. J Biol Chem, 2007. 282(21): p. 15319-23.. 15.. Martin, G.S., The hunting of the Src. Nat Rev Mol Cell Biol, 2001. 2(6): p. 467-75.. 16.. Meng, F. and C.A. Lowell, Lipopolysaccharide (LPS)-induced macrophage activation and signal transduction in the absence of Src-family kinases Hck, Fgr, and Lyn. J Exp Med, 1997. 185(9): p. 1661-70.. 17.. Leu, T.H., et al., Lipopolysaccharide-induced c-Src expression plays a role in nitric oxide and TNFalpha secretion in macrophages. Mol Immunol, 2006. 43(4): p. 308-16.. 18.. Maa, M.C., et al., Requirement of inducible nitric-oxide synthase in lipopolysaccharide-mediated Src induction and macrophage migration. J Biol Chem, 2008. 283(46): p. 31408-16. 32 . .
(43) . 19.. Tyryshkin, A., et al., Src kinase-mediated phosphorylation stabilizes inducible nitric-oxide synthase in normal cells and cancer cells. J Biol Chem. 285(1): p. 784-92.. 20.. Doyle, S.E., et al., Toll-like receptor 3 mediates a more potent antiviral response than Toll-like receptor 4. J Immunol, 2003. 170(7): p. 3565-71.. 21.. Johnsen, I.B., et al., Toll-like receptor 3 associates with c-Src tyrosine kinase on endosomes to initiate antiviral signaling. EMBO J, 2006. 25(14): p. 3335-46.. 22.. Wilson, L.K., et al., pp60c-src tyrosine kinase, myristylation, and modulatory domains are required for enhanced mitogenic responsiveness to epidermal growth factor seen in cells overexpressing c-src. Mol Cell Biol, 1989. 9(4): p. 1536-44.. 23.. Balachandran, S., E. Thomas, and G.N. Barber, A FADD-dependent innate immune mechanism in mammalian cells. Nature, 2004. 432(7015): p. 401-5.. 24.. Weinstein, S.L., M.R. Gold, and A.L. DeFranco, Bacterial lipopolysaccharide stimulates protein tyrosine phosphorylation in macrophages. Proc Natl Acad Sci U S A, 1991. 88(10): p. 4148-52.. 25.. English, B.K., et al., Hck tyrosine kinase activity modulates tumor necrosis factor production by murine macrophages. J Exp Med, 1993. 178(3): p. 1017-22.. 26.. Stefanova, I., et al., Lipopolysaccharide induces activation of CD14-associated protein tyrosine kinase p53/56lyn. J Biol Chem, 1993. 268(28): p. 20725-8. 33 . .
(44) . 27.. Lu, G., et al., ISG15 enhances the innate antiviral response by inhibition of IRF-3 degradation. Cell Mol Biol (Noisy-le-grand), 2006. 52(1): p. 29-41.. 28.. Zhao, C., et al., Human ISG15 conjugation targets both IFN-induced and constitutively expressed proteins functioning in diverse cellular pathways. Proc Natl Acad Sci U S A, 2005. 102(29): p. 10200-5.. 29.. Turan, K., et al., Nuclear MxA proteins form a complex with influenza virus NP and inhibit the transcription of the engineered influenza virus genome. Nucleic Acids Res, 2004. 32(2): p. 643-52.. 30.. Malathi, K., et al., Small self-RNA generated by RNase L amplifies antiviral innate immunity. Nature, 2007. 448(7155): p. 816-9.. 31.. Dar, A.C., T.E. Dever, and F. Sicheri, Higher-order substrate recognition of eIF2alpha by the RNA-dependent protein kinase PKR. Cell, 2005. 122(6): p. 887-900.. 32.. Torchinsky, M.B., et al., Innate immune recognition of infected apoptotic cells directs T(H)17 cell differentiation. Nature, 2009. 458(7234): p. 78-82.. 33.. Marshall-Clarke, S., et al., Polyinosinic acid is a ligand for toll-like receptor 3. J Biol Chem, 2007. 282(34): p. 24759-66.. 34.. Kariko, K., et al., mRNA is an endogenous ligand for Toll-like receptor 3. J Biol Chem, 2004. 279(13): p. 12542-50.. 35.. Oshiumi, H., et al., TICAM-1, an adaptor molecule that participates in Toll-like receptor 3-mediated interferon-beta induction. Nat Immunol, 2003. 4(2): p. 161-7. 34 . .
(45) . 36.. Gilliet, M., W. Cao, and Y.J. Liu, Plasmacytoid dendritic cells: sensing nucleic acids in viral infection and autoimmune diseases. Nat Rev Immunol, 2008. 8(8): p. 594-606.. 37.. Tomkowicz, B., et al., The Src kinase Lyn is required for CCR5 signaling in response to MIP-1beta and R5 HIV-1 gp120 in human macrophages. Blood, 2006. 108(4): p. 1145-50.. 38.. Freudenburg, W., M. Buller, and J.A. Corbett, Src Family Kinases Participate. in. the. Regulation. of. Encephalomyocarditis. Virus-Induced Cyclooxygenase-2 Expression by Macrophages. J Gen Virol. 39.. Hirasawa, K., et. al., Prevention of encephalomyocarditis. virus-induced diabetes in mice by inhibition of the tyrosine kinase signalling pathway and subsequent suppression of nitric oxide production in macrophages. J Virol, 1999. 73(10): p. 8541-8. 40.. Goody, R.J., C.C. Hoyt, and K.L. Tyler, Reovirus infection of the CNS enhances iNOS expression in areas of virus-induced injury. Exp Neurol, 2005. 195(2): p. 379-90.. 41.. Croen, K.D., Evidence for antiviral effect of nitric oxide. Inhibition of herpes simplex virus type 1 replication. J Clin Invest, 1993. 91(6): p. 2446-52.. 42.. Zaragoza, C., et al., Nitric oxide inhibition of coxsackievirus replication in vitro. J Clin Invest, 1997. 100(7): p. 1760-7.. 43.. Jacobs, A.T. and L.J. Ignarro, Lipopolysaccharide-induced expression of interferon-beta mediates the timing of inducible nitric-oxide synthase induction in RAW 264.7 macrophages. J Biol 35 . .
(46) . Chem, 2001. 276(51): p. 47950-7.. 36 .
(47) . Figure. Figure 1. Poly(I:C) treatment induces IFN-β production in murine RAW264.7 macrophages.. RAW264.7 cells were treated with poly(I:C) (20 μg/ml) for various times points as indicated. The concentration of IFN-β in culture medium of each sample was determined by ELISA.. 37 .
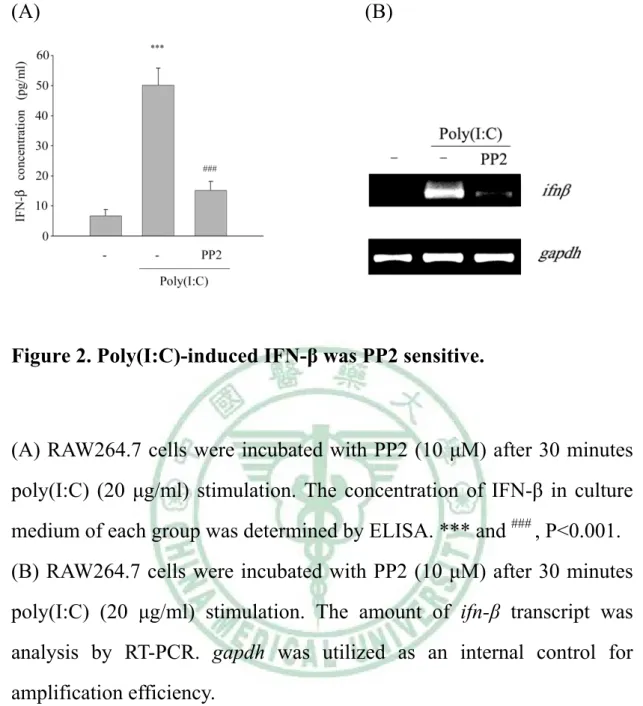
(48) . (A). (B). Figure 2. Poly(I:C)-induced IFN-β was PP2 sensitive.. (A) RAW264.7 cells were incubated with PP2 (10 μM) after 30 minutes poly(I:C) (20 μg/ml) stimulation. The concentration of IFN-β in culture medium of each group was determined by ELISA. *** and ### , P<0.001. (B) RAW264.7 cells were incubated with PP2 (10 μM) after 30 minutes poly(I:C) (20 μg/ml) stimulation. The amount of ifn-β transcript was analysis by RT-PCR. gapdh was utilized as an internal control for amplification efficiency.. 38 .
(49) . Figure 3. Poly(I:C)-mediated IFN-β secretion was inhibited by src-specific siRNA, which could be reversed by ectopic Src.. RAW264.7 cell and its derived control (ctrl-1, -2), Src attenuated cells (siRNA-1, -2) and Src attenuated cells harboring plasmid encoding avian Src (siRNA/Src6, siRNA/Src15) were stimulated without or with poly(I:C) (20 μg/ml) for 48 hours. Equal amounts of lysates (80 μg) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated. The concentration of IFN-β in culture medium of each group was determined by ELISA. The arrow indicates the position of Src. ***, P<0.001.. 39 .
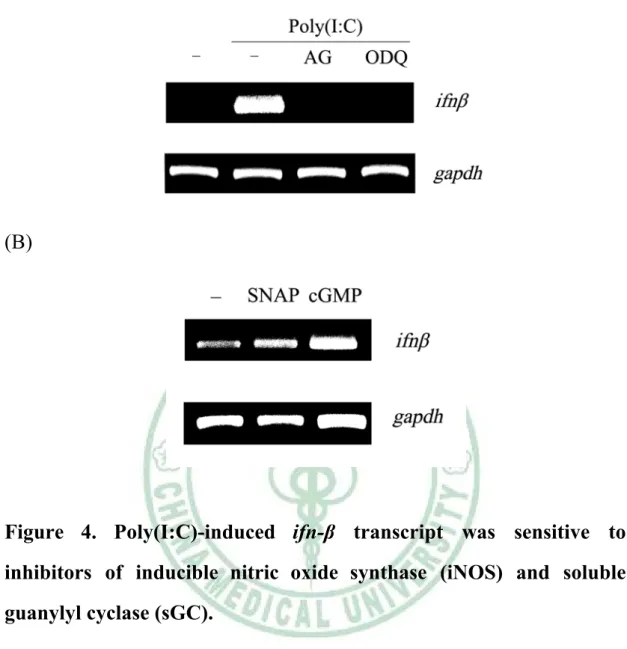
(50) . (A). (B). Figure 4. Poly(I:C)-induced ifn-β transcript was sensitive to inhibitors of inducible nitric oxide synthase (iNOS) and soluble guanylyl cyclase (sGC).. (A) RAW264.7 cells were pretreated without or with AG (2 mM), ODQ (100 μM) for 30 mins , and then cells were stimulated without and with poly(I:C). (B) RAW264.7 cells were stimulated without and with SNAP (100 μM), 8-br-cGMP (cGMP analogue, 100 μM) for 48 hours. The amount of ifn-β transcript was analysis by RT-PCR. gapdh was utilized as an internal control for amplification efficiency.. 40 .
(51) . Figure 5. iNOS was involved in poly(I:C)-mediated IFN-β production.. Primary peritoneal macrophages from wild type (WT) and iNOS-/- mice were treated without and with poly(I:C) (20 μg/ml) for 48 hours. The concentration of IFN-β in culture medium of each sample was determined by ELISA. ***, P<0.001.. 41 .
(52) . Figure 6. SNAP-mediated IFN-β secretion was inhibited by src-specific siRNA, which could be reversed by ectopic Src.. RAW264.7 cell and its derived control (ctrl-1, -2), Src attenuated cells (siRNA-1, -2) and Src attenuated cells harboring plasmid encoding avian Src (siRNA/Src6, siRNA/Src15) were stimulated without or with SNAP for 48 hours. Equal amounts of lysates (80 μg) from each sample were resolved by SDS-PAGE and probed with antibodies as indicated. The concentration of IFN-β in culture medium of each group was determined by ELISA. The arrow indicates the position of Src. ***, P<0.001.. 42 .
(53) . (A). (B). (C). Figure 7. Poly(I:C)-mediated IRF7 nuclear translocation was inhibited by src-specific siRNA, which could be reversed by ectopic Src.. (A) RAW 264.7 cells were incubated with PP2 (10 μM) after 30 minutes poly(I:C) (20 μg/ml) stimulation. The amount of nuclear IRF3 and IRF7 were analysis by nuclear extraction. (B)(C)The RAW264.7 expressing nonspecific siRNA (Ctrl-1), Src attenuated cells (siRNA-1) and Src attenuated cells harboring plasmid encoding avian Src (siRNA/Src6, siRNA/Src15) were stimulated without or with poly(I:C) (20 μg/ml) for 48 hours and 12 hours. The amount of nuclear IRF3 and IRF7 were analysis by nuclear extraction.. 43 .
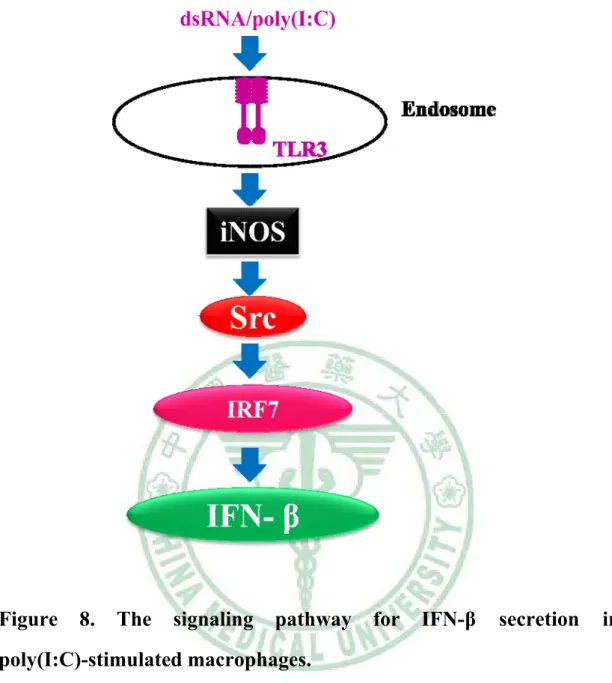
(54) . Figure. 8.. The. signaling. pathway. for. IFN-β. secretion. in. poly(I:C)-stimulated macrophages.. Following poly(I:C) treatment, the signal is transduced into macrophages via Toll-like receptor 3, increases expression of iNOS. NO produced by iNOS contribures to Src induction. And Src activation effect the amount of nuclear IRF7 cause IFN-β production.. 44 .
(55) . Appendix. Supplementary Figure 1. The inhibitory effect of PP2 on poly(I:C)-induced protein tyrosine phosphorylation. (provide for 妙 瑩學姐和青昭學姐). 45 .
(56) . Supplementary Figure 2. iNOS is involved in poly(I:C)-mediated macrophage migration. (provide for 妙瑩學姐和青昭學姐). 46 .
(57) . Supplementary Figure 3. Poly(I:C) induced Src expression in murine RAW264.7 macrophages and rat peritoneal macrophages (PEMs). (provide for 妙瑩學姐和青昭學姐). 47 .
(58) . Supplementary Figure 4. Poly(I:C) induced Src expression is dependent on iNOS. (provide for 妙瑩學姐和青昭學姐). 48 .
(59)
數據
+4
相關文件
Combines DNA biorepositories with electronic medical record (E MR) systems for large-scale, high-throughput genetic research wi th the ultimate goal of returning genomic
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
Mie–Gr¨uneisen equa- tion of state (1), we want to use an Eulerian formulation of the equations as in the form described in (2), and to employ a state-of-the-art shock capturing
The notation T n (x) is used to represent the nth partial sum of this series and we can call it as it the nth-degree Taylor polynomial of f at a... In general, it can be shown
To define surface integrals of vector fields, we need to rule out nonorientable surfaces such as the Möbius strip shown in Figure
Notice that if the dx in the notation for an integral were to be interpreted as a differential, then the differential 2xdx would occur in (1) and so, formally, without justifying our
Then we can draw a right triangle with angle θ as in Figure 3 and deduce from the Pythagorean Theorem that the third side has length.. This enables us to read from the
As discussed in the example (Figure 3.3), the goal of the min-sum algorithm is to find a valid configuration such that the sum of the local costs (over all sites and check sets) is
