行政院國家科學委員會專題研究計畫 期中進度報告
德國蜚蠊生物時鐘掩蓋機制探討(2/3)
計畫類別: 個別型計畫 計畫編號: NSC91-2313-B-002-296- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立臺灣大學昆蟲學系暨研究所 計畫主持人: 李後晶 報告類型: 精簡報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 92 年 6 月 2 日
行政院國家科學委員會專題研究計畫進度報告
德國蜚蠊生物時鐘掩蓋機制探討
主持人:李後晶 國立臺灣大學昆蟲學系
背景及目的:
德國蜚蠊生物時鐘掩蓋機制探討
地球因自轉而產生了二十四小時的日夜變化及季節變換,而生命最重要的任 務就是適應環境以產生後代,因此,在生命進化歷史中,生物體表現出日週律動 (circadian rhythm)現象。日週律動,一個接近二十四小時之週期律動,是具有適 應上的意義。這或許可以解釋為什麼從細菌到高等動物,不管在生化上、生理上, 乃至於行為上都有日週律動的表現。 基因表現、生理週期性變動與行為表現上的日週律動,都是由生物個體內的 調律器(pacemaker)所控制,也由於調律器屬於細胞層次,因此也被稱為時鐘細胞 (clock cell)。生物時鐘運作系統主要有三個組成(Takahashi, 1995):(一)調律器 (pacemaker),是產生時間訊息之所在。(二)環境訊息輸入路徑(input pathway),負 責傳遞外界環境訊息至調律器中樞,以使調律器能與外界環境時間同步。(三)時 間訊息輸出路徑(output pathway) ,傳遞調律器所生之時間訊息至下游,在生化 上、生理上、行為上表現出日週律動現象。目前,在生物時鐘運作機制研究的領 域裡,已建立出調律器中樞時間訊息產生的分子機制。整個日週律動系統是由自我調節的迴饋循環(autoregulatory feedback loop)所控制(Dunlap, 1999)。黃果蠅
(Drosophila melanogaster) 的 分 子 時 鐘 負 迴 饋 機 制 , 是 由 dCLOCK/CYCLE
per 與 tim 基因,表現出 PER 與 TIM 蛋白質於細胞質(Darlington et al.,1998;
Hogenesch et al., 1998)。當細胞質內累積足夠量的 PER 與 TIM 蛋白質,會形成
PER/TIM heterodimer,進入細胞核內抑制 dCLK/CYC heterodimer 的活性,關閉
per 與 tim 基因的表現(Lee et al., 1999)。除了 per-tim 的負迴饋循環,另一個控制
dCLK 於相反相位變動的循環,則是受到 dCLK/CYC heterodimer 的抑制,與
PER/TIM heterodimer 的活化(Glossop et al., 1999)。
生物時鐘基因 per 已經陸續從許多昆蟲選殖出來。第一個生物時鐘基因 per,由
Konopka 及 Benzer 於 197l 年所發表,他們以黃果蠅(Drosophila melanogaster)羽
化為日週律動之觀察指標,找到了 per 基因的突變型。從此以後,開始生物時鐘
分子機制研究的新紀元。per 基因在許多種果蠅都有發現,而且胺基酸序列有相
似的保守性(Colot et al., 1988)。在日夜二十四小時交替下的 LD 環境中 perRNA
及 PER 蛋白質表現量呈穩定約二十四小時週期,而最大的表現量都在夜晚。在
全暗 DD 的環境下,也就是沒有外界環境時間訊息的情況下,per RNA 及 PER
蛋白質表現量呈現一個週期接近二十四小時的自由律動(free-running)。這種現象
也與行為觀測相符。
時鐘細胞位於生物體內的何處,也是生物時鐘研究的基本課題。早期的研究
主要是破壞細胞組織,觀察行為上或是生理上的日週律動指標,以確定生物體內
的時鐘細胞,或是調律器的所在位置。近十年來由於分子生物技術上的進步,如
今一般的做法是以原位雜合反應(in situ hybridization),找尋哪些細胞或是組織表
現調律器的中樞分子如 per RNA 或 PER 蛋白質,來確定調律器的所在位置;此
外,因為 PER 並不是一個會釋出細胞外的蛋白質,使用 PER 蛋白質的抗體來
作免疫染色,也是一個方法。這比起傳統的破壞方法更為精確與直接。目前已知
高等動物老鼠的主要調律器位置是在 SCN(suprachiasmatic nuclei)。鳥類的主要調
律器是位於 pineal gland(Sassone-Corsi, 1998)。天蠶蛾(Antheraea pernyii)左右半腦
各有四個神經細胞擔負調律器的工作(Sauman and Repper, l996)。在黃果蠅腦部則
1988)。Plautz 等人於 1997 年證明在黃果蠅的全身都具有時鐘細胞的存在。這些
時鐘細胞分別擔負不同的功用,大部分的時鐘細胞是屬於區域性的時鐘細胞,它
們所產生的時間訊息只提供給自己或是周圍鄰近的細胞或組織時間訊息,使得區
域性的細胞運作能在時間上適應外在環境的變化。
德國蜚蠊雌性成蟲的卵巢在卵鞘形成過程中,會抑制雌性成蟲活動行為的日
週律動。但是卵巢摘除後的雌成蟲則恢復具有日週律動的活動行為(Lin and Lee,
1998)。關於卵巢掩蓋活動行為日週律動現象的機制並不十分清楚。掩蓋的機制 有兩種可能性:(一)卵巢存在擾亂了調律器的運作,故活動行為不具日週律動現 象。(二)卵巢存在阻礙訊息輸出路徑,故活動行為不具日週律動現象。藉由研究 日週律動被掩蓋的現象,有助於我們找出正常的訊息輸出路徑,進一步建立訊息 輸出路徑的可能機制。 要解答以上的問題可以 per 基因表現情形作為分子指標,常用的方式是用北
方墨點法(northern blotting)偵測 RNA 的表現,或是用西方墨點法(western blotting)
偵測蛋白質的表現。當然也可藉由找尋其他生化或是生理指標進行偵測判斷,但 是卻沒有比偵測 per 基因表現情形來得直接.並對訊息輸出路徑的研究來得有貢 獻。因此,我們預定的做法是利用已經找出的德國蜚蠊 per 基因,製作核酸探針, 偵測具卵巢的德國蜚蠊雌性成蟲的 perRNA 表現。如果 perRNA 的表現在有卵巢 的情況之下仍具日週律動現象,則我們可確定調律器本身並未被干擾,卵巢的掩 蓋作用是在訊息輸出的路徑上。如果 perRNA 的表現在有卵巢的情況之下,沒有 日週律動的現象.則我們可以確定卵巢掩蓋作用直接作用在調律器中樞的分子系 統,或是作用在中樞分子系統的周圍。
1,1’-dihexadecyl-3,3,3’,3’-tetramethylindocarbocyanine perchlorate (DiIC16) 是
一種長碳鏈的螢光染劑,能夠嵌入細胞膜中,隨著膜上分子的流動而擴散,從而
可 由細胞 本 體追 蹤其神 經 軸突和 樹 突之分布 (Honig and Hume, 1989; von
Barthrls et al., 1990)。比起過往的鈷離子 (Koontz and Edward, 1980) 或者鎳離子
具有「不影響細胞的生理」、「不外洩到其他細胞,不會造成污染」、「螢光不易衰
減,有利長期觀察」等優點 (Honig and Hume, 1989; von Barthrls et al., 1990)。
鑑定出雄性德國蜚蠊節律器細胞的確實位置之後,如果這些細胞的分布非常 接近、群聚成團,我們便可使用 DiI 來追蹤這些節律器細胞的樹突和軸突,以 建立日週律動調控的神經網絡。如果這些時鐘細胞是分散的,則可針對所得的細 胞位置進行「神經元移除」的設計,分別將各個細胞殺死、或者切斷其調控路徑, 然後觀察蟑螂個體的行為反應,藉以判斷各個細胞在日週律動行為調控之中扮演 的角色。此時因已知各個細胞的位置,所進行移除法的精準度將較先前更為提高。 為了提高判斷活體內時鐘細胞位置的精確度,亦可配合電神經生理之 intracellular recording 方法,篩除不可能屬於節律器的細胞。如果在活體內判斷 時鐘細胞位置實在過於困難,亦可反過來,在德國蜚蠊複眼位置放置 DiI 溶液 或者結晶,由神經末梢追逆至神經本體,再將染出的路徑與原位雜合反應所染出 的細胞進行比較、篩除,即可建立時鐘細胞在腦內的神經網路。 與其他蜚蠊一樣,德國蜚蠊的活動行為調律器位於視葉(Helfrich-Forster et al.
1998; Wen and Lee, 2000)。因為德國蜚蠊具有獨特的掩蓋現象,所以探討掩蓋作
用發生的標的,將有助於我們更進一步瞭解日週律動系統。假設控制活動行為調
律器是主要的調律器所在,如果雌性成蟲的活動行為,不表現日週律動,而其他
生理反應表現出明顯的日週律動,我們就可以判斷掩蓋因子不是作用再調律器本
身,而是作用於日週律動系統的輸出路徑。
視網膜神經圖譜(elctroretinogram (ERG))已經被證實是測量視覺感受性很有
效的方法,已有許多昆蟲利用 ERG 來測量視覺感受性的日週律動(Koehler and
Fleissner, 1978)。在馬得拉蜚蠊(Leucophaea maderae)控制活動行為日週律動的調
律器與控制 ERG 日週律動的調律器都是位於視葉中(Will et al. 1985)。所以 ERG
是一種很好的測量生理反應日週律動的指標。此計畫利用比較活動行為與視覺感
受性兩者的的日週律動系統,進一步瞭解德國蜚蠊的日週律動系統的調控。尤其
卵巢發育之掩蓋機制是否位於調律器上或時間訊息的輸出路徑上。
在相同的亮暗週期下,德國蜚蠊雌性成蟲的活動行為和雄性成蟲完全不
同;雌性成蟲不表現日週律動的活動行為,日活動量呈現與生殖週期相配合的週
期性 (Lee and Wu, 1994)。然而,經過切除卵巢手術之後,雌蟲便可表現出與雄
蟲完全相同的活動模式:在全暗環境下表現出自由律動;在亮暗週期下表現出日
週律動,日週期長度與雄蟲的沒有顯著差異,活動高峰也與雄蟲相同 (Lin and
Lee, 1996)。由此可知,雌蟲的活動行為仍然受到節律器的控制,但卵巢會對日
週律動產生掩蓋作用。進行切斷腹神經索手術之後,卵巢的訊息便無法經過神經
網路傳遞;但是此項掩蓋作用仍然不會消失。可見卵巢對於雌蟲日週律動的掩蓋
作用並非透過神經傳遞,而可能是經由荷爾蒙路徑調控 (Lin and Lee, 1998)。然
而到目前為止,尚未有任何文獻提出掩蓋作用的確實機制。我們懷疑造成掩蓋作
用的調控荷爾蒙乃是蜚蠊卵巢主要分泌的荷爾蒙 ecdysone,因此將進行以下兩
種實驗:(一)我們計畫以以下二法之一來了解 ecdysone 在雌性蜚蠊體內的主
要作用位置,並建立日週律調控傳導的路徑: 1. 以 ecdysone receptor 的抗體進
行免疫染色 (King-Jones, et al., 1999)、2. 以 ecdysone receptor 作為標的之原位雜
合反應 (Jindra et al., 1996; Crispi et al., 1998; Stocker et al., 1997; Draizen et al.,
1999; Lezzi et al., 1999)。(二)如果實驗一順利被完成,我們將以注射、飲食或 者在飼育環境中塗佈等方式,對雄性德國蜚蠊投以 ecdysone,然後觀察其活動 行為;如果 ecdysone 可以對雄性成蟲造成類似於雌性成蟲卵巢的掩蓋作用,則 假說「ecdysone 是掩蓋作用的調控荷爾蒙」便得到有力的佐證(但反之不然。 因為如果 ecdysone 無法對雄性成蟲造成類似於雌性成蟲卵巢的掩蓋作用,可能 是因為雄性成蟲 ecdysone receptor 的位置或者出現時機不利於產生掩蓋作用。 為了避免此點,在進行實驗二之前,可以先同樣對雄性成蟲進行 ecdysone receptor 的免疫染色。)
(L.)
It is an essential for a circadian clock to express a free-running rhythm of
species-specific period when the organism is under a constant environment after
entrainment (Brady, 1982). This phenomenon is found in molecular and cellular,
organismic and colonial levels (Lin et al., 2002; Lee and Loher, 1993; Moore, 2001).
However, the free-running period may change when this individual has been
experienced more than one Zeitgebers. The term “after-effects” has been toyed
around for some time (Pittendrigh and Daan, 1976), but not many chronobiologists
put effects in this aspect of circadian study. Until recently, Page and his associates
have revealed history-dependent effects on circadian periods of cockroach and other
organism (Page et al., 1999, 2001). Their contribution is important in the clock
entrainment and the development of the circadian system from embryo to adults.
The German cockroach, Blattella germanica (L.), is a cosmopolitan household
species (Koehler et al., 1999). Although several aspects of reproduction and
locomotion have been investigated in this nuisance insect, not much information has
been obtained about its locomotor pattern and environmental effects on its circadian
clock adjustment. We have established the relationship between locomotor pattern
and its reproductive functions (Lee and Wu, 1994; Lin and Lee, 1996; Tsai and Lee,
pattern (Lin and Lee, 1996; Wen and Lee, 2000). In this study, we investigate the
clock adjustment after two different Zeitgebers in sequence. The daily locomotor
pattern is analyzed to unveil the after-effects of its previous experience under various
photoperiod conditions. The function of this phenomenon and the standardization of
the subjective night of various photoperiods will be discussed in this report.
材料和方法:
供試蟲源及飼養方法
自台北市區採得德國蜚蠊(Blattella germanica (L.)),置於 28C、光週期為 16 小時光照、8 小時黑暗(16L:8D)的恆溫生長箱中,以統一寶多福特幼犬飼料及 水,累代飼養數十代之後,始為試驗之用。若蟲以群聚的方式飼養於二公升的透 明塑膠桶中,桶蓋上挖出一個直徑 6 公分的洞,以不銹鋼網覆蓋以為透氣之用;桶壁上塗上 FLUON (Northern Product, Inc.),避免昆蟲逃逸;桶中放置數塊保麗
龍,增加蜚蠊攀爬躲藏的空間。俟發育至末齡若蟲,再移到 250ml 的透明塑膠杯 (直徑 9.5 公分,高 5.5 公分,盒蓋有 3*3 平方公分之紗網)中飼育。每日將羽化 的成蟲挑出,雌、雄蟲分開飼養,以備試驗之用。成蟲日齡計算的方式,以羽化 24 小時以內者為 0 日齡,24 小時至 48 小時者為 1 日齡,餘者類推。
以 PER 蛋白質的免疫染色找出日週律動調律器的所在位置
冷凍切片 在解剖顯微鏡下,將德國蜚蠊第 5 日齡雄性成蟲的大腦完整解剖出 來,以 4% paraformaldehyde (in PBS) 固定 2 小時,然後洗滌 5 次,分別為 5、10、20、20、20 分鐘,徹底洗淨以以避免固定劑干擾溫度下降。以切片膠包埋 之後,用冷凍切片機切成連續薄片,平整黏附在載玻片上 [載玻片必須事先擦拭 乾淨,並且塗敷上一層極薄的蛋白膠 (蛋白加入等體積甘油,用力攪打使空氣充 分進入,然後靜置一夜,取其下清液)]。再用 4% paraformaldehyde 後固定 20 分 鐘,用 0.1 % PBT 洗滌 3 次,每次 5 分鐘,充分洗淨以避免固定劑干擾染色 結果。然後使用無色指甲油將切片周圍框住,以避免染劑在長時間的染色之中流
失。在 wet chamber 中,以 3 % BSA in 0.1 % PBT 進行 blocking 4 小時;之後
在切片上注以一級抗體溶液 (1/200 anti-PER, 3 % BSA in 0.1 % PBT) 染色 2 小時。以 3 % BSA in 0.1 % PBT 將非專一性附著的一級抗體徹底洗淨,然後在 切片上注以二級抗體溶液 (1/1000 anti-rabbit, 3 % BSA in 0.1 % PBT),2 小時後 洗滌乾淨。所使用的的二級抗體有兩種,一種接上 AP,一種接上 peroxidase; 分別以 AP 和 PO 相應的呈色劑呈色。使用鑷子揭起無色指甲油圍邊,在光學 顯微鏡下觀察並予記錄。 Whole mount 在解剖顯微鏡下,將德國蜚蠊第 5 日齡雄性成蟲的大腦完整解剖出 來,以 4% paraformaldehyde (in PBS) 固定 2 小時,然後洗滌 5 次,分別為 5、 10、20、20、20 分鐘,徹底洗淨以以避免固定劑干擾染色結果。以 3 % BSA in 0.1 % PBT 培養 16 小時以增加通透性並進行 blocking。之後以一級抗體溶液
(1/200 anti-PER, 3 % BSA in 0.1 % PBT) 培養 24 小時,以 3 % BSA in 0.1
% PBT 將非專一性附著的一級抗體徹底洗淨。然後以二級抗體溶液 (1/1000
anti-rabbit, 3 % BSA in 0.1 % PBT) 培養 12 小時,洗滌乾淨。所使用的的二級抗
體有兩種,一種接上 AP,一種接上 peroxidase;分別以 AP 和 PO 相應的呈
色劑呈色。以 1 M Meglumine diatrizoate (Sigma chemical co., St. Louis) 使組織透
Locomotor activity
Daily locomotion of a male cockroach was continuously monitored and
recorded by an infrared motion detector system. The system has been described in a
previous report (Lin and Lee, 1998). Unless mentioned otherwise, insects reared in
28℃, LD 16:8 h and LD 12:12 h were entrained at 28℃, LD 16:8 h and LD 12:12 h
for 5 days (the first LD condition), respectively, and then switched to constant
darkness (DD) condition for another11 days. After DD period, the cockroaches were
re-entrained to the different LD conditions (LD 12:12h and LD 16:8h, respectively)
for 5 days (the second LD condition), and then switched to constant darkness (DD)
condition again for 12 days. The cool fluorescent light (285 lux) was used to entrain
the cockroaches.
Data analysis
The rhythmicity of their daily locomotion and circadian period (τ) were
calculated and analyzed for the data obtained during 10 consecutive days by using a
chi-square periodogram at 0.05 significant level (Sokolove and Bushell, 1978).
Circadian time was expressed as degree, with 360° as one full cycle of a rhythm.
Since the German cockroach is a nocturnal animal, Zeitgeber time 0 was defined as
the time of light off under light-dark conditions. The circadian time 0 was following
the original Zeitgeber time 0 and calculated according to the circadian period decided
by the free-running rhythm under DD condition. In order to eliminate individual
within its own circadian period.
結果:
Whole mount
無論使用接有 AP (圖 1) 還是 peroxidase (圖 2) 的二級抗體,德國蜚蠊 視葉的細胞都呈現相當均一的染色結果,並未見到特定被標定上的目標細胞。冷凍切片
無論使用接有 AP (圖 3) 還是 peroxidase (圖 4) 的二級抗體,德國蜚蠊 圖 1. 第 5 日齡雄性德國蜚蠊 [Blattella germanica (L.)] 成蟲的大腦視葉,以 anti-PER 一級抗體、接上 AP 之二級抗體進行免疫染色之結果。上方弧形深色 者為複眼之色素。比例尺:100 μm。視葉的細胞都呈現相當均一的染色結果,並未見到特定被標定上的目標細胞。
(A)
圖 2. 第 5 日齡雄性德國蜚蠊成蟲的大腦視葉,以 anti-PER 一級抗體、接上 peroxidase 之二級抗體進行免疫染色之結果。上方弧形深色者為複眼之色素。比 例尺:100 μm。
Daily locomotor pattern under photic entrainment
The daily locomotor patterns of male adults under 28℃, and LD 16:8h or
12:12h conditions for 16 consecutive days are shown in Fig. 5. Under either light
regimes, the male adults did express a clear entrainment. Although the lighting
hours per day were different, the daily locomotion pattern remained the same.
Locomotor activity of Blattella germanica was restricted to the scotophase.
Regardless of different lighting hours per day, the daily locomotion was the same for
both light regimes.
Under photic entrainment condition, the German cockroach expressed a
typical nocturnal-active locomotion. Its locomotion was confined in scotophase, but
that in 12 hours of darkness (Table1). The distribution of locomotion during entire
Zeitgeber period under LD 16:8 was shown in Fig. 6A (n=16). Since a day was
divided into 100 time fragments, each 3.6° should have 1% of activities when the
locomotion was equally distributed. However, when light was turned off, the
activity of males increased sharply. At the second time fragment (3.6~7.2°), they
reached the highest activity of 6.6% in total locomotion of a day. The majority of
locomotor activities were concentrated in the scotophase. From Zeitgeber time
118.8 to 122.4°, the activity suddenly increased over 1% (t-test, p<0.05), when the
light was turned on as the beginning of the photophase. This short burst of
locomotion might be the subtle transition effect of lighting. From 122.4 to 356.4°,
each time fragment contained less than 1% of activities (t-test, p<0.05). This result
showed the inhibition effect of lighting on the locomotion. Right before the onset of
scotophase (356.4 to 360°), the activities started to build up over 1% (t-test, p>0.05).
The similar distribution pattern of locomotion was also found in the photic
entrainment LD12: 12h (Fig. 2B, n=7). The surged activity before photophase in LD
16:8h, however, did not show in this photoperiod condition. This finding confirmed
the possibility of stumble effect by lighting. In addition, in the anticipation of
scotophase, the German cockroach started to become active in locomotion which
and 6B). In order to compare the locomotion distribution under two different photic
entrainment conditions, the daily accumulated activities through 24 hours under LD
16:8h and 12:12h conditions were shown in Fig. 6C. Although majority of activities
confined in the scotophase were different (83% and 89.2% of daily activities for LD
16:8h and LD 12:12h, respectively), the velocity of activity increasing exponentially
under both conditions was the same in which showed the fastest in the beginning of
scotophase and the slowest at starting time of photophase. Eighty percent of the
accumulated locomotion occurred before Zeitgeber time 120°. Regardless the length
of scotophase, the locomotion distribution of locomotion was the same (t-test,
p>0.05).
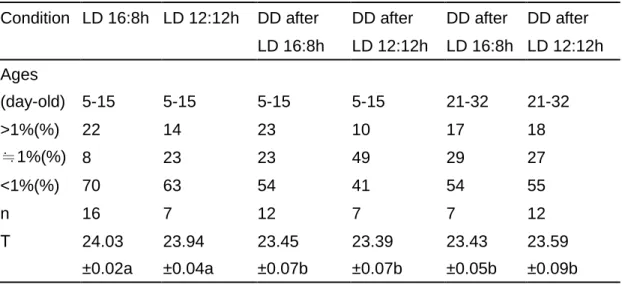
Table 1. Locomotor distribution and circadian period under different light-dark condition
Condition LD 16:8h LD 12:12h DD after DD after DD after DD after
LD 16:8h LD 12:12h LD 16:8h LD 12:12h Ages (day-old) 5-15 5-15 5-15 5-15 21-32 21-32 >1%(%) 22 14 23 10 17 18 ≒1%(%) 8 23 23 49 29 27 <1%(%) 70 63 54 41 54 55 n 16 7 12 7 7 12 Τ 24.03 ±0.02a 23.94 ±0.04a 23.45 ±0.07b 23.39 ±0.07b 23.43 ±0.05b 23.59 ±0.09b a Significant difference at p>0.01 between a and a.
b Significant difference at p>0.1 between b and b a, b Significant difference at p<0.01 between a and b.
(A) 0 6 12 18 24 0 5 10 15 (B) 0 6 12 18 24 0 5 10 15
Fig.5.Typical actograms of male Blattella germanica under 28℃ and two kinds of LD photoperiod conditions. (A) The newly emerged male cockroach from rearing under 28℃ and LD 16:8h conditions for the entire nymphal stages was placed under LD 16:8h cycle for 16 days. The shaded areas of the actogram represent periods of darkness. Each horizontal line show one day (24h), with each day attached below the previous day. Short vertical line indicates the locomotion. The superimposed line across the actogram represents the predicted onset time of the male’s locomotion according to its circadian period. The circadian period of the male was 24.0h. (B) The newly emerged male cockroach from rearing under 28℃ and LD 12:12h conditions for the entire nymphal stages was placed under LD 12:12h cycle for 16 days. The circadian period of the male was 24.0h.
After-effects on the free-running rhythm
The actograms of cockroaches that experienced two different LD cycles
consecutively and each followed by DD condition were shown in Fig. 7. The 5 days
of LD cycles clearly entrained the circadian clock, so that the phase shifting was swift
during the following DD conditions regardless the phase gap (4-8h). The different
photic entrainments did not cause the alteration of the circadian period under the
following DD conditions. The circadian period following LD 16:8 and LD 12:12h
was 23.3h and 23.2h, respectively (Fig. 7A). It showed the similar result for another
cockroach whose circadian period following LD 12:12h and LD 16:8h was 23.3h and
23.4h, respectively (Fig. 7B). However, the photic entrainment did cause some
changes on the distribution pattern of locomotion. After-effects did occur when
different Zeitgebers imposed on the same cockroach in sequence. The 12hrs of
scotophase after previous 16:8h caused activities more concentrated in DD than that
of cockroach without previous LD exposure (Fig. 7A). In addition, the 8hrs of
scotophase after pervious LD 12:12h made the activities more dispersed than that in
DD of cockroach without previous LD exposure (Fig. 7B). When the data were
grouped together, the after-effects were convincingly expressed in Table 1. Since the
photic entrainment cause the circadian clock to adjust its phase and period to
effects under DD conditions. Although the circadian periods were the same
following two different LD conditions (23.43±0.07h and 23.43±0.05h for the group of
cockroaches under LD 16:8 and 12:12h in sequence, and 23.39±0.07h and
23.59±0.09h for the after group under LD 12:12h and 16:8h in sequence, respectively),
the after-effects were represented by the changes of distribution pattern of locomotion
(Table 1). By increasing the percentage of locomotor time which was larger than 1%
from 10% in DD after LD 12:12 cycles to 17%, the activities showed more
concentrated than that of no after-effects, but not as concentrated as in DD after LD
16:8h. However, the activities became less concentrated from 23% to 18% of
locomotor time in DD after LD 16:8h.
For further analysis of after-effects on the free-running rhythm, the daily
locomotor distribution of a group of cockroaches in DD conditions after entraining by
LD 16:8h or LD 12:12h were shown in Fig. 4A and 4B, respectively. The most
active time (>1%, t-test, p<0.05) in DD after LD 16:8h were during circadian time
0~82.8° and 345.6~360° (Fig. 8A), which was different from circadian time
14.4~108° under DD condition after LD 12:12h (Fig. 8B). The peak time of
locomotion was also different which occurred at circadian time 356.4~360° and
14.4~18° in DD after LD 16:8h and LD 12:12h, respectively.
the distribution pattern of locomotion under DD condition after different LD
entrainment (Fig. 8C). The velocity of accumulative activities showed significant
increase in DD after LD 16:8h than that in DD after LD 12:12h before reaching
circadian time of 126° which was the beginning time of subjective day of DD after
LD 16:8h (t-test, p<0.05). The accumulated activity reached at 60% of the total
activities when the velocity under DD after LD 12:12h became quicker and leaded the
comparison until the end of circadian period (t-test, p<0.05). Although the
accumulated activities were the same in exponential distribution, the circadian
patterns did show little coincidence. However, this difference became the smallest
(35%) when the curve of accumulative activities in DD after LD 12:12h delayed 21.6°
(Fig. 8D). The 35% differences in accumulated activity pattern might represent the
effect of length differences in scotophase between LD 16:8h and LD 12:12h. Since
all the locomotor activities of the German cockroach were restricted in scotophase
under LD and in subjective night under DD, this result implied that the length of
subjective night was 120° and 180° in DD after LD 16:8h and LD 12:12h,
respectively.
When the same individuals experienced another Zeitgeber for 5 days after
free-running under DD, the locomotion under the following DD conditions expressed
occurred within circadian time 50.4~112.4° and peaked at 57.6~61.2° under DD after
LD 12:12h. But the distribution pattern under DD after LD 16:8h was different
which showed most active locomotion between circadian time 3.6~86.4° and peaked
at 21.6~25.2°. Since the circadian period of the same individuals remained the same
in the 2nd DD, the phase of the locomotion did delay about 68.4° in DD after LD
12:12h, but advance about 10.8° in DD after LD 16:8h. The accumulative activities
along the circadian time for the 2nd DD after LD 12:12h or LD 16:8h were shown in
Fig. 4G. The similar relationship between these two lines was found as the two
curves in 1st DD (Fig. 8C). The accumulated activities started to build up faster in
the early circadian time for the cockroaches who experienced LD 16:8h than that of
LD 12:12h. But the accumulated activities of the latter curve took over the lead at
the circadian time 190.8° till the end of circadian period. This switching point of
velocity was later than that of the curves in the 1st DD. But, the accumulated
activities at this point (80%) were higher than that in the 1st DD (60%). Although
these two curves showed little coincidence, they became inseparable (t-test, p>0.05)
when the phase of the latter curve (entrained by LD 12:12h) delayed 43.2° (Fig. 8H).
This phenomenon suggested the after-effects on the locomotor circadian rhythm
because the cockroaches experienced the same Zeitgebers but in different sequence.
A.
B.
C.
distribution pattern and phase when compared to the locomotion in the 1st DD.
However, the after-effects made the two curves in 2nd DD showed the same
distribution pattern of locomotion with only different in phase.
0 1 2 3 4 5 6 7 0 72 144 216 288 360 0 1 2 3 4 5 6 7 0 72 144 216 288 360 P ro p o rt io n a l a c ti v it y o f d a y (﹪ )
0 20 40 60 80 100 0 72 144 216 288 360 LD 16:8h LD 12:12h
Fig.6. The distribution of locomotion during entire circadian period under light-dark cycle. (A) Locomotor distribution of male cockroachs (5-15days-old) (n=16) under LD 16:8h cycle. X-axis is the phase angle of one rhythm. A rhythm was divided into 100 parts. Y-axis is proportional activity of a rhythm. Each bar was the mean of activities from 16 cockroaches and 10 circadian period cycles of everyone during 3.6°. The black bars indicate that the mean activities are significantly more then 1% (t-test, p<0.05). The deep gray bars indicate no significant difference between the mean activities with 1% (t-test, p>0.05). The light gray bars indicate that the mean activities are significantly less then 1% (t-test, p<0.05). (B) Locomotor distribution of male cockroachs (5-15days-old) (n=7) under LD 12:12h cycle. (C) The accumulative patterns of (6A) and (6B).
Accu mulat ive pro p ortio n al act ivity o f a p eriod ( ﹪) Z e i t g e b e r t i m e ( ° ) (A) 0 6 12 18 24 0 10 20 30 (B) 0 10 0 6 12 18 24
0 1 2 3 4 5 0 72 144 216 288 360 0 1 2 3 4 5 0 72 144 216 288 360 P ro p o rt io n a l a c ti v it y o f a p e ri o d ( ﹪)
B. C. D. F. G. H. 0 1 2 3 4 5 0 72 144 216 288 360 0 1 2 3 4 5 0 72 144 216 288 360 0 20 40 60 80 100 0 72 144 216 288 360 entrained by LD 16:8h entrained by LD 12:12h 0 20 40 60 80 100 0 72 144 216 288 360 entrained by LD 16:8h entrained by LD 12:12h 0 20 40 60 80 100 0 72 144 216 288 360 entrained by LD 16:8h entrained by LD 12:12h 0 20 40 60 80 100 0 72 144 216 288 360 entrained by LD 16:8h entrained by LD 12:12h Accu mulat ive pro p ortio n al a c tivity of a pe ri o d ( ﹪)
Fig.8. Analysis on the mechanical properties of locomotor free-running rhythm of the German cockroach under 28℃ and DD conditions. (A)&(E) The locomotor distribution of Fig.(7A) of the first (5-15 day-old) and second (21-32 day-old) DD conditions (n=12). (B)&(F) The locomotor distribution of Fig.(7B) of the first (5-15 day-old) and second (21-32 day-old) DD conditions (n=7). (C) The accumulative activities of (8A) and (8B). The solid and open circles represent accumulative activities of (8A) and (8B). The cockroaches were 5-15 day-old, but entrained by LD 16:8 h or 12:12 h cycles. The black ones are significant difference between each other (t-test, p<0.05) and the gray ones are not (p>0.05).
(G) The accumulative activities of (8E) and (8F). The solid and open circles represent accumulative activities of (8F) and (8E). The cockroaches were 21-32 day-old, but entrained by LD 16:8 h or 12:12 h cycles. (D) Fitting the accumulative patterns of (8A) and (8B). The distribution of (8B) was shifted and accumulated, and then compared with the fixing pattern of accumulative (8A). When the phase of (8B) was advanced 21.6°(6% of a period), the two patters of accumulative activities were the most similar to each other. (H) Fitting the accumulative patterns of (8E) and (8F). The distribution of (8E) was shifted and accumulated, and then compared with the fixing pattern of accumulative (8F). When the phase of (8E) was advanced 43.2°(12% of a period), the two patters of accumulative activities were the most similar to each other. The other symbols are described as in Fig.6.
討論:
德國蜚蠊生物時鐘掩蓋機制探討
目前所得結果和預期結果不同。預期希望能夠染出特定的 PER 蛋白質以標 定時鐘細胞,但目前所得染色結果各細胞並無差異。檢討其原因,可能因為實驗 動物為德國蜚蠊,而所採用的抗體所針對之抗原乃是小家鼠 (mouse) 的 PER 蛋 白質;兩種生物在類緣上相差太遠,以致抗體無法辨識德國匪蠊之 PER。 針對以上可能原因,接下來我們將採取以下策略來改進: 1. 直接以德國蜚蠊之 PER 為抗原,製作抗體。再以此抗體來進行 PER 蛋 白質的免疫染色。 2. 進行 per mRNA 的原位雜合反應。如此,即可避免物種類緣差異過大所造成的問題,亦可得到 per mRNA 層
次的資料,和 PER 蛋白質層次所得的資料互為印證。
After-effects on the Locomotor Circadian Rhythm of Blattella germanica
(L.)
Photoperiod affects many physiological and behavioral phenomena in insect.
In the wood roach, Ectobius lapponicus, its growth rate is positively correlated to the
length of photophase between the conditions of LD 12:12h and 18:6h (Wassmer and
Page, 1993). Moreover, the adult diapause of the bean bug, Riptortus clavatus, is
controlled by the photoperiod that larvae experienced. Diapause is triggered by the
suppression of corpus allatum activity under short-day conditions (Morita et al., 1999).
However, locomotor activities of the German cockroach remained the same under LD
16:8h and 12:12h conditions. Since the photoperiodism is important for the insects
living in the Temperate Zone to time the seasonal changes, its significance is greatly
reduced for the animals living in the low latitude whereas no or less drastically
changed seasons in the environment. Although the German cockroach is a
cosmopolitan species, it is a completed household species which lives in a relative
stable environment. It is not essential to express this type of photoperiodic response
because its continuous development in the man-made shelters throughout the years.
After-effects of consecutive entrainments were detected in various animals
al., 1997; Page, 2000; Page et al., 2001). In general, after-effects of photoperiodic
response included three aspects: (1) free running period (τ) changes; (2) various
length of activity time (α); (3) different phase-response curves.
Firstly, the most dramatic changes of the after-effects was found in the
lengthening or shortening the free running period. This phenomenon was not only
found in the insect species (Page et al., 2001), but also existed in several species of
rodent and sparrow (Binkley and Mosher, 1986; Pittendrigh and Daan, 1976). For
those photoperiods equal to 24 hours, the free-running period became shorter
immediately after a long-photophased Zeitgeber (LD 16:8h) than that of a
short-photophased Zeitgeber (LD 1:23h, or LD 8:16h). However, free running
periods (τ) of the German cockroach were remained the same (near 23.5h) following
two different Zeitgebers (LD 16:8h and LD 12:12h) (Fig. 3). This difference maybe
contributed to the small difference (4hrs) between the photophases of the two
Zeitgebers. Under non-24 photoperiod conditions, the after-effects of τ was more
remarkable (Page et al., 1997; Page et al., 2001). The after-effects of τ of the
cockroach, Leucophaea maderae, were significantly different after either Zeitgeber
LD 8:14h or 8:18h. Even though the after-effects of prior photoperiod on τ might
have lingered throughout several days (Binkley and Mosher, 1986), eventually it
Zeitgeber had exerted influence on the working property of the circadian clock, the
resistance of the τ changes in the German cockroach implied the quick smooth
adjustment of the clock in either entrainment or free-running conditions. The
consequence of this finding suggests a well-adapted cockroach in a relatively
turbulent photoperiod environment (artificial lighting in human shelters).
Secondly, since acrophase reflects the adaptive behaviors that are timing the
right moments for the survival, animals can be grouped as diurnal, nocturnal and
crepuscular. When the active phase of the photoperiod become longer, activity time
(α)should also increase accordingly. The German cockroach was active during scotophase as a typical nocturnal animal and its α did correspond to the length of
darkness (Table 1). The after-effects showed two folds of influence on α, when the
insects were put under DD condition after a Zeitgeber. During free-running
condition, the α not only reflected the acrophase of the circadian clock, but also
showed the residue effects of previous Zeitgebers. This phenomenon was found in
diurnal sparrow (Binkley and Mosher, 1986), and the German cockroach (Table 1).
Although the quantification of the after-effects has not been investigated in previous
reports the same α distribution patterns after experiencing two consecutive Zeitgebers
(Fig. 4) provide a potential to construct a model for analyzing the interactions
Thirdly, the after-effects of previous Zeitgebers can and do contribute in the
phase-response curve (PRCs), plots the magnitude and direction of phase shifts
induced by light pulses, and it is a characteristic of a species (Johnson, 1999).
Although many PRCs of various species have been reported, the adjustability of the
circadian clock shown by the PRC could not be compared between species, if the
Zeitgebers were different. The comparison between Nauphoeta cinerea and B.
germanica was invalid because the different Zeitgebers caused the inbalance between
subjective day and night (LD 12:12h for the former (Saunders, 1977) and for the later
16:8h (Wei and Lee, 2001), respectively). Researchers have found that sparrows
expressed different PRCs when they experienced different Zeitgebers (LD 16:8h and
18:6h), but the shapes of the curves were similar (Binkley and Mosher, 1986).
Based on our analysis of locomotion data, the subjective day and night could be
clearly defined by the light-dark cycle before constant condition. Every PRC,
therefore, can be re-adjusted to a standard subjective day and night of 12:12h. Then,
the comparison on adjustability of the circadian clock between species becomes a
possible task.
Up to date, none of molecular mechanism has been proposed to underlie the
after-effects. However, after-effects did show the history-dependent influences of
reveals the ecological significance of clock as a mean to cope with the rhythmic
changes of environment. The previous Zeitgebers had a profound effect on the
expression of circadian rhythm under current environmental conditions. The
possible function of this characteristic may lie on the acceleration of adjustment to the
rapid changing environment.
參考文獻:
Brady, J., 1982. Biological Timekeeping. Cambridge University Press, Cambridge.
Barrett, R. K., and T. L. Page, 1989. Effects of light on circadian pacemake
development. Journal of Comparative Physiology A 165, 41-49.
Binkley, S., and K. Mosher, 1986. Photoperiod modifies circadian resetting responses
in sparrows. American Journal of Physiology 20, 1156-1162.
Colot, H. V., J. C. Hall, and M. Rosbash. 1988. Interspecific comparasion of
period gene of Drosophila reveals large blocks of non-conserved coding DNA.
EMBO 7: 3929-3937.
Crispi, S., E. Giordano, P. P. D’Avino, and M. Furia. 1998. Cross-talking among
Drosophila nuclear receptors at the promiscuous response element of the ng-1
and ng-2 intermolt genes. J. Mol. Bio. 275: 561-574.
Darlington, T. K., K. Wager-Smith, M. F. Ceriani, D. Staknis, N. Gekakis, T. D. L.
circadian loop: CLOCK-induced transcription of its own inhibitors per and tim.
Scinece 280: 1599-1603.
Dunlap, J. C. 1999. Molecular bases for circadian clocks. Cell 96: 271-290.
Glossop, N. R. J., L. C. Lyons and P. E. Hardin. 1999. Interlocked feedback loops
within the Drosophila circadian oscillator. Science 286: 766-768.
Helfrich-Forster C., M. Stengl, and U. Homberg. 1998. Organization of the
circadian system in insects. Chronobiol. Int. 15: 567-594.
Hogenesch, J. B., Y. Gu, S. Jain and C. A. Bradfield. 1998. The
Basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active
complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 95:
5474-5479.
Honig, M. G., and R. I. Hume. 1989. DiI and DiO: versatile fluorescent dyes for
neuronal labeling and pathway tracing. TINS 12: 333-341.
Jindra, M., F. Malone, K. Hiruma, and L. M. Riddiford. 1996. Developmental
profiles and ecdysteroid regulation of the mRNAs for two ecdysone receptor
isoforms in the epidermis and wings of the tobacco hornworm, Manduca
sexta. Dev. Bio. 180: 258-272.
Johnson, C. H., 1999. Forty years of PRCs-What have we learned? Chronobiology
International 16, 711-743.
King-Jones, K., G. Korge, and M. Lehmann. 1999. The helix-loop-helix proteins
dAP-4 and daughterless bind both in vitro and in vivo to SEBP3 sites required
for transcriptional activation of the Drosophlia Gene Sgd-4. J. Mol. Bio.
291: 71-82.
Koehler W. K., and G. Fleissner. 1978. Internal desynchronization of bilaterally
708-710.
Koehler, P. G., C. A. Strong, and R. S. Patterson, 1994. Harborage width preferences
of German cockroach (Dictyoptera: Blattellidae) adults and nymphs.
Household and Structural Insects 87, 699-704.
Konopka, R. J. and S. Benzer. 1971. Clock mutants of Drosophila melanogaster.
Proc. Natl. Acad. Sci. USA 68: 2112-2116.
Koontz, M., and J. S. Edward. 1980. The projection of neuroendocrine fibers in
the brain of three Orthopteroid insect. J. Morphol. 165: 285-299.
Lee, C., K. Bae, and l. Edery. 1999. PER and TIM inhibit the DNA binding
activity of a Drosophila CLOCK-CYC/dBMALI heterodimer without
disrupting formation of the heterodimer: a basis for circadian transcription.
Molecular and Cellular Biology 19: 5316-5325.
Lee, H. J., and W. Loher, 1993. The mating strategy of the male short-tailed cricket,
Anurogryllus muticus de Geer. Ethology 95, 327-344.
Lee, H. J. and Y. L. Wu. 1994. Mating effects on the feeding and locomotion of
the German cockroach, Blattella germanica. Physiol. Entomol. 19: 39-45.
Leppla, N. C., and P. G. Koehler, and H. R. Agee, 1989. Circadian rhythms of the
German cockroach (Dictyoptera: Blattellidae): locomotion in response to
different photoperiods and wavelengths of light. Journal of Insect Physiology
35, 63-66.
Lezzi, M., T. Bergman, J. -F. Mouillet, and V. C. Henrich. 1999. The ecdysone
receptor puzzle. Arch. Insect Bio. Physio. 41: 99-106.
Lin, G. H., R. F. Liou, and H. J. Lee, 2002. The period gene of the German cockroach
Chronobiology International 19, 1023-1042.
Lin, T. M., and H. J. Lee. 1996. The expression of locomotor circadian rhythm in
female German cockroach, Blattella germanica (L.). Chronobiol. Int. 13:
81-91.
Lin T. M., and H. J. Lee. 1998. Parallel control mechanisms underlying
locomotor activity and sexual receptivity of the female German cockroach,
Blattella germanica (L.). J. Insect Physiol. 44: 1039-1051.
Lococo, D. J., and S. S. Tobe. 1984a. Neuroanatomy of the retrocerebral
complex, in particular the pars intercerebralis and pars lateralis in the
cockroach Diploptera punctata. Int. J. Insect Morphol. Embryol. 13: 65-76.
Lococo, D. J., and S. S. Tobe. 1984b. Retrograde and orthograde axon transport by
brief-exposure to nickel chloride: methodology and parameters for success in
the brain-retrocerebral complex of the cockroach Diploptera punctata. J.
Insect Physiol. 30: 635-642.
Morita, A., K. Soga, T. Hoson, S. Kamisaka, and H. Numata, 1999. Changes in
mechanical properties of the cuticle and lipid accumulation in relation to adult
diapause in the bean bug, Riptortus clavatus. Journal of Insect Physiology 45,
241-247.
Moore, D., 2001. Honey bee circadian clock: behavioral control from individual
workers to whole-colony rhythms. Journal of Insect Physiology 47, 843-857.
Page, T. L., 2000. A novel mechanism for the control of circadian clock period by
Page, T.L., R. K. Barrett, 1989. Effects of light on circadian pacemaker development.
II. Responses to light. Journal of Comparative Physiology A 165, 51-59.
Page, T.L., Mans, C., Griffeth, G., 2001. History dependence of circadian pacemaker
period in the cockroach. Journal of Insect Physiology 47, 1085-1093.
Page, T.L., Wassmer, G.T., Fletcher, J., Block, G.D., 1997. Aftereffects of entrainment
of the period of the pacemaker in the eye of the mollusk Bulla gouldiana.
Journal of Biological Rhythms 12, 218-255.
Pittendrigh, C.S., Daan, S., 1976. A functional analysis of circadian pacemakers in
nocturnal rodents. I. The stability and lability of spontaneous frequency.
Journal of Comparative Physiology A 106, 223-252.
Plautz, J. D., M. Kaneko, J. C. Hall, and S.A.Kay. 1997. Independent
photoreceptive circadian clock throughout Drosophila. Science 278:1632-1635.
Saez, L. and M. W. Young. 1988. In situ localization of the per clock protein
during development of Drosophila melanogaster. Mol. Cell. Biol. 8:
5378-5385.
Sassone-Corsi, P. 1998. Molecular clocks: mastering time by gene regulation.
Nature 392:871-874.
Sauman, I. and S. M. Repper. 1996. Circadian clock neurons in the silkmoth
Antheraea pernyi: novel mechanisms of Period protein regulation. Neuron
17: 889-900.
locomotor activity in a cockroach (Nauphoeta cinerea). Nature 270, 241-243.
Sokolove, P. G., and W. N. Bushell, 1978. The chi square periodogram: Its utility for
analysis of circadian rhythms. Journal of Theoretical Biology 72, 131-160.
Stocker, A. J., J. M. Amabis, E. Gorab, C. Elke, and M. Lezzi. 1997. Antibodies
against the D-domain of a Chironomus ecdysone receptor protein react with
DNA puff sites in Trichosia pubescens. Chromosoma 106:456-464.
Takahashi, J. S. 1995. Molecular neurobiology and genetics ofcircadian rhythms
in mammals. Annu. Rev. Neurosci. 18: 531-553.
Tsai, C. W., and H. J. Lee, 2000. Circadian locomotor rhythm masked by the female
reproduction cycle in cockroaches. Physiological Entomology 25, 63-73.
von Bartheld, C. S., D. E. Cunningham, and E. W. Rubel. 1990. Neuronal tracing
with DiI: decalcification, cryosectioning, and photoconversion for light and
electron microscopic analysis. J. Histochem. Cytochrm. 38: 725-733.
Wassmer, G. T., and T. L. Page, 1993. Photoperiodic time measurement and a graded
response in a cockroach. Journal of Biological Rhythms 8, 47-56.
Wei, Y. S., and H. J. Lee, 2001. Adjustability of the circadian clock in the
cockpoaches: A comparative study of two closely related species, Blattella
germanica and Blattella bisignata. Chronobiology International 18, 767-780.
Wen, H. W., and H. J. Lee. 2000. Unequal coupling between locomotor
pacemakers of the German cockroach, Blattella germanica(L.). J. Insect
Physiol. 46: 89-97.
of the German cockroach, Blattella germanica. Journal of Insect Physiology
46 89-97.
Wills S. A., T. L. Page, and C. S. Colwell. 1985. Circadian rhythms in the