ELSEVIER Plant Science 114 (1996) 121-128
Chuan Chi Lin, Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan. Republic of China Received 18 May 1995; revision received 12 December 1995; accepted 18 December 1995
Abstract
The effects of NaCl on changes in the proline level in roots and root growth of rice seedlings were investigated. NaCl was effective in inhibiting root growth and in increasing proline accumulation in roots. Mannitol was also effective in inhibiting root growth, but proline accumulation was observed only at the concentration iso-osmotic with 150 mM NaCI. Accumulation of proline in roots preceded inhibition of root growth caused by NaCl. The inhibition of root growth and accumulation of proline in roots by NaCl are reversible. Exogenous L-proline and gabaculine, which were found to enhance in an accumulation of proline in roots, inhibited root growth of rice seedlings. The relationship be- tween growth inhibition of roots caused by NaCl and changes in the proline level in roots is discussed.
Keywords: Gabaculine; NaCI; Oryza suriva L.; Proline; Root growth
1. Introduction
Proline accumulation in plant cells exposed to salt or water stress is a widespread phenomenon
[ 1,2]. Proline is believed to protect plant tissues against stress by acting as an N-storage com- pound, osmo-solute and hydrophobic protectant for enzymes and cellular structures [3]. However, its role in imparting resistance to salt stress con- tinues to be controversial. Some workers consider enhanced proline levels simply as a stress effect, rather than a cause of stress tolerance [4-61. There are reports that proline accumulation is correlated with growth inhibition induced by NaCl [7,8]. The
* Corresponding author.
role of proline accumulation in salinity stress therefore needs to be critically examined. In this paper, we investigate the role of proline in the con- trol of NaCl-inhibited root growth of rice seedlings.
2. Materials and methods
Rice (Oryza sutiva L., cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlor- ite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dishes (20-cm) containing distilled water at 37°C under dark conditions. After l-day incubation, uniformly germinated seeds were selected and transferred to a Petri dish (9.0-cm) containing 2
0168-9452/96/315.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved PII SO168-9452(96)04323-3
122 C.C. Lin, C.H. Kao/PIanr Science 114 (19%) 121-128
sheets of Whatman No. 1 filter paper moistened with 10 ml distilled water or test solution. All test compounds were dissolved in distilled water. Their pHs are similar to that of distilled water, which is 5.5. Each Petri dish contained 20 germinated seeds. Each treatment was replicated 4 times. The germinated seeds were allowed to grow at 27°C in darkness and an additional 3 ml distilled water or test solutions was added to each Petri dish on day 3 of the growth.
Proline was extracted and its concentration determined by the method of Bates et al. [9]. Roots were homogenized with 3% sulfosalicylic acid and the homogenate was centrifuged. The supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h and then absorbance at 520 nm was determined. Levels of proline are expressed as PmoYg dry weight.
For all measurements, each treatment was re- peated 4 times. All experiments described here were repeated 3 times. Similar results and identical trends obtained each time. The data reported here are from a single experiment.
3. Results
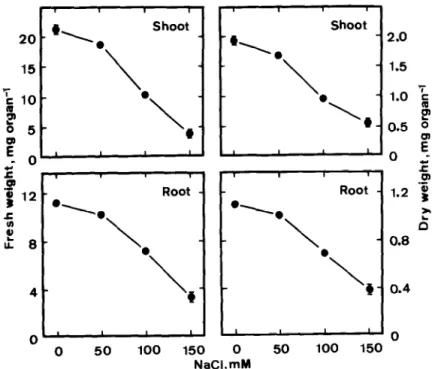
Fig. 1 shows the effect of NaCl on the growth of rice seedlings. NaCl significantly reduced growth of shoots and roots, as judged by fresh weight and dry weight, of rice seedlings. Increasing concentra- tions of NaCl from 50 to 150 mM progressively decreased root and shoot growth. Similar findings were recently reported when seedling growth was measured in terms of root and shoot length [lo]. These results are in general agreement with those reported by Flowers and Yeo [ 111.
The effects of NaCl on proline levels in shoots and roots of rice seedlings are shown in Fig. 2. In- creasing concentrations of NaCl from 50 to 150 mM progressively increased proline levels in roots. However, proline accumulation in shoots was observed only when seedlings were treated at high concentration (150 mM) of NaCI. Clearly, the reduction of root growth, but not shoot growth, is closely associated with the accumulation of proline in roots.
Mannitol inhibited root growth, but proline ac-
20 15 Shoot Root -
l
I. \ 0 \- @ NaCl,mM t Shoot Il. \ Root 2.0 1.5 c UJ 1.2 ‘f i? D 0.8 0.4 0Fig. 1. Effects of NaCl on seedling growth of rice. Seedling growth was measured after 5 days of treatment. Vertical bars represent standard errors. Only those standard errors larger than symbol size are shown.
C.C. Lin. C.H. Kao/Planl Science 114 (19%) 121-128 I I I I
25 -
Shoot 20 - 15 - I .zP
p 25 0 50 100 150 NaCl,mMFig. 2. Effects of NaCl on proline levels in shoots and roots of rice seedlings. Proline level was determined after 5 days of treatment. Vertical bars represent standard errors. Only those standard errors larger than symbol size are shown.
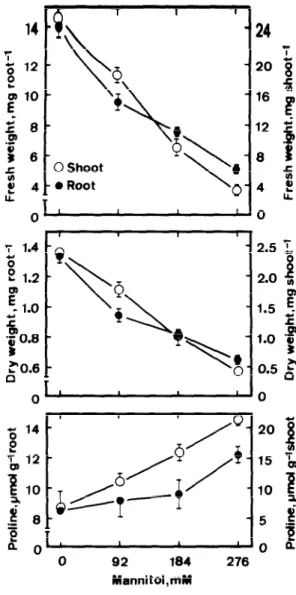
cumulation was observed only at a concentration iso-osmotic with 150 mM NaCl (Fig. 3). Mannitol treatment also inhibited shoot growth and resulted in an accumulation of proline. All these results suggest that proline accumulation in NaCl-treated roots and shoots is associated with the ionic rather than the osmotic component of NaCl stress.
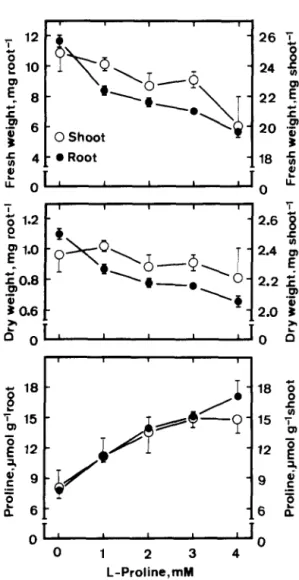
If proline accumulation is important in regul- ating growth inhibition of roots but not shoots of rice seedlings caused by NaCl, then exogenous application of proline is expected to inhibit root growth but not shoot growth of rice seedlings. As indicated in Fig. 4, exogenous L-proline treatment resulted in an inhibition of root growth and an increase in proline level in roots. However, addi- tion of L-proline did not affect shoot growth (Fig. 4). The effect of L-proline on root growth is unlikely to be due to microorganisms, since inhibi- tion of root growth was also obtained when L- proline was added together with chloramphenicol
0 Shoot - 0 Root 123
24
i 20 3 f 18 F 12 ; 5 8 Y f 4 ; 0 - 2.5 ‘; 0 92 184 276 Mannitol.mMFig. 3. Effects of mannitol on seedling growth and proline level in rice seedlings. Mannitol concentrations of 92, 184, 276 mM were iso-osmotic with 50, 100 and I50 mM, respectively. Seed- ling growth and proline level were measured after 5 days of treatment. Vertical bars represent standard errors.
(0.25 mg/l) to prevent bacterial growth. Chlor- amphenicol itself had no effect on root growth. In a recent work, we demonstrated that L-proline enhanced NaCl-induced inhibition of root growth, but reversed the inhibitory effect of NaCl on shoot growth of rice seedlings [ 121. It seems that proline may play different roles in regulating shoot and root growth of rice seedlings under NaCl con- ditions.
124 C.C. Kao/Plant Science 114 (19%) 121-128 7 ‘2- - 26 L *ci g 10 - t 8- E .P P 6- 0 Shoot f 4 -@Root -18 $, L 00 1.2
k
1.0 E go.8 ‘6 3 0.6 *I 0’ 0 Trn 15 z E ‘2 3 f 9 ._ $ 6 0 -06
I
0 1 2 3 4 L-Proline,mM fFig. 4. Effects of L-proline on seedling growth and proline level in rice seedlings. Seedling growth and proline level were measured after 5 days of treatment. Vertical bars represent standard errors. Only those standard errors larger than symbol size are shown.
Kandpal and Rao [ 131 have proposed that or- nithine aminotransferase may be a potential point of control of proline synthesis in plants subjected to stress. Gabaculine is known to inhibit omithine aminotransferase [14] and has been shown to increase proline accumulation in radish cotyledons
[ 151. To characterize further the role of proline ac- cumulation in regulating root growth reduction of rice seedlings under NaCl stress, rice seedlings were grown in the presence of various concentra- tions of gabaculine. As indicated in Fig. 5, gabaculine increased the proline level in roots and
21 - T 38 - E 5 15 - e 5 12 - tz 9- 6- 0- 0 10 20 30 40 50 Gabaculine,yM
Fig. 5. Effects of gabaculine on root growth and proline level in roots of rice seedlings. Root growth and proline level were measured after 5 days of treatment. Vertical bars represent standard errors. Only those standard errors larger than symbol size are shown.
inhibited root growth. From the shape of the curves, it is apparent that the proline level is close- ly linked with root growth reduction.
C.C. Lin. C.H. Kao/Plant Science 114 (19%) 121-128 125
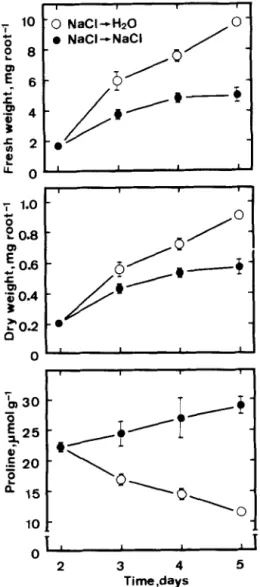
or gabaculine are transferred to distilled water, they then resume root growth (Figs. 6 and 7). Root growth was only slightly restored if seedlings re- mained in NaCl or gabaculine (Figs. 6 and 7). Figs. 6 and 7 also show that the proline level in roots of seedlings remaining in NaCl or gabaculine
- 0 NaCI+H20 l NaCl +NaCI 7 1.0 - 5 0 L 0.8 0- 2 3 4 5 Time ,days
Fig. 6. Changes in growth of and proline level in roots of NaCI- treated rice seedlings grown in the presence or absence of NaCl (I 50 mM). Rice seeds were germinated for 2 days in NaCl(150 mM) and then seedlings were transferred to distilled water (0) and NaCl (0). respectively. Vertical bars represent standard er- rors. Only those standard errors larger than symbol size are shown. I I I 0 GabaculinedH20 l Gabaculine-- Gabaculine 7 I I ; 0.8 -
e
a 0.6 - E / 0’ ,0-e-.- i 0- 2 3 4 5 Time,daysFig. 7. Changes in growth of and proline level in roots of gabaculine-treated rice seedlings grown in the presence or ab- sence of gabaculine (25 PM). Rice seeds were germinated for 2 days in gabactdine (25 PM) and then seedlings were transferred to distilled water (0) and gabaculine (0) respectively. Vertical bars represent standard errors. Only those standard errors larger than symbol size are shown.
medium is higher than that of seedlings transferred to distilled water.
To test the causal relationship between proline accumulation and root growth reduction caused by NaCI, 2-day old seedlings were transferred to distilled water and NaCl medium for 4, 8, 12 and 16 h, respectively. Changes in proline levels in roots and root growth were then monitored. As in-
126 C.C. Lin. C.H. Kao/PIant Science 114 (19%) 121-128 i 6 0 H20+H20 2 7 t 0 H20-NaCl ^ 6 E ul ._ $ 5 I- 25 ‘i 020 - Z $5 - ./I /f_ / g 10 -&~_~_o-o- k 5- 0- 0 4 6 12 16 Time,h
Fig. 8. Changes in growth of and proline level in roots of NaCI- treated rice. seedlings grown in the presence or absence of NaCl (I50 mM). Rice seeds were germinated for 2 days in distilled water and then seedlings were transferred to distilled water (0) and NaCl (a), respectively. Vertical bars represent standard er- rors. Only those standard errors larger than symbol size are shown.
dicated in Fig. 8, proline accumulation was observed at 4 h after NaCl treatment, whereas root growth was observed to be inhibited at 8 h.
4. Discusion
Rice is a salt-sensitive species and accumulates Two pathways of proline biosynthesis, gluta- sodium ion at only moderate external salinities mate and omithine pathways, have been reported [16]. Among different parameters responding to [25]. In higher plants, the proline biosynthesis NaCl-stress in rice, the most significant one is rapid accumulation of proline [ 17,181. The present investigation shows that the proline level in roots is closely associated with the NaCl concentration applied, whereas proline accumulation in shoots was observed only at high concentration of NaCl (150 mM). We also indicate that proline accumula- tion in NaCl-treated roots is associated with the ionic rather than the osmotic component of NaCl stress.
The present investigation indicates that proline accumulation participates in the regulation of root growth of rice seedlings under NaCl conditions. This conclusion was based on the observations that (a) proline accumulation in roots preceded root growth reduction caused by NaCl, (b) proline level in roots is inversely associated with root growth and (c) NaCl-induced root growth reduc- tion is produced in rice seedlings fed with L- proline or gabaculine which was found to result in a proline accumulation. When rice seedlings pretreated with NaCl or gabaculine were transfer- red to distilled water, root growth resumed and proline level in roots declined. These results sug- gest that growth inhibition of roots induced by NaCl is not irreversible and further support the close link between proline accumulation and root growth reduction induced by NaCl. Proline ac- cumulation induced by NaCl or Na2S04 has also been demonstrated to be correlated with growth inhibition of callus [7,8].
Le Dily et al. [19] reported that the polyamine level of a habituated callus of Beta vulgar-is was strongly diminished after treatment with gabacul- ine. It has been shown that exogenous application of polyamine can overcome the harmful effects of NaCl [20-221 and endogenous levels of poly- amines decreased under NaCl stress [23]. How- ever, our recent work provided evidence to show that endogenous polyamines do not play a signifi- cant role in the control of NaCl-inhibited growth of rice seedlings [24]. Thus, the inhibition effect of gabaculine on root growth of rice seedlings is unlikely to be due to the decrease of polyamine levels.
C. C. Lin, C. H. Kao /Plant Science 114 (19%) 121-128 127
route is generally
thought
to involve
the glutamate
pathway which implies pyrroline S-carboxylate
synthase and pyrroline 5-carboxylate reductase 1251. In a few studies with germinated seedlings, a pathway using omithine as the proline precursor has been put forward, in which ornithine amino- transferase is the key enzyme [25]. We do not know whether these two pathways contribute to the accumulation of proline in NaCl-treated roots. Further studies should provide valuable informa- tion on the mechanism of proline accumulation in rice roots in response to NaCl. In animal systems, omithine aminotransferase is a reversible enzyme. It has been proposed that omithine aminotrans- ferase, which transforms glutamate 5-semialde- hyde and glutamate into omithine and keto- glutarate, could link proline catabolism to the syn- thesis of omithine [26]. Gabaculine has been shown to be a potent inhibitor of omithine amino- transferase [27]. It appears that proline accumula- tion in gabaculine-treated rice roots is most likely due to the inhibition of proline catabolism.
Possible functional roles of proline under stress conditions, as noted by Aspinall and Paleg [I], may include (a) cytoplasmic osmoticum, (b) the hydration of polymers, and (c) serving as a nitro- gen source compound during periods of inhibited growth. Since growth is generally considered to be an energy-requiring process, elevated proline lev- els caused by NaCl are most likely acting as a way to save energy by inhibiting root growth and as a readily utilizable source of energy and amino groups once NaCl is relieved.
Acknowledgements
This work was supported financially by the Na- tional Science Council of the Republic of China (NSC 84-2321-B-002-097) to C.H.K.
References
HI
121
D. Aspinall and L.G. Paleg, Proline accumulation: Physiological aspects, in: L.G. Paleg and D. Aspinall (Eds.), The Physiology and Biochemistry of Drought Re- sistance in Plants, Academic Press, Sydney, 1981, pp. 205-241.
S.F. Chandler and T.A. Thorpe, Variation from plant tis-
[31 141 PI VI 171 VI [91 UOI VII WI [I31 iI41 WI 1161 1171
sue cultures: biotechnological application to improving salinity tolerance. Biotech. Adv., 4 (1986) 117-135.
H. Greenway and R. Munns, Mechanism of salt toler- ance in nonhalophytes. Annu. Rev. Plant Physiol., 31 (1980) 149-190.
A.E. Moftah and B.E. Michel, The effect of sodium chlo- ride on solute potential and proline accumulation in soy- bean leaves. Plant Physiol., 83 (1987) 238-240. A.D. Hanson, C.E. Nelsen and E.H. Everson, Evaluation of free proline accumulation as an index of drought resis- tance using two contrasting barley cultivars. Crop Sci., 17 (1977) 720-726.
A.D. Hanson, C.E. Nelsen, A.R. Pederson and E.H. Everson, Capacity for proline accumulation during water stress in barley and its implication for breeding for drought resistance. Crop Sci., 19 (1979) 489-493. SF. Chandler and T.A. Thorpr, Characterization of growth, water relations, and proline accumulation in so- dium sulfate tolerant callus of Erassica napus L. cv. Westar (Canola). Plant Physiol., 84 (1987) 106-I 11. F. Perez-Alfocea, A. Santa-Cruz, G. Guerrier and MC. Bolarin, NaCl stress-induced organic solute changes on levels and calli of Lycopersicon esculentum, L. Pennellii and their interspecific hybrid. J. Plant Physiol., 143 (1994) 106-I 11.
L.S. Bates, R.P. Waldren and I.D. Teare, Rapid deter- mination of free proline for water-stress studies. Plant Soil, 39 (1973) 205-207.
C.C. Lin and C.H. Kao, NaCl stress in rice seedlings: effect of exogenous polyamine on seedling growth. Chinese Agron. J., 4 (1994) 167-171.
T.J. Flowers and A.R. Yeo, Variability in the resistance of sodium chloride salinity within rice (Oryza saliva L.) varieties. New Phytol., 88 (1981) 363-373.
C.C. Lin and C.H. Kao, NaCl stress in rice seedlings: ef- fects of t-proline, glycinebetaine, L- and o-asparagine on
seedling growth. Biol. Plant, 37 (1995) 305-308. R.P. Kandpal and N.A. Rao, Water stress induced alter- nations in the properties of omithine aminotransferase from ragi (Eleusine coracana) leaves. B&hem. Int., 5 (1982) 297-302.
F. Hervieu, F. Le Dily, J. Le Saos, J.-P. Billard and C. Huault, Inhibition of plant omithine aminotransferase by gabaculine and 4-amino-5-hexynoic acid. Phytochem- istry, 34 (1993) 1231-1234.
F. Le Dily, J.-P. Billard, F. Le Saos and C. Huault, Ef- fects of NaCl and gabaculine on chlorophyll and proline levels during growth of radish cotyledons. Plant Physiol. B&hem., 31 (1993) 303-310.
R.A. Yeo and J.T. Flowers, Accumulation and localiza- tion of Na+ within the shoots of rice (Oryza saliva L.) varieties differing in salinity resistance. Physiol. Plant., 56 (1982) 343-348.
R. Krishnamurthy, M. Anbazhagan and A.K. Bhagwat, Accumulation of free amino acids and distribution of Na+, Cl- and K+ in rice varieties exposed to NaCl stress. Indian J. Plant Physiol., 30 (1987) 183-188.
128 C.C. Lin. C.H. Kao/Plant Science 114 (19%) 121-128
[I81 D. Roy, A. Bhunia, N. Basu and S.K. Banerjee, Effect of NaCl-salinity on metabolism of proline in salt-sensitive and salt-resistant cultivars of rice. Biol. Plant., 34 (1992)
159-162.
[I91 F. Le Dily, C. Huault, Th. Gaspar and J.-P. Billard, Gabaculine as a tool to investigate the polyamine biosyn- thesis pathway in habituated callus of Beta vulgaris (L.). Plant Growth Regul., 13 (1993) 221-223.
[20] R. Krishnamurthy, Amelioration of salinity effect in salt tolerant rice (Oryza saliva L.) by foliar application of putrescine. Plant Cell Physiol., 32 (1991) 699-703. [21] L. Prakash and G. Prathapasenan, Effect of NaCl salin-
ity and putrescine on shoot growth, tissue ion concentra- tion and yield of rice (Oryza sariva L. GR3). J. Agric. Crop Sci., 160 (1988) 325-334.
[22] L. Prakash and G. Prathapasenan, Putrescine reduces NaCl-induced inhibition of germination and early seed- ling growth of rice (Oryza saliva L.). Aust. J. Plant Physiol., 15 (1988) 761-767.
[23] L. Prakash, M. Dutt and G. Prathapasenan, NaCl alters contents of nucleic acids, protein, polyamines and the ac- tivity of agmatine deiminase during germination and seedling growth of rice (Oryza sativa L.). Aust. J. Plant Physiol., 15 (1988) 769-776.
[24] C.C. Lin and C.H. Kao, Levels of endogenous polyamines and NaCl-inhibited growth of rice seedlings. Plant Growth Regul., 17 (1995) 15-20.
[25] F. Hervieu, F. Le Dily, C. Huault and J.-P. Billard, Con- tribution of omithine aminotransferase to proline ac- cumulation in NaCl-treated radish cotyledons. Plant Cell Environ., 18 (1995) 205-210.
(26) M.E. Jones, Conversion of glutamate to ornithine and proline: pyrroline-5carboxylate, a possible modulator of arginine requirements. J. Nutr., 115 (1985) 509-515. (27) J.M. Jung and N. Seiler, Enzyme-activated irreversible
inhibitors of L-omithine: 2-oxoacid aminotransferase. J. Biol. Chem., 253 (1978) 7431-7439.