Polymorphisms in XRCC1 and
Glutathione S-Transferase
Genes and Hepatitis
B–Related Hepatocellular
Carcinoma
Ming-Whei Yu, Shi-Yi Yang, I-Jen
Pan, Chih-Lin Lin, Chun-Jen Liu,
Yun-Fan Liaw, Shi-Ming Lin,
Pei-Jer Chen, Shou-Dong Lee,
Chien-Jen Chen
Chronic infection with hepatitis B vi-rus (HBV) causes DNA damage. An arginine (Arg)-to-glutamine (Gln) polymorphism at codon 399 in the XRCC1 gene is putatively associated with DNA damage. In a case–control study of 577 HBV surface antigen car-riers with hepatocellular carcinoma (HCC) and 389 HBV carrier control subjects, we investigated the associa-tion between this polymorphism and the risk of HCC and assessed whether this association varied with glutathi-one S-transferase (GST) status; GSTs are involved in carcinogen metabo-lism. All statistical tests were two-sided. The XRCC1 Gln allele was associated with a dose-dependent increased risk of early-onset HCC (<50 years) but not with the risk of late-onset HCC (Ptrend = .01). The GSTT1-null geno-type alone did not affect risk, but the GSTM1-null genotype was associated with a decreased risk for early-onset HCC. Various combinations of GSTM1 and GSTT1 genotypes differentially modified the association of XRCC1 with HCC (Pinteraction = .005); e.g., for individuals with the GSTT1-null/ GSTM1-present genotype, the risk of HCC was greater for those with the Gln/Gln genotype (odds ratio = 8.07, 95% confidence interval = 1.67 to 38.93) than for those with the Arg/Arg genotype. Thus, GST status appears to affect the risk of HCC associ-ated with this XRCC1 polymorphism. [J Natl Cancer Inst 2003;95:1485–8]
Chronic infection with hepatitis B vi-rus (HBV) is the most important cause of hepatocellular carcinoma (HCC) in Taiwan, where the predominant mode of HBV transmission is perinatal (1,2). Al-though the relative risk is about 20 for HBV surface antigen (HBsAg) carriers
compared with noncarriers, only a frac-tion of HBsAg carriers eventually de-velop HCC. The age at onset of HCC also varies over a wide range among HBsAg carriers (1,3).
Persistent HBV infection can cause genomic damage directly, through a mechanism of chromosomal integration, or indirectly, through increased oxida-tive stress and free-radical generation during the course of chronic hepatitis (1,4–7). The XRCC1 protein is required for genomic stability and participates in the repair of endogenous oxidative DNA damage and DNA single-strand breaks (8–10). The XRCC1 gene has an argi-nine (Arg)-to-glutamine (Gln) polymor-phism at codon 399 (XRCC1-Arg399Gln polymorphism) that has been associated with DNA damage phenotypes and can-cer risk (11–15). We conducted a mul-ticenter case–control study to examine whether this polymorphism influences risk for HCC among HBsAg carriers. We specifically investigated whether this association varied by age at onset of HCC and the status of glutathione S-transferase genes GSTM1 and GSTT1, which are involved in the metabolism of potential carcinogens, including en-dogenously generated cancer-causing reactive oxygen species continuously produced through HBV-induced inflam-matory disease (16,17).
The study included 577 HBsAg car-riers with incident HCC selected among 1572 patients who participated in our ongoing genetic epidemiology study of
Affiliations of authors: M.-W. Yu, S.-Y. Yang,
I-J. Pan, C.-J. Chen, Graduate Institute of Epide-miology, College of Public Health, National Tai-wan University, Taipei, TaiTai-wan; C.-L. Lin, Divi-sion of Gastroenterology, Department of Internal Medicine, Taipei Municipal Jen-Ai Hospital, Tai-pei; C.-J. Liu, P.-J. Chen, Hepatitis Research Cen-ter, National Taiwan University Hospital, Taipei; S.-D. Lee, Department of Medicine, Veterans General Hospital and School of Medicine, Na-tional Yang-Ming University, Taipei; Y.-F. Liaw, S.-M. Lin, Liver Research Unit, Chang-Gung Memorial Hospital, Chang-Gung University, Taoyuan, Taiwan.
Correspondence to: Ming-Whei Yu, PhD,
Graduate Institute of Epidemiology, College of Public Health, National Taiwan University, No. 1 Jen-Ai Rd., Sec. 1, Rm. 1550, Taipei 100, Taiwan (e-mail: [email protected]).
See “Notes” following “References.”
DOI: 10.1093/jnci/djg051
Journal of the National Cancer Institute, Vol. 95,
No. 19, © Oxford University Press 2003, all rights reserved.
HCC between September 1, 1997, and December 31, 2001 (3). The 1572 pa-tients were recruited from three major hospitals (Chang-Gung Memorial Hos-pital, Taipei Veterans General HosHos-pital, and National Taiwan University Hospi-tal) in northern Taiwan, representing 90% of the patients originally contacted. The diagnosis of HCC was confirmed by liver biopsy or the combination of increased ␣-fetoprotein (艌400 ng/mL) plus typical features on angiography, so-nography, or computed tomography. To include sufficient numbers of early- and late-onset patients, we selected approxi-mately equal numbers of case patients in the following five age groups: younger than 30, 30–39, 40–49, 50–59, and 60– 69 years old. Because early-onset HCC patients (<30 years old at diagnosis) are rare, only 43 patients younger than 30 years were included. Finally, the propor-tions of case patients in the five age groups were 7.5%, 21.0%, 21.3%, 28.2%, and 22.0%, respectively.
Control subjects selected from a pool of HBsAg carriers were frequency-matched to the case patients by sex and year of birth (±10 years). This HBsAg carrier pool consisted of the following two groups: 1) 5508 individuals born in or before 1958 who were enrolled dur-ing regular physical examinations at the Chang-Gung Memorial Hospital or the Government Employee Central Clinics (Taipei, Taiwan) between August 1, 1988, and June 30, 1992 (3,18) and 2) 418 individuals born between January 1, 1928, and December 31, 1987, who were biological or non-biological rela-tives selected from different families of patients with HCC, resided in northern Taiwan, and participated in our ongoing genetic epidemiology study for clinical evaluation. All the HBsAg carriers are being followed up through various chan-nels, including a clinical evaluation ev-ery 6–12 months and a data linkage with Taiwan’s national cancer registry and death certification systems. More case patients (143 males and 19 females) than cancer-free HBV carriers (68 males and 59 females) in the pool were born after 1958. Consequently, to achieve frequency matching, we calculated the expected number of control subjects needed in each sex-year of birth stratum. There were 389 control subjects selected by stratified random sampling. The study was approved by the research eth-ics committee at the College of Public
Health, National Taiwan University. All study participants provided written in-formed consent.
GSTM1 and GSTT1 genotypes were determined by a multiplex polymer-ase chain reaction that used primers as described previously (19,20). The XRCC1-Arg399Gln polymorphism was determined by a polymerase chain reac-tion–restriction fragment length poly-morphism method that used primers X1–399NF (5⬘-CAACACCCCCAAGT ACAGCC-3⬘) and 28265R (5⬘-GGCTG GGACCACCTGTGTT-3⬘). Odds ratios and 95% confidence intervals from un-conditional logistic regression models were used to evaluate relative risks. All statistical tests were two-sided.
The mean age at enrollment in this case–control study (i.e., in 2002) was 52.3 ± 12.7 years (± standard deviation) for case patients and 53.0 ± 12.5 years for control subjects. Case patients and control subjects had identical sex distri-butions (14% were women). Cigarette smoking and alcohol consumption were more frequent among case patients (57.4% and 35.9%, respectively) than among control subjects (34.2% and 17.0%, respectively). Twenty-eight per-cent of case patients and 19.3% of con-trol subjects had a first-degree relative with a history of HCC.
Among control subjects, the fre-quency of the XRCC1 genotype was consistent with the Hardy–Weinberg equilibrium (chi-square goodness of fit, P⳱ .497). The risk of HCC was statis-tically nonsignificantly greater for the Gln/Gln XRCC1 genotype than for the Arg/Arg genotype (Table 1). The mean age at diagnosis of HCC was lower for case patients with the Gln/Gln XRCC1 genotype than for case patients with the Gln/Arg or Arg/Arg XRCC1 genotype (43.8 years versus 48.9 years; P⳱ .004 by Student’s t test). After stratification by age at diagnosis of HCC, the Gln XRCC1 allele was associated with a dose-dependent increased risk of early-onset HCC (<50 years) (Ptrend ⳱ .01)
but not with the risk of late-onset HCC, although this difference was not statisti-cally significant (P ⳱ .098). No evi-dence indicates that this association var-ies by family history of HCC or by the status of cigarette smoking and alcohol consumption (data not shown).
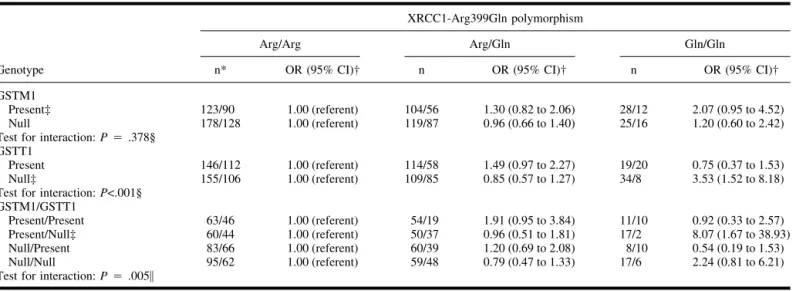
The GSTT1 genotype was not asso-ciated with the risk of HCC, overall or stratified by age at diagnosis. After
ad-justing for matching factors (sex and year of birth), a family history of HCC, tobacco and alcohol use, and other poly-morphisms, the GSTM1-null genotype was associated with a decreased risk for early-onset HCC (OR⳱ 0.64, 95% CI ⳱ 0.43 to 0.97) but not for late-onset HCC (data not shown). Table 2 shows that the association between XRCC1 and HCC is stronger for individuals with the GSTT1-null genotype than for those with the GSTT1-present genotype (Pinteraction<.001). Conversely, this
asso-ciation appeared to be stronger for indi-viduals with the GSTM1-present geno-type than for those with the GSTM1-null genotype, although the difference be-tween the two groups was not statisti-cally significant. We also observed that the association of XRCC1 with HCC was modified by GSTT1 and GSTM1 status (Pinteraction ⳱ .005). For those
with the GSTT1-null/GSTM1-present genotype, a greater risk of HCC was as-sociated with the Gln/Gln XRCC1 ge-notype than with the Arg/Arg gege-notype (adjusted OR⳱ 8.07, 95% CI ⳱ 1.67 to 38.93); the 95% confidence interval is wide because there were only two con-trol subjects with the Gln/Gln genotype in this stratum.
Extensive oxidative DNA damage has been detected in hepatocytes of HBV transgenic mice with chronic ac-tive hepatitis (5). The XRCC1–399Gln allele is associated with lower efficiency of DNA repair (11). GSTT1 protects cells from the natural by-products of lipid peroxidation and oxidative stress, and deletion of the GSTT1 gene is as-sociated with enhanced endogenous mu-tagenic processes and is implicated in susceptibility to other inflammation-related cancers, such as pancreatic can-cer, for which pancreatitis is a risk factor (17,21,22). We found that the XRCC1-399Gln allele appeared to modify the age-associated risk of HCC among HBsAg carriers. Moreover, a greater-than-additive risk of HCC was obtained when the combination of the Gln/Gln XRCC1 genotype and GSTT1-null ge-notype was analyzed. These observa-tions suggest that the lower DNA repair capacity and the increased level of reac-tive oxygen species resulting from the decreased activity of detoxification en-zyme may increase the risk of HBV-related HCC.
The XRCC1–399Gln allele has also been associated with elevated aflatoxin
B1-DNA adduct levels (11), and null
genotypes for GSTM1 and GSTT1 may increase the risk for aflatoxin B1-related
HCC (23,24). However, the highest risk of HCC associated with the Gln/Gln XRCC1 genotype for HBsAg carriers
was found in the GSTT1-null/present subgroup. Moreover, the GSTM1-null genotype was associated with
Table 1. Frequency distribution of the XRCC1-Arg399Gln polymorphism among hepatitis B surface antigen (HBsAg)–positive case
patients with hepatocellular carcinoma (HCC) and HBsAg-positive control subjects
Characteristic
XRCC1-Arg399Gln polymorphism
Arg/Arg Arg/Gln Gln/Gln Ptrend*
All case patients versus all control subjects
Case patients, No. (%) 301 (52.2) 223 (38.6) 53 (9.2)
Control subjects, No. (%) 218 (56.0) 143 (36.8) 28 (7.2)
Univariate OR (95% CI)† 1.00 (referent) 1.13 (0.86 to 1.48) 1.37 (0.84 to 2.24) .166
Multivariable OR (95% CI)ठ1.00 (referent) 1.10 (0.82 to 1.46) 1.54 (0.92 to 2.58) .129
Early-onset case patients versus control subjects born after 1948㛳
Case patients, No. (%) 142 (49.5) 112 (39.0) 33 (11.5)
Control subjects, No. (%) 110 (59.8) 60 (32.6) 14 (7.6)
Univariate OR (95% CI)† 1.00 (referent) 1.45 (0.97 to 2.16) 1.83 (0.93 to 3.58) .025
Multivariable OR (95% CI)ঠ1.00 (referent) 1.58 (1.03 to 2.42) 2.12 (1.03 to 4.35) .01
Late-onset case patients versus control subjects born in or before 1948
Case patients, No. (%) 159 (54.8) 111 (38.3) 20 (6.9)
Control subjects, No. (%) 108 (52.7) 83 (40.5) 14 (6.8)
Univariate OR (95% CI)† 1.00 (referent) 0.91 (0.62 to 1.32) 0.97 (0.47 to 2.00) .714
Multivariable OR (95% CI)ठ1.00 (referent) 0.66 (0.42 to 1.03) 1.07 (0.46 to 2.50) .294
*From Wald test for trend performed in the logistic regression model assigning scores of 1, 2, and 3, respectively, to the Arg/Arg, Arg/Gln, and Gln/Gln genotypes.
†OR⳱ odds ratio; CI ⳱ confidence interval.
‡Adjusted for year of birth (continuous variable), sex, cigarette smoking (yes or no), alcohol consumption (yes or no), first-degree family member with history of HCC, and GSTM1 and GSTT1 genotypes.
§One case patient was excluded from analysis because of missing data on alcohol consumption.
㛳The cut point for age at diagnosis between early- (<50 years) and late-onset HCC (艌50 years), 50 years old, was as defined in our previous study of familial aggregation (3). Because control subjects were frequency-matched to the case patients by year of birth and the earliest birth year for the early-onset case patients in this study is 1945, approaching the median year of birth in the control group, to compare separately with the early- and late-onset case patients, control subjects were dichotomized (born in or before 1948 or later) according to their median year of birth.
¶Likelihood ratio test for interaction of the stratified variable (onset age for case patients and year of birth for control subjects) and XRCC1 genotype was calculated as a test for the homogeneity of ORs across strata (Pinteraction⳱ .098).
Table 2. Odds ratios (ORs) and 95% confidence intervals (CIs) for the association between the Arg399Gln polymorphism in the XRCC1 gene and
hepatocellular carcinoma (HCC) among hepatitis B surface antigen (HBsAg) carriers, stratified by GSTM1 and GSTT1 genotypes
Genotype
XRCC1-Arg399Gln polymorphism
Arg/Arg Arg/Gln Gln/Gln
n* OR (95% CI)† n OR (95% CI)† n OR (95% CI)†
GSTM1
Present‡ 123/90 1.00 (referent) 104/56 1.30 (0.82 to 2.06) 28/12 2.07 (0.95 to 4.52)
Null 178/128 1.00 (referent) 119/87 0.96 (0.66 to 1.40) 25/16 1.20 (0.60 to 2.42)
Test for interaction: P⳱ .378§ GSTT1
Present 146/112 1.00 (referent) 114/58 1.49 (0.97 to 2.27) 19/20 0.75 (0.37 to 1.53)
Null‡ 155/106 1.00 (referent) 109/85 0.85 (0.57 to 1.27) 34/8 3.53 (1.52 to 8.18)
Test for interaction: P<.001§ GSTM1/GSTT1
Present/Present 63/46 1.00 (referent) 54/19 1.91 (0.95 to 3.84) 11/10 0.92 (0.33 to 2.57)
Present/Null‡ 60/44 1.00 (referent) 50/37 0.96 (0.51 to 1.81) 17/2 8.07 (1.67 to 38.93)
Null/Present 83/66 1.00 (referent) 60/39 1.20 (0.69 to 2.08) 8/10 0.54 (0.19 to 1.53)
Null/Null 95/62 1.00 (referent) 59/48 0.79 (0.47 to 1.33) 17/6 2.24 (0.81 to 6.21)
Test for interaction: P⳱ .005㛳
*n⳱ No. of case patients/No. of control subjects.
†Adjusted for year of birth (continuous variable), sex, cigarette smoking (yes or no), alcohol consumption (yes or no), first-degree family history of HCC, and the GSTM1 or GSTT1 genotype (when appropriate).
‡One case patient was excluded from analysis because of missing data on alcohol consumption.
§From the likelihood ratio test comparing the fit of the logistic model that included the main effects of various polymorphisms and all potential confounders (year of birth, sex, cigarette smoking, alcohol consumption, and first-degree family history of HCC) with a fully parameterized model containing all possible interaction terms of XRCC1 and GSTM1 (or GSTT1) genotypes.
㛳From the likelihood ratio test comparing the fit of the logistic model that included the main effects of the XRCC1 genotype, the combinations of GSTM1 and GSTT1 genotypes, and all other factors listed above with a fully parameterized model containing all possible interaction terms of XRCC1 genotype and the combinations of GSTM1 and GSTT1 genotypes.
decreased risk for early-onset HCC. Al-though the mechanisms underlying the decreased risk associated with the GSTM1-null genotype are currently un-known, another Taiwanese study (24) of HCC has identified a statistically sig-nificantly inverse association with the GSTM1-null genotype despite the prob-able synergistic interaction between af-latoxin exposure and the null type. Con-jugation with glutathione catalyzed by GSTM1 promotes elimination not only of carcinogens but also of anticarcino-genic compounds, notably isothiocya-nates that occur naturally in cruciferous vegetables (16,17,25). In fact, many studies (22,26) have evaluated the rela-tionships between GSTM1 and GSTT1 gene deletions and other cancers, but the results are equivocal.
This is, to our knowledge, the first report that the combination of a DNA repair gene polymorphism and GST ge-notype is associated with the develop-ment of HCC. Given the small number of participants in certain comparisons, our findings require confirmation in larger studies. However, in light of our hypothesis, we believe that future stud-ies on other genotypic variants involved in oxidative stress response and DNA repair are warranted.
REFERENCES
(1) Yu MW, Chen CJ. Hepatitis B and C viruses
in the development of hepatocellular carci-noma. Crit Rev Oncol Hematol 1994;17: 71–91.
(2) Stevens CE, Beasley RP, Tsui J, Lee WC.
Vertical transmission of hepatitis B antigen in Taiwan. N Engl J Med 1975;292:771–4.
(3) Yu MW, Chang HC, Liaw YF, Lin SM, Lee
SD, Liu CJ, et al. Familial risk of hepatocel-lular carcinoma among chronic hepatitis B carriers and their relatives. J Natl Cancer Inst 2000;92:1159–64.
(4) Chen PJ, Chen DS. Hepatitis B virus
infec-tion and hepatocellular carcinoma: molecular genetics and clinical perspectives. Semin Liver Dis 1999;19:253–62.
(5) Hagen TM, Huang S, Curnutte J, Fowler P,
Martinez V, Wehr CM, et al. Extensive oxi-dative DNA damage in hepatocytes of trans-genic mice with chronic active hepatitis destined to develop hepatocellular carci-noma. Proc Natl Acad Sci U S A 1994;91: 12808–12.
(6) Takada S, Gotoh Y, Hayashi S, Yoshida M,
Koike K. Structural rearrangement of inte-grated hepatitis B virus DNA as well as cel-lular flanking DNA is present in chronically infected hepatic tissues. J Virol 1990;64: 822–8.
(7) Yaginuma K, Kobayashi M, Yoshida E,
Koike K. Hepatitis B virus integration in he-patocellular carcinoma DNA: duplication of cellular flanking sequences at the integration site. Proc Natl Acad Sci U S A 1985;82: 4458–62.
(8) Whitehouse CJ, Taylor RM, Thistlewaite A,
Zhang H, Karimi-Busheri F, Lasko DD, et al. XRCC1 stimulates human polynucleotide ki-nase activity at damaged DNA termini and accelerates DNA single-strand break repair. Cell 2001;104:107–17.
(9) Vidal AE, Boiteux S, Hickson LD, Radicella
JP. XRCC1 coordinates the initial and late stages of DNA abasic site repair through pro-tein-protein interactions. EMBO J 2001;20: 6530–9.
(10) Lindahl T, Wood RD. Quality control by
DNA repair. Science 1999;286:1897–1905.
(11) Lunn RM, Langlois RG, Hsieh LL,
Thomp-son CL, Bell DA. XRCC1 polymorphisms: effects on aflatoxin B1-DNA adducts and glycophorin A variant frequency. Cancer Res 1999;59:2557–61.
(12) Kim SU, Park SK, Yoo KY, Yoon KS, Choi
JY, Seo JS, et al. XRCC1 genetic polymor-phism and breast cancer risk. Pharmacoge-netics 2002;12:335–8.
(13) Park JY, Lee SY, Jeon HS, Bae NC, Chae
SC, Joo S, et al. Polymorphism of the DNA repair gene XRCC1 and risk of primary lung cancer. Cancer Epidemiol Biomarkers Prev 2002;11:23–7.
(14) Shen H, Xu Y, Qian Y, Yu R, Qin Y, Zhou
L, et al. Polymorphisms of the DNA repair gene XRCC1 and risk of gastric cancer in a Chinese population. Int J Cancer 2000;88: 601–6.
(15) Nelson HH, Kelsey KT, Mott LA, Karagas
MR. The XRCC1 Arg399Gln polymorphism, sunburn, and non-melanoma skin cancer: evi-dence of gene-environment interaction. Can-cer Res 2002;62:152–5.
(16) Hayes JD, Pulford DJ. The glutathione
S-transferase supergene family: regulation of GST and the contribution of the isoenzymes to cancer chemoprotection and drug resis-tance. Crit Rev Biochem Mol Biol 1995;30: 445–600.
(17) Ketterer B. Glutathione S-transferases and
prevention of cellular free radical damage. Free Radic Res 1998;28:647–58.
(18) Yu MW, Chang HC, Chen PJ, Liu CJ, Liaw
YF, Lin SM, et al. Increased risk for hepatitis B-related liver cirrhosis in relatives of pa-tients with hepatocellular carcinoma in
northern Taiwan. Int J Epidemiol 2002;31: 1008–15.
(19) Pemble S, Schroeder KR, Spencer SR,
Meyer DJ, Hallier E, Bolt HM, et al. Human glutathione S-transferase theta (GSTT1): cDNA cloning and the characterization of a genetic polymorphism. Biochem J 1994;300: 271–6.
(20) Bell DA, Taylor JA, Paulson DF, Robertson
CN, Mohler JL, Lucier GW. Genetic risk and carcinogen exposure: a common inherited defect of the carcinogen-metabolism gene glutathione S-transferase M1 (GSTM1) that increases susceptibility to bladder cancer. J Natl Cancer Inst 1993;85:1159–64.
(21) Wiencke JK, Pemble S, Ketterer B, Kelsey
KT. Gene deletion of glutathione S-trans-ferase theta: correlation with induced genetic damage and potential role in endogenous mu-tagenesis. Cancer Epidemiol Biomarkers Prev 1995;4:253–9.
(22) Duell EJ, Holly EA, Bracci PM, Liu M,
Wiencke JK, Kelsey KT. A population-based, case-control study of polymorphisms in carcinogen-metabolizing genes, smoking, and pancreatic adenocarcinoma risk. J Natl Cancer Inst 2002;94:297–306.
(23) Chen CJ, Yu MW, Liaw YF, Wang LW,
Chi-amprasert S, Matin F, et al. Chronic hepatitis B carriers with null genotypes of glutathione S-transferase M1 and T1 polymorphisms who are exposed to aflatoxin are at increased risk of hepatocellular carcinoma. Am J Hum Genet 1996;59:128–34.
(24) Sun CA, Wang LY, Chen CJ, Lu SN, You
SL, Wang LW, et al. Genetic polymorphisms of glutathione S-transferases M1 and T1 as-sociated with susceptibility to aflatoxin-related hepatocarcinogenesis among chronic hepatitis B carriers: a nested case-control study in Taiwan. Carcinogenesis 2001;22: 1289–94.
(25) Kolm RH, Danielson UH, Zhang Y, Talalay
P, Mannervik B. Isothiocyanates as sub-strates for human glutathione transferases: structure-activity studies. Biochem J 1995; 311:453–9.
(26) Rebbeck TR. Molecular epidemiology of the
human glutathione S-transferase genotypes GSTM1 and GSTT1 in cancer susceptibility. Cancer Epidemiol Biomarkers Prev 1997;6: 733–43.
NOTES
Supported by grants NSC 90-2314-B-002-100 (to M.-W. Yu) from the National Science Council and DOH91-TD-1003 (National Research Pro-gram for Genomic Medicine) (to M.-W. Yu) from the Department of Health, Executive Yuan, Taiwan.
Manuscript received February 21, 2003; revised July 8, 2003; accepted July 15, 2003.