Abscisic acid induced changes in cell wall peroxidase activity and
hydrogen peroxide level in roots of rice seedlings
Chuan Chi Lin, Ching Huei Kao *
Department of Agronomy, National Taiwan Uni6ersity, Taipei, Taiwan, PR China
Received 5 June 2000; received in revised form 19 September 2000; accepted 19 September 2000
Abstract
The changes in the activity of peroxidase (POD) extracted from the cell wall and the level of H2O2of rice seedling roots treated
with abscisic acid (ABA) and their correlation with root growth were investigated. Increasing concentrations of ABA from 3 to 18mM progressively reduce root growth and increase POD activities (using guaiacol or ferulic acid as a substrate) extracted from the cell wall of rice roots. The reduction of root growth by ABA is also correlated with an increase in H2O2level. Both diamine
oxidase (DAO) and NADH peroxidase (NADH-POD) are known to be responsible for the generation of H2O2. ABA treatment
increased NADH-POD and DAO activities in roots of rice seedlings, suggesting that NADH-POD and DAO contribute to the generation of H2O2in the cell wall of ABA-treated roots. An increase in the level of H2O2and the activity of POD extracted from
the cell wall of rice roots preceded root growth reduction caused by ABA. An increase in DAO and NADH-POD activities coincided with an increase in H2O2in roots caused by ABA. Since DAO catalyzes the oxidation of putrescine, the results that
ABA increases the activity of DAO in roots is consistent with those that ABA decreases the level of putrescine. In conclusion, cell wall stiffening catalyzed by POD is possibly involved in the regulation of root growth reduction caused by ABA. © 2001 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Abscisic acid; Diamine oxidase; H2O2; Oryza sati6a; Peroxidase; Root growth
www.elsevier.com/locate/plantsci
1. Introduction
It has been suggested that growth reduction is likely to result from cell wall stiffening processes related to formation of cross-links among cell wall polymers [1]. The formation of cross-links between cell wall components is mediated by cell wall-asso-ciated peroxidase (POD) enzymes [1]. There are many reports showing that POD activity extracted from cell walls is inversely related to cell growth [2 – 8]. With the aid of specific antibodies for POD it was shown that hypocotyl growth could be enhanced in the presence of the antibodies, or
alternatively, with additional POD the growth was stunted [9].
Abscisic acid (ABA) is ubiquitous in higher plants and has many physiological effects on the growth and differentiation of plants [10]. It is usually accepted that ABA is a potent growth inhibitor, although in a few cases it may promote growth [11]. It has been shown that ABA is re-quired for maize primary root elongation at low water potentials [12,13]. However, the mechanism underlying the inhibition of plant growth by ABA is not yet clear. Zocchi and De Nisi [14] reported that ABA inhibited elongation of the maize coleoptile and reduced proton extrusion. We have previously reported that an increase in ionically bound POD (removable from homogenized tissue with high ionic strength buffer) activity is associ-ated with growth inhibition of rice seedling roots caused by ABA [15]. H2O2is a necessary substrate Abbre6iations: DAO, Diamine oxidase; DW, Dry weight; FPOD,
Ferulic acid peroxidase; FW, Fresh weight; POD, Peroxidase; Put, Putrescine; Spd, Spermidine; Spm, Spermidine.
* Corresponding author. Tel.: + 23698159; fax: + 886-2-23620879.
E-mail address:[email protected] (C.H. Kao).
0168-9452/01/$ - see front matter © 2001 Elsevier Science Ireland Ltd. All rights reserved. PII: S 0 1 6 8 - 9 4 5 2 ( 0 0 ) 0 0 3 9 6 - 4
for cell wall stiffening process catalyzed by POD [5,16]. Using a sensitive tissue-print assay, Schopfer [17] has been able to demonstrate that H2O2 is localized in the cell wall of pea epicotyl.
The formation of H2O2 by isolated cell wall from
horseradish has been reported [18]. It has been shown that H2O2 inhibits auxin-mediated growth
of maize coleoptile segments [16] and H2O2causes
a rapid cross-links of cell wall polymers [16,19]. Therefore, to ensure complete stiffening of the cell wall a sufficient supply of H2O2 is required. The
present investigation was designed to study the changes in POD activity extracted from the cell wall and H2O2 level in roots of ABA-treated rice
seedlings and their correlation with root growth.
2. Materials and methods
Rice (Oryza sati6a L., cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlor-ite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dish (20 cm) containing distilled water at 37°C under dark condition. After 1 day incubation, uniformly germinated seeds were selected and transferred to Petri dishes (9.0 cm) containing two sheets of Whatman No. 1 filter paper moistened with 10 ml of distilled water or test solutions. Each Petri dish contained 20 germinated seeds. Each treatment was replicated 4 times. The germinated seeds were allowed to grow at 27°C in darkness. To avoid the loss by evaporation and uptake by the seeds, a further 3 ml of distilled water or test solutions was added to each Petri dish on day 3. Cell walls were prepared by homogenizing roots in ice-cold phosphate buffer (50 mM, pH 5.8) using a pestle and mortar. The homogenate was centrifuged at 1000 × g, and washed at least four times with 50 mM phosphate buffer [6]. The pellet was collected and used as a cell wall fraction.
POD ionically bound to the cell wall was ex-tracted with 1 M NaCl. Cell walls were prepared as described above were incubated in 1 M NaCl for 2 h with shaking at 30°C and centrifuged at 1000 × g. The supernatant was used for enzyme assays. Guaiacol was used as substrate to deter-mine cell wall POD. The assay medium contained 0.05 M phosphate buffer (pH 5.8), 7.2 mM guaia-col, 11.8 mM H2O2and 0.1 ml enzyme extract in a
final assay volume of 3.0 ml. The reaction was
initiated by the addition of H2O2 and the change
in absorbance at 470 nm was measured [20]. Activ-ity was calculated using the extinction coefficient (26.6 mM− 1 cm− 1 at 470 nm) for tetraguaiacol
[20]. One unit of cell wall POD was defined as the amount of enzyme that causes the formation of 1 mmol tetraguaiacol per min. Ferulic acid POD (FPOD) was assayed according to Sanchez et al. [21]. The oxidation of ferulic acid was measured spectrophotometrically following the absorance decrease at 310 nm in a reaction mixture contain-ing 1.35 ml Na-phosphate buffer (0.2 M, pH 5.8), 0.5 ml ferulic acid (240mM), 0.5 ml H2O2(3 mM)
and 0.15 ml enzyme extract. One unit of FPOD was defined as a decrease of 1 A310 min− 1.
NADH-POD, which catalyzes the reduction of O2to H2O2using NADH as electron donor,
activ-ities in the ionic cell wall fraction were determined according to the method of Ishida et al. [22]. The assay mixture contained 50mM NADH in Na-ace-tate buffer (30 mM, pH 6.5), 5 mM MnCl2and 20
mM p-coumaric acid. The reaction was started by adding the enzyme, and the decrease of ab-sorbance at 340 nm by oxidizing NADH was measured at 25°C. One unit of NADH-POD was defined as a decrease of 1 A340 min− 1.
DAO activities in the ionic cell wall fraction were measured by the method of Naik et al. [23]. The reaction mixture contained 50 mM phosphate buffer (pH 7.8), 10 mM putrescine (Put), 0.1 mM pyridoxal phosphate and enzyme extract in a total volume of 4 ml. After incubation at 30°C for 1 h the reaction was terminated using 1 ml 20% (w/v) trichloroacetic acid. After 30 min, the incubation mixture was centrifuged at 5000 × g for 15 min. One ml of ninhydrin mixture (250 mg ninhydrin in 6 ml acetic acid and 4 ml phosphoric acid) was added to the supernatant. Colour was developed at 100°C for 30 min. After adding 1 ml of acetic acid absorbance was measured at 510 nm. In controls, trichloroacetic acid added prior to the enzyme solution. One unit of DAO was defined as an increase of 1 A510 per h.
The H2O2level was colorimetrically measured as
described by Jana and Choudhuri [24]. H2O2 was
extracted by homogenizing 10 roots with 3 ml of phosphate buffer (50 mM, pH 6.8) containing catalase inhibitor hydroxylamine (1 mM). The ho-mogenate was centrifuged at 6000 × g for 25 min. To determine H2O2 levels, 3 ml of extracted
in 20% (v/v) H2SO4 and the mixture was then
centrifuged at 6000 × g for 15 min. The intensity of the yellow colour of the supernatant was mea-sured at 410 nm. H2O2 level was calculated using
the extinction coefficient 0.28 mmol− 1
cm− 1
. Proline was extracted and its concentration de-termined by the method of Bates et al. [25]. Roots were homogenized with 3% sulfosalicylic acid and the homogenate was centrifuged. The supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h and then absorbance at 520 nm was determined. Proline level was calculated using the extinction coefficient 3.24 mmol− 1 cm− 1.
Ammo-nium was extracted by homogenizing roots in 0.3 mM sulphuric acid (pH 3.5). The homogenate was centrifuged for 10 min at 39 000 × g, and the supernatant was used for determination of ammo-nium as described by Lin and Kao [26].
For determinations of Put, spermidine (Spd) and spermine (Spm), roots were homogenized in 5% (v/v) perchloric acid. Levels of Put, Spd and Spm were determined using HPLC after benzola-tion as described previously [27].
For all measurements, each treatment was re-peated four times. All experiments described here were repeated at least three times. Similar results and identical trends were obtained each time. The data reported here are from a single experiment.
3. Results and discussion
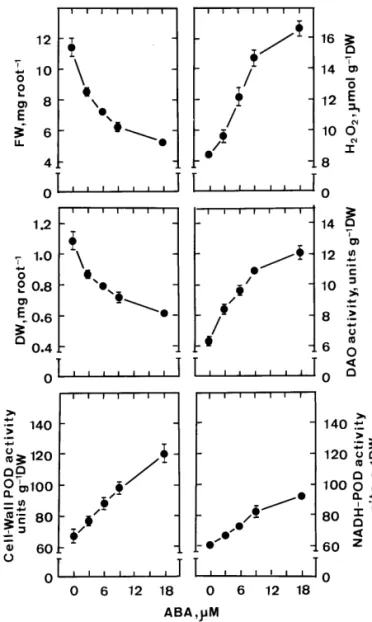
Root growth was followed by measuring FW and DW of roots. Fig. 1 shows the effect of ABA on root growth of rice seedlings. Increasing con-centrations of ABA from 3 to 18mM progressively decrease root growth. The reduction of root growth with increasing ABA concentrations is cor-related with an increase in cell-wall POD activity. Although there are some reports indicating that ABA may promote root growth [11 – 13], the re-sults of the present investigation are consistent with the generally accepted idea that ABA is a potent root growth inhibitor.
A key role of cell-wall POD in the stiffening of the cell wall through formation of cross-links be-tween wall polymers and consequently, in the re-duction or cessation of cell growth has been proposed [1]. Guaiacol was used as a substrate for cell-wall POD reported in Fig. 1. However, guaia-col is not the natural substrate in the formation of
cross-links between wall polymer. The polysaccha-rides of plant cell wall contain hydroxycinnamic acids that are presented as an ester-linked side chain. Ferulic acid has been identified as being ester linked to arabinoxylans in cell walls of monocotyledonous plants [28,29]. A key role in the cell-wall stiffening of dimerization of ferulic acid catalyzed by cell-wall POD has been pro-posed [21]. Although we do not know the con-stituents of polysaccharides of the cell wall in rice seedling roots, it is most likely that ferulic acid is also present in the cell wall of rice roots used in the present investigation. If this is indeed the case, then ABA is expected to increase the POD activity against ferulic acid (ferulic acid peroxidase or
Fig. 1. Effects of ABA on root growth, cell wall POD activities, H2O2levels, diamine oxidase (DAO) activities and
NADH peroxidase (NADH-POD) activities in roots of rice seedlings. Root growth, H2O2 levels and enzyme activities
were determined after 5 days of treatment. Vertical bars represent standard errors (n = 4).
Fig. 2. Effects of ABA on ferulic acid peroxidase (FPOD) activities in roots of rice seedlings. FPOD activities were determined after 5 days of treatment. Vertical bars represent standard errors (n = 4).
were found to increase with increasing ABA con-centrations in rice seedling roots (Fig. 1). Diamine oxidase (DAO), which is involved in polyamine catabolism, oxidizes Put with the formation of D1
-pyrroline together with H2O2 and ammonia
[32]. DAO activity is mainly localized in cell walls [33] and perhaps plays a role in regulating Put levels [32] or providing H2O2 required for
peroxi-dation reactions that occur in the cell walls for the formation of cross-links [33]. It seems that DAO is another source leading to H2O2 generation in
ABA-inhibited root growth of rice seedlings. To test this, we determined the activities of DAO in rice seedling roots in response to various concen-trations of ABA (Fig. 1). As expected, increasing concentrations of ABA from 3 to 18 mM progres-sively increased DAO activities. This result is con-sistent with the observations that ABA treatment decreases Put level but has no effect on Spd and Spm levels (Fig. 3).
It is known that ammonium strongly inhibits the growth of many plants [34]. Exogenous appli-cation of NH4Cl was also found to reduce root
growth [26] and to increase ionically bound POD FPOD). Fig. 2 shows that the reduction of root
growth with increasing ABA concentrations is cor-related with an increase in FPOD activity.
The results reported in the Figs. 1 and 2 seem to suggest that ABA-induced inhibition in root growth of rice seedlings is due to cell-wall stiffen-ing process catalyzed by POD. H2O2is a necessary
substrate for POD [1,5,16]. H2O2has been
demon-strated to cause a rapid cross-links of cell wall polymers [5,16,19]. If POD regulates cell-wall stiff-ening by catalyzing the oxidative cross-linking of cell wall polymers, there must be a sufficient sup-ply of H2O2. Thus, it is of great interest to know
whether ABA increases the level of H2O2 in roots
of rice seedlings. Fig. 1 shows that increasing concentration of ABA from 3 to 18 mM progres-sively increased H2O2levels in roots. Schopfer [17]
reported that in the hypocotyl of sunflower and cucumber seedlings the light-mediated inhibition of elongation growth was correlated with a strong increase in H2O2 in the epidermis and in the
vascular bundle. In a recent work, we demon-strated that exogenous application of H2O2
re-sulted in an inhibition of root growth of rice seedlings [30]. These results are in agreement with those of Schopfer [16] who demonstrated that H2O2 inhibited auxin-mediated growth of maize
coleoptile segments [16]. Our results seem to sug-gest POD-mediated cell-wall stiffening of rice roots is involved in ABA-inhibited root growth. One of potential sources of H2O2in plant cells is
cell wall- or membrane-bound NADH-POD are responsible for H2O2 generation [18,31]. As
ex-pected, increasing cell-wall NADH-POD activities
Fig. 3. Effects of ABA on the levels of polyamines in roots of rice seedlings. Putrescine (Put), spermidine (Spd) and sper-mine (Spm) were detersper-mined after 5 days of treatment. Verti-cal bars represent standard errors (n = 4).
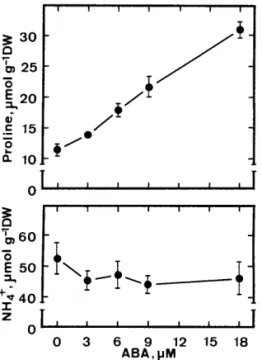
Fig. 4. Effects of ABA on the levels of proline and ammo-nium in roots of rice seedlings. Proline and ammoammo-nium were determined after 5 days of treatment. Vertical bars represent standard errors (n = 4).
increases proline level but had no effect on ammo-nium level. In our unpublished data, we have shown that exogenous proline application resulted in a reduction of root growth, an increase in H2O2
level, cell-wall POD and FPOD activities in roots of rice seedlings. The observations that rice seedlings fed with proline, which resulted in an increase in POD activity extracted from the cell wall and H2O2level in roots, reduced root growth
in the same way that ABA did, further support that cell-wall stiffening may be involved in the regulation of root growth reduction of rice seedlings exposed to ABA.
To test the causal relationship among root growth reduction, cell wall POD and FPOD activ-ities, H2O2 level, and cell wall DAO and
NADH-POD activities caused by ABA, 2-day-old seedlings were transferred to distilled water and ABA, respectively, for 4, 8, 12 and 16 h. Changes in root growth, cell wall POD and FPOD activi-ties, H2O2 level, and cell wall DAO and
NADH-POD activities were then monitored. As indicated in Table 1, an increase in cell wall POD and FPOD activities and H2O2 level preceded
inhibi-tion of root growth caused by ABA. Clearly, the links between ABA treatment, cell wall POD and FPOD, H2O2and root growth are well established.
The observations that an increase in DAO and NADH-POD activities coincides with an increase in H2O2 level in roots caused by ABA (Table 1)
activity [7] of rice seedlings. Previously, we also reported that exogenous application of proline resulted in an inhibition of root growth of rice seedlings and an increase in ionically bound POD [7]. Thus, it would be of great interest to under-stand whether ABA treatment resulted in an accu-mulation of proline and ammonium in roots of rice seedlings. Fig. 4 shows that ABA treatment
Table 1
Changes in root growth, cell wall POD activity, ferulic acid peroxidase (FPOD) activity, H2O2 level, diamine oxidase (DAO)
activity and NADH peroxidase (NADH-POD) activity in roots of rice seedlings treated with ABAa
Time, hours 0 4 8 12 16 6.4890.31 6.6590.11 6.6190.22 FW (mg root−1) H 7.3990.14 2O 7.7890.22 6.4590.14 6.5890.19 6.8290.15 ABA 7.0890.06 0.7890.02 0.7490.01 0.6690.02 0.6790.03 DW (mg root−1) H 0.6490.03 2O 0.6490.01 0.6690.02 0.6890.02 ABA 0.7190.01 H2O 43.1891.46 44.3292.37
Cell Wall POD (units g−1DW) 42.9092.77 46.1891.98 52.1993.71
60.4393.18 54.3092.11 49.1493.32 43.1293.48 ABA H2O 20494 224917 FPOD (units g−1DW) 21899 216922 247910 22397 259911 ABA 26598 312911 H2O 6.4690.21 6.6090.33 H2O2(mmol g−1DW) 6.6390.26 7.0890.44 7.6890.20 ABA 6.8690.41 7.1890.19 7.9490.31 8.6790.45 5.7190.35 H2O 7.0590.31 DAO (units g−1DW) 5.8890.29 6.1390.18 6.6790.32 7.9690.35 7.5490.22 6.8990.34 ABA 6.1490.37 H2O 57.7793.21 56.2993.15 55.4792.27 58.7093.35 65.4393.19 NADH-POD (units g−1DW) 74.2192.94 66.9892.16 62.2192.77 54.6792.35 ABA
suggest that DAO and NADH-POD are the sources for the generation of H2O2in the cell wall.
NADH-POD catalyzes the reduction of O2 to
H2O2 using NADH. The question now arises by
which mechanism NADH is provided. Gross [31] provided evidence to show that cell wall-bound malate dehydrogenase was involved in the produc-tion of NADH. However, we have not succeeded in detecting cell wall-bound malate dehydrogenase in roots of rice seedlings. Frahry and Schopfer [35] also failed to detect cell wall-bound malate dehy-drogenase activity in soybean roots. It seems that NADH-POD plays minor role, if any, in provid-ing H2O2 in ABA-treated roots of rice seedlings.
Since H2O2can rapidly pass from the cytoplasm to
the cell wall [36], a cytoplasmic origin of released H2O2 cannot be ruled out.
In conclusion, the data reported in the present investigation support the involvement of cell-wall stiffening in growth reduction in rice seedling roots exposed to ABA.
Acknowledgements
This work was supported by the National Sci-ence Council of the Republic of China (NSC 89-2313-B-002-009).
References
[1] S.C. Fry, Cross-linking of matrix polymers in the grow-ing cells of angiosperms, Annu. Rev. Plant Physiol. 37 (1986) 165 – 186.
[2] M.A. Bacon, D.S. Thompson, W.J. Davies, Can cell wall peroxidase activity explain the leaf growth response of Lolium temulentum L. during drought?, J. Exp. Bot. 48 (1997) 2075 – 2085.
[3] S.L. Chen, C.H. Kao, Cd induced changes in proline level and peroxidase activity in roots of rice seedlings, Plant Growth Regul. 17 (1995) 67 – 71.
[4] R. Goldberg, M. Liberman, C. Mathieu, M. Pierron, A.M. Catesson, Development of epidermal cell wall peroxidases along the mung been hypocotyl: possible involvement in the cell wall stiffening process, J. Exp. Bot. 193 (1987) 1378 – 1390.
[5] M. Hohl, H. Greiner, P. Schopfer, The cryptic growth response of maize coleoptile and its relationship to H2O2-dependent cell wall stiffening, Physiol. Plant 94
(1995) 491 – 498.
[6] T.-M. Lee, Y.-H. Lin, Changes in soluble and cell wall-bound peroxidase activities with growth in anoxia-treated rice (Oryza sati6a L.) coleoptiles and roots, Plant Sci. 106 (1995) 1 – 7.
[7] C.C. Lin, C.H. Kao, NaCl induced changes in ionically bound peroxidase activity in roots of rice seedlings, Plant Soil 216 (1999) 147 – 153.
[8] J.M. MacAdam, R.E. Sharp, C.J. Nelson, Peroxidase activity in the leaf elongation zone of tall fescue. II. Spatial distribution of apoplastic peroxidase activity in genotypes differing in length of elongation zone, Plant Physiol. 99 (1992) 879 – 885.
[9] X. Zheng, R.B. van Huystee, Peroxidase-regulated elon-gation of segments from peanut hypocotyls, Plant Sci. 81 (1992) 47 – 56.
[10] R.A. Creelman, Abscisic acid physiology and biosynthe-sis in higher plants, Physiol. Plant 75 (1989) 131 – 136. [11] P.-E. Pilet, D.W Barlow, The role of abscisic acid in root
growth and gravireaction: A critical review, Plant Growth Regul. 6 (1987) 217 – 265.
[12] I.N. Saab, R.E. Sharp, J. Pritchard, G.S. Voetberg, Increased endogenous abscisic acid maintains primary root growth and inhibits shoot growth of maize seedlings at low water potentials, Plant Physiol. 93 (1990) 1329 – 1336.
[13] R.E. Sharp, Y. Wu, G.S. Voetberg, I.N. Saab, M.E. Lenoble, Confirmation that abscisic acid accumulation is required for maize primary root elongation at low water potentials, J. Exp. Bot. 45 (1994) 1743 – 1751.
[14] G. Zocchi, P. De Nisi, Physiological and biochemical mechanisms involved in the response to abscisic acid in maize coleoptiles, Plant Cell Physiol. 37 (1996) 840 – 846. [15] F.Y. Tsai, C.C. Lin, C.H. Kao, A comparative study of the effects of abscisic acid and methyl jasmonate on seedling growth of rice, Plant Growth Regul. 21 (1997) 37 – 42.
[16] P. Schopfer, Hydrogen peroxide-mediation cell-wall stiffening in vitro in maize coleoptiles, Planta 199 (1996) 43 – 49.
[17] P. Schopfer, Histochemical demonstrations and localiza-tion of H2O2 in organs of higher plants by tissue printing on nitrocellulose paper, Plant Physiol. 104 (1994) 1269 – 1275.
[18] E.F. Elstner, A. Heupel, Formation of hydrogen perox-ide by isolated cell walls from horseradish (Armoracia
lapathifolia Gilib), Planta 193 (1976) 283 – 289.
[19] D.J. Bradley, P. Kjellbom, C.J. Lamb, Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: a novel, rapid defense response, Cell 70 (1992) 21 – 30.
[20] M. Kato, S. Shimizu, Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing to-bacco leaves; phenolic-dependent peroxidative degrada-tion, Can. J. Bot. 65 (1987) 729 – 735.
[21] M. Sanchez, M.J. Pena, G. Revilla, I. Zarra, Changes in dehydrodiferulic acids and peroxidase activity against ferulic acid associated with cell walls during growth of
Pinus pinaster hypocotyl, Plant Physiol. 111 (1996) 941 –
946.
[22] A. Ishida, K. Ookubo, K. Ono, Formation of hydrogen peroxide by NAD (P) H oxidation with isolated cell wall-associated peroxidase from cultured liverwort cells,
Morchantia polymorpha L, Plant Cell Physiol. 28 (1987)
[23] B.I. Naik, R.G. Goswami, S.K. Srivastawa, A rapid and sensitive colorimetric assay of amine oxidase, Anal. Biochem. 111 (1981) 146 – 148.
[24] S. Jana, M.A. Choudhuri, Glycolate metabolism of three submerged aquatic angiosperms during aging, Aquat. Bot. 12 (1981) 345 – 354.
[25] L.S. Bates, R.P. Waldren, I.D. Teare, Rapid determina-tion of free proline for water stress studies, Plant Soil 39 (1973) 205 – 207.
[26] C.C. Lin, C.H. Kao, Disturbed ammonium assimilation is associated with inhibition of roots in rice seedlings caused by NaCl, Plant Growth Regul. 18 (1996) 233 – 238.
[27] C.T. Chen, C.H. Kao, Senescence of rice leaves XXX. Levels of endogenous polyamines and dark-induced senescence of rice leaves, Plant Cell Physiol. 32 (1991) 934 – 941.
[28] Y. Kato, D.J. Nevin, Isolation and identification of O - (5 - O - feruloyl -a-L- arabinofuranosyl) - (1 - 3) - O -b-D
-xylopiranosyl-(1-4)-D-xylopyranose as a component of
Zea shoot cell walls, Carbohydr. Res. 154 (1985) 177 –
187.
[29] R.D. Hartley, W.H. Morrison, D.S. Himmelsbach, W.S. Borneman, Cross-linking of cell wall phenolic ara-binoxylans in graminaceous plants, Phytochemistry 29 (1990) 3705 – 3709.
[30] L.-M. Chen, C.C. Lin, C.H. Kao, Copper toxicity in rice seedlings: changes in antioxidative enzyme activities, H2O2 level, and cell wall peroxidase activity in roots,
Bot. Bull. Acad. Sin. 41 (2000) 99 – 103.
[31] G.G. Gross, Cell wall-bound malate dehydrogenase from horseradish, Phytochemistry 16 (1977) 319 – 321. [32] T.A. Smith, Di- and polyamine oxidases of higher
plants, Biochem. Soc. Trans. 13 (1985) 319 – 322. [33] R. Angelini, R. Federico, Histochemical evidence of
polyamine oxidation and hydrogen peroxide production in the cell wall, J. Plant Physiol. 135 (1989) 212 – 217. [34] R.I. Haynes, K.M. Goh, Ammonium and nitrate
nutri-tion of plants, Biol. Rev. 53 (1978) 465 – 510.
[35] G. Frahry, P. Schopfer, Hydrogen peroxidase produc-tion by roots and its stimulaproduc-tion by exogenous NADH, Physiol. Plant 103 (1988) 395 – 404.
[36] A.C. Allan, R. Fluhr, Two distinct sources of elicited reactive oxygen species in tobacco epidermal cells, Plant Cell 9 (1997) 1559 – 1572.