Original Paper
J Biomed Sci 2004;11:928–939 DOI: 10.1159/000081840
Induction of Apoptosis in MCF-7 Human
Breast Cancer Cells by Phytochemicals
from
Anoectochilus formosanus
Lie-Fen Shyur
aChih-Huai Chen
b, cChiu-Ping Lo
aSheng-Yang Wang
aPei-Ling Kang
aShow-Jane Sun
aC. Allen Chang
cChi-Meng Tzeng
bNing-Sun Yang
aaInstitute of BioAgricultural Sciences, Academia Sinica, and bDepartment of R & D, U-Vision Biotech Inc., Taipei,
and cDepartment of Biological Science and Technology, National Chiao Tung University, Hsinchu, Taiwan, ROC
Received: February 19, 2004 Accepted: June 29, 2004
Key Words
Anoectochilus formosanus Hayata (Orchidaceae)W
ApoptosisW Fas ligandW Functional genomicsW MCF-7
cells
Abstract
The antitumor activity of Anoectochilus formosanus Hayata (Orchidaceae), a popularly used folk medicine in the treatment of cancers in Asia, was investigated in MCF-7 human mammary carcinoma cells. Plant extracts of A. formosanus were observed to induce apoptosis of MCF-7 cells as evidenced by cell-morphological changes, an early redistribution of plasma membrane phosphatidylserine, and DNA content distribution stud-ies. Bioactivity-guided fractionation of A. formosanus extracts produced a specific ethyl acetate (EA)-parti-tioned fraction in which apoptotic activity was enriched. The chemical profile and candidate index compounds of the active EA fraction were obtained using HPLC and var-ious spectral analyses. Western blot analysis showed that upon treatment of MCF-7 cells with the EA fraction, cleavage of pro-caspases-8, -9, and -7, and
poly(ADP-ribose) polymerase as well as significant release of mito-chondrial cytochrome c into the cytosol were readily observed. Flow cytometry showed that the Fas ligand protein was overexpressed in EA-treated MCF-7 cells. Functional genomic studies indicated that specific genes related to cytoskeleton rearrangement, apoptotic signal transduction, and various transcription factors were dif-ferentially regulated in EA-treated MCF-7 cells. Putative apoptotic signaling pathways of MCF-7 cells in response to the EA extract of A. formosanus are proposed.
Copyright © 2004 National Science Council, ROC and S. Karger AG, Basel
Introduction
Apoptosis, a form of programmed cell death, is either developmentally regulated or activated in response to spe-cific extracellular and intracellular stimuli or cell injury [18]. In mammalian cells, apoptosis can be triggered by members of the Fas and TNF receptor families [14]. Recent studies have shown that apoptosis often involves disruption of mitochondrial membrane integrity, result-ing in the release of cytochrome c from mitochondria into the cytosol, and this activity serves as a decisive factor for the onset of the cell death process [27]. A number of other cellular mechanisms are also known to be involved in the Chih-Huai Chen and Chiu-Ping Lo contributed equally to this work.
process of apoptosis, including the regulation of biochem-ical activities of a family of aspartate-specific cysteine proteases, known as caspases. Activated caspases can spe-cifically cleave cellular death substrates and cause bio-chemical and morphological changes leading to apoptosis [6].
Recent advances in understanding the modes of action of various anticancer agents indicate that regardless of the diverse chemical nature of anticancer drugs, most of them elicit apoptosis in test tumor cells [7, 25]. For example, taxol has been demonstrated to induce tumor cell apop-tosis via a mitochondrial-dependent pathway [9]. Also, some potent chemopreventive agents, such as sulindac and other nonsteroidal anti-inflammatory drugs, induce apoptosis in colon tumors, leading to the prevention of cancer [21]. Therefore, the discovery and use of apopto-sis-inducing molecules and the evaluation of their mecha-nisms of action should aid in the therapeutic manipula-tion of cancers and also help to understand the intracellu-lar pathways of apoptosis in cancerous cells.
Extracts of Anoectochilus formosanus are commonly used in herbal therapies and drink supplements adminis-tered to patients with cancer in Asian countries. However, the pharmacological activities of this empirically used medicinal herb remain to be scientifically elucidated. Recently, we reported that plant extracts of A. formosanus exhibit strong antioxidant activities, indicating the poten-tial for its development into a chemopreventive agent [24]. In this report, we observed that an enriched ethyl acetate (EA) fraction from the hot water extract of A.
for-mosanus conferred significant antiproliferative effects
and induced apoptosis in the human MCF-7 breast cancer cell line. A phytochemical profile of the EA-bioactive frac-tion was obtained using HPLC, IR, LC-mass, and one-and two-dimensional NMR analyses.
Rapid analysis of differential gene expression on a genome-wide scale became feasible with the development of DNA microarray technology [8]. This strategy offers a systematic approach to search for effective targets for drug discovery and diagnostics [4]. Results of human cDNA microarray analysis in this report revealed that a specific set of apoptosis-related genes in MCF-7 cells was highly responsive to treatment with the EA fraction of A.
formosanus. Our findings indicate that the EA fraction of A. formosanus induces apoptosis in MCF-7 cells via a Fas
ligand-mediated mechanism, in which the gene and pro-tein expressions of cytochrome c are dramatically in-creased, and the apical and executive caspases (caspases-8, -9, and -7) are activated.
Materials and Methods
Chemicals
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide, MTT, N)-bis(2-ethanesulfonic acid, PIPES, 3-[(3-cholamidopropyl)-dimethylammonio]-1-propane-sulfonate, CHAPS, N-(2-hydroxyeth-yl) piperazine-N)-(2-ethanesulfonic acid), HEPES, and plumbagin were purchased from Sigma (St. Louis, Mo., USA). Agarose was obtained from Bio-Rad (Hercules, Calif., USA), curcumin was from ACROS ORGANIES (Morris Plains, N.J., USA), and RPMI 1640 growth medium from GIBCOTM (Invitrogen, N.Y., USA). All other
chemicals and solvents used in this study were of reagent or HPLC grade.
Plant Materials
Fresh A. formosanus Hayata (Orchidaceae) plants in the flower-ing stage were purchased from a reputable Anoectochilus Cultural Station at Puli, Nantou County, central Taiwan, as officially recom-mended by Dr. Hsin-Sheng Tsay of the Taiwan Agricultural Re-search Institute, Taichung, Taiwan, ROC. The authenticity of these A. formosanus plants was validated by the specific morphological and anatomical features of the flowers [12]. Voucher specimens (No. 642008010) were deposited at the Herbarium, Institute of Botany, Academia Sinica, Taipei, Taiwan, ROC.
Preparation and Fractionation of Plant Extracts
Fresh whole plants of A. formosanus (1 kg) were extracted twice, by boiling for 2 h with 3 volumes of distilled water. The total crude plant extract (AF-hot, F49 g dry weight) was collected by centrifuga-tion at 24,000 g for 20 min at 4° C, and then lyophilized to dryness. A stepwise ethanol fractionation procedure, using 50, 75, and 87.5% (v/v) ethanol, respectively, was employed to obtain the subfractions AH-I, AH-II, AH-III, and AH-sup from the AF-hot crude extract. The AH-sup (19.3 g), i.e. the remaining soluble fraction of the AF-hot extract treated with 87.5% ethanol, was then further subjected to dif-ferential solvent partitioning using EA, followed by butanol (BuOH), to yield the EA (" 1.2 g), BuOH (" 3.4 g), and water (" 14.4 g) subfrac-tions, respectively.
Cell Culture
MCF-7 cells obtained from the American Type Culture Collec-tion (ATCC, Manassas, Va., USA) were grown in RPMI 1640 medium supplemented with 10% fetal bovine serum, 100 U/ml peni-cillin, and 100 Ìg/ml streptomycin. In initial comparative studies, 10% fetal bovine serum was tested for substitution with serum replacement 2 (Sigma). H184B5F5/M10 (ATCC) cells, a non-can-cerous human mammary epithelial cell line, were grown in 90% MEM (GIBCOTM) and 10% fetal bovine serum as suggested by the
instruction manual provided by ATCC. All experiments were carried out at 37° C and 5% CO2 on confluent cells in the medium.
Cell Viability Assay
MCF-7 cells (5 ! 103 to 1 ! 104) were incubated with or without
test plant extracts in medium in 96-well plates for 1–3 days. Cell via-bility was assayed using the MTT colorimetric dye reduction meth-od. Survival of MCF-7 cells after treatment with plant extracts was calculated using the following formula: viable cell number (%) = [OD570 (treated cell culture)/OD570 (control, untreated cell culture)]
! 100. A 3H-thymidine incorporation assay was also employed to
A. formosanus, following the experimental protocol described by Seufferlein and Rozengurt [18].
Fluorescent Microscopy and Flow-Cytometric Analyses of Tumor Cells
The Apoalert Annexin V Apoptosis kit (Clontech, Palo Alto, Cal-if., USA) was used to analyze apoptotic activity in MCF-7 carcinoma cells. Test cells were treated for 24 h with the EA fraction (1 mg/ml) or the vehicle control (0.1% DMSO), washed with PBS buffer, resus-pended in binding buffer, and then stained with Annexin V-EGFP and/or propidium iodide (PI) for 15 min in the dark. Treated cells (F2 ! 105) were cytospun at 800 rpm for 5 min and viewed using a
Nikon (Melville, N.Y., USA) ECLIPSE E800 phase-contrast fluores-cent microscope.
MCF-7 cells (1 ! 106 cells), treated with or without plant
extracts, were harvested and fixed with 1 ml ice-cold 70% ethanol at 4° C for 2 h. Total cellular DNA was stained for 30 min in the dark with 50 Ìg/ml of a PI solution containing 100 Ìg/ml RNase A in PBS buffer. The number of cells with a sub-G1 DNA content divided by the number of cells with the DNA content in the whole-cell cycle was taken as a measure of the percentage of the apoptotic cell population, using a Coulter EPICS XL flow cytometer (Beckman/Coulter, Dur-ham, N.C., USA). Data were generated from samples with at least 10,000 cells per assay and analyzed with the DNA MultiCycle pro-gram (Beckman/Coulter).
Western Blot Analysis
MCF-7 cells (2 ! 106) were washed with PBS once and lysed in
100 Ìl lysis buffer [20 mM PIPES (pH 7.0), 10 mM NaCl, 1 mM EDTA, 0.1% CHAPS, 10% sucrose, 10 mM DTT, 5 mM HEPES, 0.05% (v/v) Triton X-100, 1 mM MgCl2, 2.5 mM EGTA, and 0.44%
(w/v) ß-glycerol-phosphate] plus 2 Ìl protease inhibitor cocktail set III (Calbiochem, San Diego, Calif., USA) for 20 min on ice and extensively vortex mixed every 5 min. After centrifugation, the pro-tein concentration in the supernatant was determined using a dye-binding assay (Bio-Rad). Protein samples (40 Ìg) were separated in a 5–20% gradient mini-SDS-polyacrylamide gel and transferred onto a PVDF membrane (Millipore, Bedford, Mass., USA). Membranes were first blocked with 3% (w/v) non-fat dry milk in TBS buffer (10 mM Tris, pH 7.5, and 100 mM NaCl) for 30 min and then incu-bated with specific primary antibodies at 4° C overnight with agita-tion. Mouse anti-glyceraldehyde-3-phosphate dehydrogenase (GA3PDH; Biogenesis, Kingston, N.H., USA), rabbit anti-NF-ÎB (Oncogene Research Products, Boston, Mass., USA), mouse anti-poly(ADP-ribose) polymerase (PARP; Transduction Laboratories, Lexington, Ky., USA), mouse anti-cytochrome c (Pharmingen, San Diego, Calif., USA), mouse anti-procaspases-2, -7, and -9 (Pharming-en), and rabbit anti-procaspase-8 (BD Research Products, San Diego, Calif., USA) antibodies were individually used as primary anti-bodies. After washing with TBST buffer (0.1% Tween 20 in TBS buffer), the blotting membrane was treated with alkaline-phospha-tase-conjugated secondary antibodies at room temperature for 2 h. Immunoreactive proteins were visualized by the enhanced chemilu-minescence system according to the manufacture’s protocol (Amers-ham Pharmacia Biotech, Amers(Amers-ham Place, UK).
mRNA Preparation and cDNA Probe Generation
Total cellular RNA samples of MCF-7 cells treated with plant extracts or vehicle control (0.4% DMSO) for 3, 6, 9, 12, 18, and 24 h were prepared using the Trizol® reagent and further purified using
the RNeasy® kit (QIAGEN, Cologne, Germany). Biotin-labeled
cDNA probes were generated using the RT-PCR method described by Lau et al. [11].
Generation, Hybridization, and Colorimetric Signal Detection of a cDNA Microarray
Candidate cDNAs for generating cDNA microarrays were pre-pared from IMAGE consortium gene pools or self-constructed cDNA clones using PCR. The PCR reaction mixtures (100 Ìl) consisted of 10 U Taq polymerase (Merck, Darmstadt, Germany), 0.2 ÌM of the desired primers, 200 ÌM dNTPs, and 10 ng of template DNA. PCR was performed with a thermocycling program of 3 min at 95° C, fol-lowed by 40 cycles of 1 min at 94° C, 1 min at 50 ° C, and 2 min at 72 ° C on a PrimusHT thermocycler (MWG-Biotech, High Point, N.C., USA). The PCR products were then precipitated by 70% EtOH (Merck, Darmstadt, Germany), vacuum dried, and dissolved in steril-ized water to a final concentration of 1.1 Ìg/Ìl DNA ("108 molecules of
DNA/spot), before being spotted onto a nylon membrane. Conversion of coordinate and database constructions was established using Gene-TrendTM software (U-Vision, Taipei, Taiwan, ROC). Probe
hybridiza-tion and color development were adapted from the colorimetric system published elsewhere [11]. The PA5500TM arrayer and nanoliter
quan-titative aspirate-dispenser (nQUADTM) with microplate stacker
Pega-sysTM 150 (Cartesian Technologies, Research Triangle Park, N.C.,
USA) were the printing system for generating the DNA microarrays. A 3,000-dpi light scanner (Umax, Taipei, Taiwan, ROC) combined with Imagene® (Biodiscovery, Boston, Mass., USA) and QuantArray®
soft-ware, and statistical programs developed at U-Vision Biotech, Taipei, Taiwan, ROC (http://www.u-vision-biotech.com/) were used to per-form image and microarray data analyses.
Chemical Fingerprint Analysis of the EA Fraction of A. formosanus
Analytical HPLC was performed to analyze the chemical finger-print of the EA fraction from the A. formosanus extract. A Waters (Milford, Mass., USA) HPLC system equipped with a Waters 600 con-troller, Waters delta 600 pump, and 2487 duel Ï absorbance detector was used. A 5-Ìm Si-60 column (250 ! 10 mm, Merck) was employed with two solvent systems: n-hexane (A) and ethyl acetate (B). The elu-tion gradient profile was set as follows: 0–5 min, 70% A to B (isocrat-ic); 5–15 min, 70–30% A to B (linear gradient); 15– 20 min 30% A to B (isocratic), and 20–30 min 30–0% A to B, while the flow rate was set at 1.5 ml/min and the detector wavelength was set to 254 nm. Six pure compounds (1–6) eluted with retention times of 6.0 (1), 8.9 (2), 9.7 (3), 13.5 (4), 14.6 (5), and 18.0 (6) min, respectively, were collected for structural determination using various spectroscopic analyses. UV spectra of candidate index compounds were recorded with a Jasco V-550 spectrometer and IR spectra obtained from a Bio-Rad FTS-40 spectrophotometer. Electron-impact mass spectrometric (EIMS) and high-resolution EIMS data were collected with a Finnigan (Dreieich, Germany) MAT-958 mass spectrometer; NMR spectra were recorded with Bruker Avance (Bremen, Germany) 500 and 300 MHz FT-NMR spectrometers at 500 (1H) and 75MHz (13C). Profiling and
quantifica-tion of index compounds in the EA fracquantifica-tion were performed by HPLC. The peak areas corresponding to the contents of candidate index com-pounds in the chromatogram of a known concentration of the EA frac-tion were calculated based on a standard calibrafrac-tion curve of individu-al index compounds. Standard cindividu-alibration curves (peak areas vs. con-centrations) of compounds 2 and 4, ranging from 0.05 to 1 mg/ml, were first determined at OD254.
Results
Cytotoxic Effects of A. formosanus Extracts on MCF-7 Cells
The cytotoxic effects of plant extracts from A.
formo-sanus on the viability of MCF-7 cells were determined by
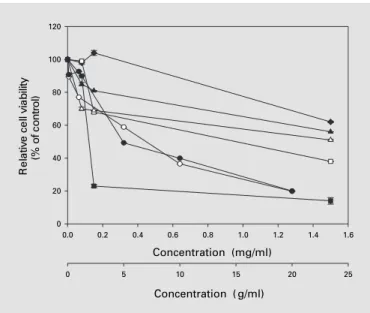
MTT assays. In setting up the test system, we observed a very similar pattern of cytotoxic effects for the total crude extract of the plant (AF-hot), regardless of whether the culture medium was supplemented with fetal bovine serum or serum substitute (data not shown), suggesting that components of fetal bovine serum did not affect the inhibition of MCF-7 cell proliferation conferred by the phytocompounds in A. formosanus extracts. We then observed that the AH-sup fraction showed the most sig-nificant cytotoxic effect on MCF-7 cells relative to the AF-hot, AH-I, AH-II, and AH-III fractions (data not shown). Less than 45% of tested cells were detected as being viable after treatment with 1.5 mg/ml AH-sup for 48 h. In contrast, more than 70% cell viability was observed in MCF-7 cells treated with AF-hot, I, AH-II, or AH-III using the same dosage. The AH-sup fraction was therefore further fractionated using EA and butanol partitions to obtain more enriched and more potent sub-fractions. As shown in figure 1, the EA fraction conferred the greatest cytotoxicity to MCF-7 cells, as compared to the AF-hot, AH-sup, BuOH, and water subfractions. Dos-ages of plant extract/fraction required to inhibit 50% of MCF-7 cell proliferation (IC50) were found to be 0.86 mg/
ml for AH-sup, 0.08 mg/ml for EA, 1.5 mg/ml for BuOH, and 2.7 mg/ml for the water fraction. Therefore, a 10-fold lower IC50 value for anti-MCF-7 cell proliferation was
detected for the EA fraction as compared to that of AH-sup. Similar results were obtained from the 3H-thymidine
incorporation assays (data not shown). These results strongly suggest that the cytotoxic effect of the A.
formo-sanus extract on MCF-7 cells was effectively enriched in
the EA fraction. We obtained very similar cytotoxicity results of EA fractions from different batch preparations (data not shown). In this study, two well-known phyto-compounds, namely curcumin and plumbagin, were em-ployed as positive controls in the anti-MCF-7 cell prolifer-ation assays, with data also shown in figure 1. Curcumin has been reported to be involved in anti-cell proliferation and induction of apoptosis in various types of tumor cells including MCF-7 cells [23] and human renal carcinoma Caki cells [26]. Plumbagin, a single-compound drug iso-lated from Plumbago rosea, is a known plant naphthoqui-none with demonstrated antitumor and antibacterial properties [10]. IC50 values for anti-MCF-7 cell
prolifera-Fig. 1. Antiproliferative effect of AF-hot (j), AH-sup ()), EA ($), BuOH (o), and water (d) fractions of A. formosanus extract on MCF-7 mammary carcinoma cells. Relative cell viability (%) was measured 48 h after treatment of test cells with or without (control) the indicated doses of plant extracts (0.075–1.5 mg/ml) or curcumin (1–20 Ìg/ml, [), using MTT assays. Cell viability of plumbagin-treated MCF-7 cells (P) was determined after 3 h of treatment at doses ranging from 0.1 to 20 Ìg/ml. Curcumin and plumbagin were used as positive controls in this experiment. Each data point repre-sents the mean B SE of 3 independent experiments.
Concentration (mg/ml) 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 yti li b ai v ll e c e vit al e R )l ort n o c f o %( 0 20 40 60 80 100 120 Concentration ( g/ml) 0 5 10 15 20 25
tion of curcumin and plumbagin were found to be 7.5 Ìg/ ml (48-hour treatment) and 5 Ìg/ml (3-hour treatment), respectively.
H184B5F5/M10 cells, a non-cancerous human mam-mary epithelial cell line, were employed to test the poten-tial cytotoxicity of the EA extract and curcumin using the MTT assay. We observed that the EA fraction exhibited an approximately 15-fold higher IC50 value (F1,200 Ìg/
ml) for the antiproliferative effect against H184B5F5/ M10 cells than that for anti-MCF-7 cell proliferation (80 Ìg/ml) after 48 h of treatment. The IC50 values of curcu-min for anti-MCF-7 and anti-H184B5F5/M10 cell prolif-eration were both determined to be approximately 7.5 Ìg/ ml after 48 h of treatment. These results indicate that cur-cumin conferred a lower specificity for discriminating between tumor and normal cell types as compared to that of the EA fraction of A. formosanus.
The EA Fraction of A. formosanus Extract Induces Apoptosis in MCF-7 Cells
Light microscopy was employed to examine the cellu-lar morphology of MCF-7 cells after treatment with A.
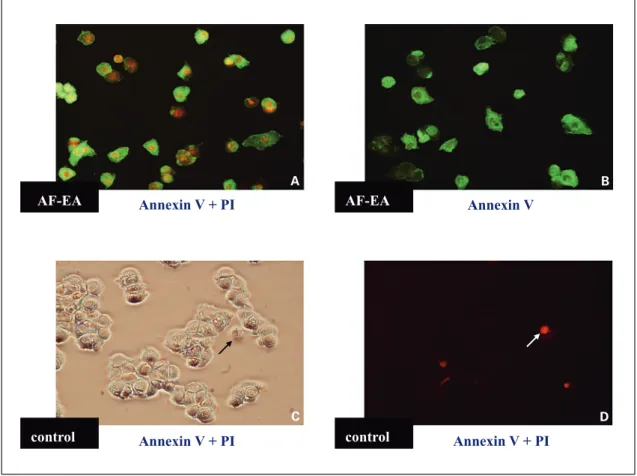
for-Fig. 2. Microscopic examination of normal versus EA fraction-treated MCF-7 cells in culture, as stained with Annex-in-V and PI. log phase MCF-7 cells in culture were treated with the EA fraction (1 mg/ml) for 24 h and stained with PI plus Annexin-V (A) or Annexin-V alone (B). C, D MCF-7 cells treated with 0.1% DMSO (vehicle control), and stained with PI plus Annexin-V. A, B, D Fluorescent photomicrographs taken with an Olympus PM-30 camera on a Nikon ECLIPSE E800 microscope (magnification: !200) using a B-2A filter (Ex 450–490). C Bright-field view of D.
AF-EA AF-EA control control Annexin V + PI Annexin V + PI Annexin V + PI Annexin V A C B D
mosanus extracts. In comparison to vehicle
control-treated cells, EA-control-treated MCF-7 cells showed cell shrink-age, cytoplasmic blebbing, and the formation of apoptotic bodies (data not shown). An early redistribution of plas-ma membrane phosphatidylserine was also readily de-tected in MCF-7 cells after treatment with the EA fraction (1 mg/ml) for 24 h, as determined using Annexin V-GFP and PI staining (fig. 2), suggesting that the EA fraction effectively induced apoptosis in MCF-7 cells. We further used Annexin V-GFP staining (Clontech Laboratories) and flow cytometric analysis to examine the early events of apoptosis in MCF-7 cells induced by the EA fraction. We observed that the population of Annexin-V-stained MCF-7 cells increased in a dose-dependent manner when treated with the EA fraction for 24 h at concentrations ranging from 50 to 1,000 Ìg/ml. Percentages of apoptotic
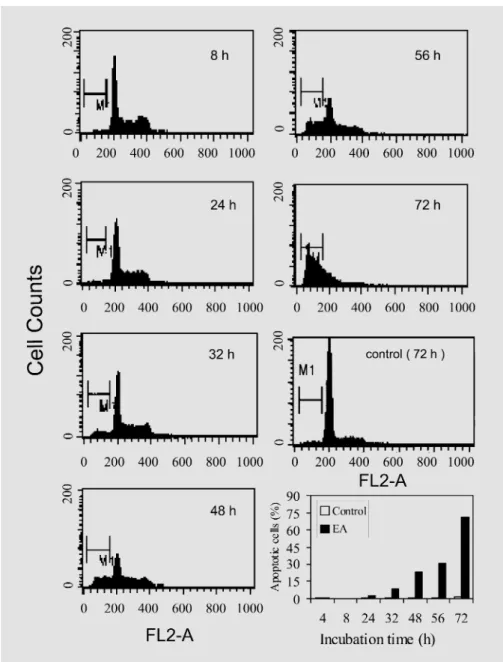
MCF-7 cells were determined to be 1.2, 2.9, 7, and 20% when test cells were treated with 50, 150, 500, and 1,000 Ìg/ml EA, respectively. Figure 3 shows the typical flow cytometric profiles of DNA content and cell cycle behavior of MCF-7 cells with or without treatment with the EA extract. After 48 h of treatment, approximately 25% of total MCF-7 cellular DNA was detected as apoptot-ic (determined from the sub-G1 peak), and the level rose to 71% 72 h after treatment. Cell cycle arrest activity was not detected in MCF-7 cells treated with the EA fraction.
The EA Extract Induces FasL Expression in MCF-7 Cells
It is known that expression of the Fas ligand (FasL) can effectively mediate apoptosis by its binding to the cognate receptor, Fas [14]. In this study, results obtained from
Fig. 3. Flow cytometric analysis of DNA content in normal and apoptotic MCF-7 cells, examined as a function of treatment time with the EA extract. MCF-7 cells were treated with either 0.4% DMSO (vehicle control) or 1 mg/ml of the EA fraction of A. formosanus extract, and later harvested at the indicated time points. M1 denotes the apoptotic peak.
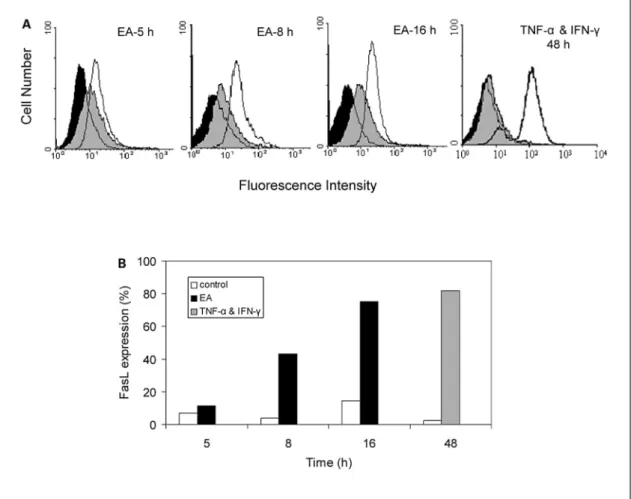
flow cytometric analysis using the FITC-labeled anti-human FasL IgG1 antibody demonstrated that the EA extract of A. formosanus significantly induced the expres-sion of FasL in MCF-7 cells. Upon treatment with EA for 5–16 h, FasL protein expression in MCF-7 cells drastical-ly increased from 10 to 76%, relative to non-treated cells (fig. 4A, B), in a time-dependent manner. The TNF-·-and IFN-Á-induced cell surface expressions of FasL in MCF-7 cells [15] were employed as reference controls in this experiment. Eighty percent FasL protein expression was observed in MCF-7 cells treated with both cytokines (fig. 4A, B).
Cellular Mechanisms of EA-Fraction-Induced Apoptosis in MCF-7 Cells
The potential cellular mechanisms of apoptosis in MCF-7 cells induced by the EA plant extract of A.
formo-sanus were characterized using immunoblot analysis.
Fig-ure 5 shows that significant increases in the levels of cyto-solic cytochrome c and proteolytic cleavage of pro-cas-pase-8 (F55 kDa) were detected within 16 h of MCF-7 cells being treated with the EA fraction (fig. 5A). Between 24 and 32 h after treatment, proteolytic processing of cas-pase-7 and -9, known to be involved in the early activa-tion cascade of apoptosis, was also effectively activated in
Fig. 4. EA fraction of A. formosanus induces FasL expression in MCF-7 cells. MCF-7 cells were untreated (isotype control) or treated with 1 mg/ml EA extract or 0.4% DMSO (vehicle control), for 5, 8, or 16 h. MCF-7 cells treated with 10 ng/ml TNF-· and 200 U/ml IFN-Á for 48 h were employed as a reference control of FasL expression. FasL expression was quantified using flow cytometry, by staining cells with mouse anti-human FasL IgG (clone NOK-1) or isotype control mouse IgG (MOPC-21), followed by FITC-conjugated rat
anti-mouse IgG. A Fluorescent intensities obtained from isotype control cells (black images), vehicle control cells (gray images), and EA- or TNF-·+IFN-Á-treated cells (white images), as analyzed using a Coulter EPICS XL flow cytometer (Beckman Coulter) with Expo XL 4 Cytometer software. B FasL protein expression (%) of vehicle control cells (white bars) and EA- (black bars) or TNF-·+IFN-Á-treated cells (grey bar), relative to isotype control cells.
MCF-7 cells. Upon EA treatment, the level of Rel A, a subunit (p65) of NF-ÎB known to be directly involved in the inhibition of apoptosis [13], was only slightly sup-pressed in MCF-7 cells (fig. 5A). Time-dependent proteo-lytic cleavage of PARP, another hallmark of apoptosis, was observed with an accumulation of a 28-kDa prod-uct and a concomitant disappearance of the full-size, 113-kDa enzyme molecule (fig. 5B). Bcl-2 and Bax pro-teins in MCF-7 cells were not found to be responsive to EA treatment (data not shown).
Gene Expression Profiles in MCF-7 Cells in Response to EA Treatment
Broad-range, 9,800-dot, duplicate spot-formatted sets of known human cDNA microarrays were employed to collect genomic information on the mechanisms of A.
for-mosanus extract-induced apoptosis in MCF-7 cells.
Re-sults from two duplicate experiments showed that the scatter plots derived from normalized signals at various treatment time points with EA were well correlated and reproducible. Along with statistical analysis of the annota-tion of specific genes and gene clusters, a summary of dif-ferentially expressed genes with at least 3-fold changes in
Fig. 5. Western blot analyses of caspases-7, -8, and -9, cytochrome c, NF-ÎB, GAPDH (A), and PARP (B) in MCF-7 cells treated with 0.4% DMSO (vehicle control) or with the EA fraction of A. formosanus. Aliquots of cell lysates were subjected to 5–20% SDS gradient gel electrophoresis. Detailed trans-blotting and immunostaining procedures are described in the Materials and Methods. Specific protein contents in control or EA-treated MCF-7 cells were quantified using densitometry. Relative protein contents (%) of EA-treated and control cells are presented as the protein level detected at various time points divided by the protein content at 4 h in either EA-treated or control cells.
Table 1. Overview of differentially expressed genes in MCF-7 cells responsive to the EA fraction of A. formosanus revealed by DNA microarray and bioinformatic analyses
Genes x-fold change
Apoptosis
c-Jun proto-oncogene (JUN) 11
14-3-3 epsilon 5
Cytochrome c 5
ADP-ribosyltransferase (NAD+; PARP) 5
Protein kinase Ca 4
FAST 3
Cell proliferation and growth arrest
Cyclin-dependent kinase inhibitor 2A 17
Protein phosphatase 2, regulatory subunit B (B56), Á isoforma 12
DNJ3/CPR3 mRNA 10
Regulator of mitotic spindle assembly 1 9
Cytoskeleton
Group-specific component (vitamin D binding protein) 4 Cell division cycle 42 (GTP binding protein, 25 kDa): CDC42 3 Actin-related protein 2/3 complex, subunit 2, 34 kDa: ARPC2 3 Signal transduction
Protein phosphatase 2, regulatory subunit B (B56), Á isoforma 12
Phosphotyrosine-independent ligand p62 for the Lck SH2 10
Tight junction protein 1 (zona occludens 1) 10
Tyrosine kinase (Tnk1) 6
Galactokinase 1 6
Diacylglycerol kinase ‰ 5
Protein tyrosine phosphatase, non-receptor type 12 5 ATP synthase lipid-binding protein P2 precursor 5
Phosphatidylinositol 4-kinase · (PI4K-·) 5
Protein kinase Ca 4
Phospholipase C ‰ 1 3
Type 1 inositol 1,4,5-trisphosphate receptor (IP3R-1) 3
Stress-activated protein kinase JNK2a 3
Stress-responsive serine/threonine protein kinase Krs-1a 5 Protein tyrosine phosphatase, receptor type, c polypeptide 3 Genes denoted in italics represent those that were downregulated in MCF-7 cells.
a Genes that can be subgrouped into more than 1 functional category.
MCF-7 cells is shown in table 1. In total, 52 genes of MCF-7 cells were found to exhibit more than a 3-fold change in specific activity when treated with the EA frac-tion, and 28 of them functionally relevant to apoptosis and signal transduction are listed in table 1. For example, the mRNA expression levels of cytochrome c, PARP, and TGF-ß superfamily proteins were upregulated, correlat-ing well with the observed activity of apoptosis in test cells. Gene expressions of caspases-2 and -8, however, were not found to be responsive to EA treatment. It has been demonstrated that activation of caspase-8 occurs through autoproteolytic processing after formation of
pro-caspase-8 protein oligomers, leading to the initiation of the caspase signaling cascade in apoptotic cells [5, 17, 29]. Our observations agree well with and are supported by the model in which the autoprocessing of caspase-8, without affecting its transcription level, initiates the proteolysis of the downstream executive caspases, e.g. caspase-7 in our case. The cell division cycle 42 (cdc42) gene was found to be downregulated after 9 h of EA treatment. More than a 5-fold increase in gene expression levels were detected for the regulator of mitotic spindle assembly 1, diacylglycerol kinase ‰, and c-Jun proteins, which were demonstrated to be correlated to morphological changes in apoptotic cells,
Fig. 6. Chemical fingerprints and candidate index compounds of the EA fraction of A. formosanus. The chromatogram of the EA fraction was obtained from a Si-60 HPLC system. Six compounds are suggested in this study as useful index or reference com-pounds, including (1) a long-chain aliphatic acid; (2) ·-amyrin trans-p-hydroxy cinamate; (3) ·-amyrin cis-p-hydroxy cinamate; (4) isorhamnetin; (5) kinsenone, and (6) p-hy-droxybenzyl alcohol.
as we suggested here for EA-treated MCF-7 cells. Fas-acti-vated serine/threonine kinase (FAST) has been shown to be activated during Fas-mediated apoptosis [22]. In our experiments, a 3-fold increase in FAST gene expression, as evaluated using the cDNA microarray as well as North-ern blot analyses, was observed in EA-treated MCF-7 cells (with 9 h of treatment).
Chemical Fingerprinting of the EA Fraction from
A. formosanus
Mass spectrometric and NMR analyses together with spectral data published from our laboratory and others [1, 24] have led to the identification of six compounds in the EA fraction of A. formosanus. These include a long-chain aliphatic acid (1), ·-amyrin trans-p-hydroxy cinamate (2), ·-amyrin cis-p-hydroxy cinamate (3), isorhamnetin (4), kinsenone (5), and p-hydroxybenzyl alcohol (6), as indi-cated by HPLC in figure 6. In this study, ·-amyrin
trans-p-hydroxy cinamate (2) and isorhamnetin (4) were chosen
as candidate index compounds of A. formosanus, and their contents were quantitatively determined in the bioactive EA fraction using HPLC. One gram of EA extract was quantitatively detected to contain 4.9 (0.49%)
and 52.3 mg (5.23%) of ·-amyrin trans-p-hydroxy cina-mate and isorhamnetin, respectively. The spectral data of ·-amyrin trans-p-hydroxy cinamate and isorhamnetin are summarized as follows.
·-Amyrin trans-p-hydroxy cinamate (2): white solid; mp 105–106°C; EIMS for C29H56O3 found 572.43; 1HNMR (CDCl
3): Û (ppm) 7.62 (d, J = 8.0), 7.45 (d, J =
8.0), 6.85 (d, J = 18.2), 6.32 (d, J = 18.2), 5.22 (m), 4.70 (m), 1.17 (s), 1.01 (s), 1.00 (s), 0.96 (s), 0.90 (s), 0.89 (s), 0.86 (s).
Isorhamnetin (4): yellow amorphous powder; EIMS for C16H12O7 found 312.6; 1HNMR (CDCl3): Û (ppm) 7.96
(d, J = 2.0), 7.56 (d, J = 8.0, 2.1), 7.04 (d, J = 7.8), 6.39 (d, J = 2.0), 6.20 (d, J = 2.0).
Discussion
In the present study, we demonstrate that the EA extract of A. formosanus inhibits proliferation of MCF-7 human mammary carcinoma cells through the induction of cell apoptosis. Cellular, biochemical, and functional genomic evidences indicate that treatment with the A.
for-mosanus extract may have caused transcriptional
disor-der of cytoskeletal biosynthesis, accompanied by both FasL-induced proteolytic cell death and a protein-kinase-C-associated apoptotic process in MCF-7 cells.
FasL, a 40-kDa type II transmembrane protein, has previously been shown to mediate apoptosis by binding to its cognate receptor Fas (APO-1/CD95) [14], and FAST is activated during Fas-mediated apoptosis [22]. In this study, FasL protein levels were significantly increased and the mRNA expression level of FAST was upregulated upon EA treatment of MCF-7 cells (fig. 4, table 1). It is generally believed that there are two distinct primary apoptotic signaling pathways in mammalian systems: one mediated by death receptors controlled by caspases-8 and -10, and the other by release of cytochrome c and activa-tion of a capase-9/Apaf1/cytochrome c apoptosome [19]. We found that EA fraction-induced MCF-7 cell apoptosis occurred concomitantly with activation of caspase-8 (16 h after treatment), significant cytosolic cytochrome c accu-mulation (16–24 h after treatment), and the proteolytic digestion of caspase-9 (32 h after treatment; fig. 5). In this study, pro-caspase-7 was proteolytically cleaved in MCF-7 cells (24–32 h after EA treatment) after significant accu-mulation of cytochrome c in the cytosol (fig. 5). It has been reported that cytochrome c-triggered caspase activa-tion can result in rapid cleavage of PARP [6]. In this study, the results of cDNA microarray, Northern blot, and Western blot analyses together demonstrated that in response to EA treatment, PARP was cleaved, and this cleavage, which was regulated at both the transcriptional and post-translational levels in MCF-7 cells, may have been due to the release of cytochrome c into the cytosol and activation of caspases. We, therefore, propose herein a putative apoptotic pathway (mechanism I) via FasL expression and activation of apical and executive cas-pases in MCF-7 cells upon treatment with the EA fraction of A. formosanus.
On the basis of the results obtained from functional genomic studies, we propose that another distinct apop-totic signaling pathway may be active in EA-treated MCF-7 cells. Mechanism II involves several proteins, including phospholipase C, protein kinase C, the type 1 inositol 1,4,5-triphosphate receptor (IP3R-1), c-Jun N-terminal kinase (JNK), and c-Jun, which have been reported to play important roles in apoptosis [2, 6, 20]. We show here-in that the expression of their encoded genes was sequen-tially upregulated 9–24 h after MCF-7 cells were treated with the EA fraction. Moreover, expression of the cyto-skeleton-related gene, cdc42 [3], and actin-related protein 2 (ARPC2) [16] was suppressed during the early stage of
EA treatment (3–6 h after treatment). These observations may be correlated to alterations in shape and culture of EA-treated MCF-7 cells. Further biochemical and cellu-lar biological experiments are necessary to confirm oper-ation of the mechanism II signaling pathway in these test cells.
In conclusion, we demonstrate in this study that the EA fraction of A. formosanus extract triggers apoptosis in MCF-7 cells. We also tested the anti-MCF-7 cell prolifera-tion effect of six major compounds isolated from the EA fraction, namely ·-amyrin trans-p-hydroxy cinamate (2), isorhamnetin (4), kinsenone (5), caffeic acid, quercetin, and rutin. However, none of these constituent com-pounds, alone or in combination, showed significant anti-MCF-7 cell proliferation as compared to the EA fraction. Obviously, we cannot rule out that untested minor com-ponents might account for the observed effect. Alterna-tively, the detected bioactivity of the EA fraction against MCF-7 cells might be combinational or synergistic, mani-fested by a group of cross-interacting phytocompounds present in the EA fraction, resulting in a relatively com-plex mode of action. This hypothesis is based on repeated observations and experiences of many researchers work-ing in phytochemistry and herbal medicine. They have observed many cases of an isolated single major com-pound from a herbal extract which does not possess bioac-tivity similar to that detected in partially fractionated plant extracts. Furthermore, synergistic or cross-talk ef-fects are now quite familiar to molecular biologists, as revealed by the large volume of findings from DNA microarray gene expression profiling and proteomic stud-ies. We recently reported a case relevant to the present study in which a medicinal herb extract and a single-com-pound drug conferred similar complex pharmacogenomic activities in MCF-7 cells [28]. Therefore, many re-searchers acknowledge that the bioactivity of a herbal extract may not be solely dependent on the action of a single compound. We show in this study that various bioactivities detected from the partially purified (EA) fraction of the medicinal plant A. formosanus can in fact be highly specific as demonstrated by a series of biochem-ical and microarray analyses. The consistency and repro-ducibility of the anti-MCF-7 cell bioactivity of the EA fraction were repeatedly confirmed by qualitative and quantitative determinations of the metabolite profile and index compounds in the EA fraction. Therefore we sug-gest that our results provide useful scientific evidence to support that A. formosanus extract, a folk medicine his-torically used for cancer treatment, exhibits antitumor cell activities. Previously, we have also reported that the
EA fraction of A. formosanus exhibits significant antioxi-dant activity [24]. Taken together, this study suggests that the EA fraction of the A. formosanus extract merits fur-ther investigation as a potential cancer chemopreventive agent.
Acknowledgments
This study was supported in part by grant NSC 91-2317-B-001-008 from the National Science and Technology Program for Agricul-tural Biotechnology and an institutional grant from Academia Sinica, Taipei, Taiwan, ROC.
References
1 Ali M, Heaton A, Lench D. Triterpene esters from Australian acacia. J Nat Prod 60:1150– 1156;1997.
2 Brodie C, Blumberg PM. Regulation of cell apoptosis by protein kinase C delta. Apoptosis 8:19–27;2003.
3 Brunner D. How to grab a microtubule on the move. Dev Cell 3:2–4;2002.
4 Celis JE, Kruhøffer M, Gromova I, Frederik-sen C, Østergaard M, Thykjaer T, et al. Gene expression profiling: monitoring transcription and translation products using DNA microar-rays and proteomics. FEBS Lett 481:2–16; 2000.
5Chen M, Wang J. Initiator caspases in apopto-sis signaling pathways. Apoptoapopto-sis 7:313–319; 2002.
6 Chinnaiyan AM, Dixit VM. The cell-death ma-chine. Curr Biol 6:555–562;1996.
7 Cuvillier O, Nava VE, Murthy SK, Edsall LC, Levade T, Milstien S, Spiegel S. Sphingosine generation, cytochrome c release, and activa-tion of caspase-7 in doxorubicin-induced apop-tosis of MCF7 breast adenocarcinoma cells. Cell Death Differ 8:162–171;2001.
8 Duggan DJ, Bittner M, Chen Y, Meltzer P, Trent JM. Expression profiling using cDNA microarrays. Nat Genet 21:10–14;1999. 9 Fang G, Chang BS, Kim CN, Perkins C,
Thompson CB, Bhalla KN. ‘Loop’ domain is necessary for taxol-induced mobility shift and phosphorylation of Bcl-2 as well as for inhib-iting taxol-induced cytosolic accumulation of cytochrome c and apoptosis. Cancer Res 58: 3202–3208;1998.
10 Jaiswal AS, Bloom LB, Narayan S. Long-patch base excision repair of apurinic/apyrimidinic site DNA is decreased in mouse embryonic fibroblast cell lines treated with plumbagin: involvement of cyclin-dependent kinase inhibi-tor p21Waf-1/Cip-1. Oncogene 21:5912–5922; 2002.
11 Lau WK, Chiu SK, Ma JT, Tzeng CM. Linear amplification of catalyzed reporter deposition technology on nylon membrane microarray. Biotechniques 33:564–570;2002.
12 Lin HL, Liu TS, Huang TC, Koyama T, DeVol CE. Flora of Taiwan. Taipei, Epoch Publishing, 1978, vol 5, pp 874–875.
13 Liu ZG, Hsu H, Goeddel DV, Karin M. Dissec-tion of TNF receptor 1 effector funcDissec-tions: INK activation is not linked to apoptosis while NF-ÎB activation prevents cell death. Cell 87:565– 576;1996.
14 Nagata S. Apoptosis by death factor. Cell 88: 355–365;1997.
15Naujokat C, Sezer O, Possinger K. Tumor ne-crosis factor-· and interferon-Á induce expres-sion of functional Fas ligand on HT29 and MCF7 adenocarcinoma cells. Biochem Bio-phys Res Commun 264:813–819;1999. 16 Robinson RC, Turbedsky K, Kaiser DA,
Marchand JB, Higgs HN, Choe S, et al. Crystal structure of Arp2/3 complex. Science 294: 1679–1684;2001.
17 Salvesen GS, Dixit VM. Caspase activation: the induced-proximity model. Proc Natl Acad Sci USA 96:10964–10967;1999.
18 Seufferlein T, Rozengurt E. Sphingosylphos-phorylcholine activation of mitogen-activated protein kinase in Swiss 3T3 cells requires pro-tein kinase C and a pertussis toxin-sensitive G protein. J Biol Chem 270:24334–24342;1995. 19 Tang D, Lahti JM, Kidd VJ. Caspase-8
activa-tion and Bid cleavage contribute to MCF7 cel-lular execution in a caspase-3-dependent man-ner during staurosporine-mediated apoptosis. J Biol Chem 275:9303–9307;2000.
20 Thompson CB. Apoptosis in the pathogenesis and treatment of disease. Science 267:1456– 1462;1995.
21 Thomposon HJ, Jiang C, Lu J, Mehta RG, Piazza GA, Paranka NS. Sulfone metabolite of sulindac inhibits mammary carcinogenesis. Cancer Res 57:267–271;1997.
22 Tian Q, Taupin JL, Elledge S, Robertson M, Anderson P. Fas-activated serine/threonine ki-nase (FAST) phosphorylates TIA-1 during Fas-mediated apoptosis. J Exp Med 182:865–874; 1995.
23 Verma, SP, Goldin BR. Copper modulates ac-tivities of genistein, nitric oxide, and curcumin in breast tumor cells. Biochem Biophy Res Commun 310:104–108;2003.
24 Wang SY, Kuo YH, Chang HN, Kang PL, Tsay HS, Lin KF, et al. Profiling and characteriza-tion antioxidant activities in Anoectochilus
for-mosanus Hayata. J Agric Food Chem 5:1859–
1865;2002.
25Wang TH, Wang HS, Soong YK. Paclitaxel-induced cell death – where the cell cycle and apoptosis come together. Cancer 88:2619– 2628;2000.
26 Woo JH, Lim YH, Choi YJ, Kim DG, Lee KS, Bae JH, Min DS, Chang JS, Jeong YJ, Lee YH, Park JW, Kwon TK. Molecular mechanisms of curcumin-induced cytotoxicity: induction of apoptosis through generation of reactive oxy-gen species, down-regulation of Bcl-XL and IAP, the release of cytochrome c and inhibition of Akt. Carcinogenesis 24:1199–1208;2003. 27 Yang J, Liu X, Bhalla K, Naekyung K, Ibrado
AM, Cai J. Prevention of apoptosis by Bcl-2: release of cytochrome c from mitochondria blocked. Science 275:1129–1132;1997. 28 Yang NS, Shyur LF, Chen CH, Wang SY,
Tzeng CM. Medicinal herb extract and a single-compound drug confer similar complex phar-macogenomic activities in MCF-7 cells. J Biomed Sci 11:418–422; 2004.
29 Yang X, Chang HY, Baltimore D. Autoproteo-lytic activation of pro-caspases by oligomeriza-tion. Mol Cell 1:319–325; 1998.