Plant Cell Physiol. 32(2): 269-272 (1991) JSPP © 1991
Characteristics of the Induction of the Accumulation of Proline
by Abscisic Acid and Isobutyric Acid in Detached Rice Leaves
Irene T. Chou, Chien Teh Chen and Ching Huei Kao
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
The effects of abscisic acid and isobutyric acid on levels of proline in detached rice leaves were compared. The lowest concentration of abscisic acid that induces accumulation of proline in detached rice leaves was much lower than that of isobutyric acid. The accumulation of porline induced by isobutyric acid was associated with acidification of the cell sap, whereas abscisic acid increased levels of proline without decreasing the pH of the cell sap. Potassium chloride en-hanced the accumulation of proline that was induced by abscisic acid, but it did not stimulate that induced by isobutyric acid. Of particular interest is the finding that detached rice leaves treated with fusicoccin showed an increase in levels of proline and a decrease in the pH of the cell sap. A synergistic increase in levels of proline was observed when fusicoccin and abscisic acid applied simultaneously. However, fusicoccin had no effect on the induction of the accumulation of pro-line by isobutyric acid. Benzyladenine inhibited the induction by abscisic acid of the accumula-tion of proline, but it did not inhibit the inducaccumula-tion by isobutyric acid. It is concluded that the mode of action of abscisic acid in inducing the accumulation of proline differs from that of isobutyric acid. Our results also indicate that factors other than acidification of the cell sap are in-volved in regulation of the accumulation of proline.
Key words: Abscisic acid — Isobutyric acid — Proline accumulation — Oryza sativa.
Recent reports have suggested that a decrease in in-tracellular pH, measured as the pH of the cell sap, is involv-ed in stress-, ABA- and IBA-inducinvolv-ed accumulation of pro-line in wheat coleoptiles and segments of barley leaves (Pesci and Beffagna 1984, 1985, Goring and Plescher 1986). Venekamp (1989) showed that organic acids are the source of drought-induced synthesis of proline in bean plants in the field. Previously, we reported that proline ac-cumulated in detached rice leaves during dark-induced se-nescence (Wang et al. 1982). More recently, we reported that secretion of protons from detached rice leaves plays a regulatory role in senescence (Chen et al. 1990). Secretion of protons decreases during senescence of detached rice leaves. The present investigation was conducted to com-pare the effects of ABA and IBA on the levels of proline in detached rice leaves and to test the hypothesis that ABA-and IBA-induced changes in levels of proline might be me-diated by changes in intracellular pH, as proposed by Pesci and Beffagna (1985).
Abbreviations: BA, benzyladenine; FC, fusicoccin; IBA, isobutyric acid.
Materials and Methods
Rice (Oryza sativa cv. Taichung Native 1) seedlings were grown as described previously (Kao 1980). Briefly, seedlings were grown on a stainless-steel net floated on half-strength Johnson's modified nutrient solution (John-son et al. 1975) in a 500-ml beaker. The nutrient solution (pH 4.5) was replaced every three days. Rice seedlings were grown in a greenhouse with natural daylight at 30°C by day and 25°C by night, with 95% humidity.
Apical 3-cm segments, excised from the third leaves of 12-day-old seedlings, were used. A group of 20 segments were floated in a Petri dish that contained 20 ml of basal medium (1 nui potassium phosphate, pH 5.5) or test solu-tions. All test solutions were prepared by addition of chemicals of interest to 1 mM potassium phosphate buffer (pH 5.5). Incubations were carried out at 27°C in dark-ness.
Proline was extracted and its concentration determin-ed by the method of Bates et al. (1973). Leaf segments were homogenized with 3% sulfosalicylic acid and the ho-mogenate was centrifuged. The supernatant was treated
270 I. T. Chou, C. T. Chen and C. H. Kao
with acetic acid and acid ninhydrin, boiled for 1 h, and then absorbance at 520 nm was determined. Levels of pro-line are expressed as ^umol/g fresh weight.
The method of Pesci and Beffagna (1985) was used to measure the pH of the cell sap. Leaf segments were trans-ferred at the end of treatment directly into a syringe and were frozen in liquid nitrogen. After thawing, the cell sap was pressed out from the syringe and its pH was measured.
Results
Figure 1 shows the changes in levels of proline in de-tached rice leaves incubated in various concentrations of ABA or IBA in darkness for 8 h. Levels of poline increas-ed with the increase in the concentrations of ABA or IBA. The level of proline was first found to increase in 10 JUM ABA, but only in 1 mil IBA. The lowest concentration of ABA to cause increased accumulation of proline was 100 times lower than that of IBA. Figure 1 shows the changes in the pH of the cell sap of detached leaves in-cubated in various concentrations of ABA and IBA in the dark for 8 h. The pH of the cell sap decreased with in-creases in the concentration of IBA. The pH of the cell sap also decreased during incubation in 1 mM IBA.
How-1 0 ' 1 O6 10a 10" 10" Concent rationCM}
1 0-2
Fig. 1 Effects of various concentrations of ABA and IBA on levels of proline and pH of the cell sap. Detached rice leaves were incubated in darkness for 8 h. Bars represent SE; 3 replicates.
Table 1 Effects of KC1 on ABA- and IBA-induced
ac-cumulation of proline
Treatment (//mol/g fr wt)Proline level
None 0.25 ±0.01 ABA, 10//M 0.48±0.04 ABA, 10/m + KC1, 50 mM 0.60±0.04 IBA, 10 mM 1.01 ±0.09 IBA, 10mM + KC1, 50 mM 1.06±0.04 Detached rice leaves were incubated in the dark for 8 h in either ABA or IBA with or without KC1 (mean ± SE; 4 replicates).
ever, the pH was not significantly affected by addition of ABA at the concentration from 0.1 //M to 100 fXM. Clearly, the accumulation of proline induced by IBA, and not that induced by ABA, was associated with acidification of the cell sap. Since leaf segments treated with 100//M ABA showed slight signs of wilting, 10 ^M ABA was adopted for subsequent experiments.
The increase in levels of proline induced by ABA in de-tached barley leaves was stimulated by potassium chloride (Pesci 1987, 1988, Pesci and Beffagna 1986). The effects of potassium chloride on ABA- and IBA-induced accumula-tion of proline in detached rice leaves were, therefore, com-pared. Table 1 shows that potassium chloride at 50 mM stimulated the ABA-induced accumulation of proline, but it did not stimulate the IBA-induced accumulation of pro-line. These results are in agreement with those observed in
1.8
-si
J0.6-n
T-T
T
Control FC ABA ABA
+
FC
IBA IBA
+ FC
Fig. 2 Effects of fusicoccin (FC) on levels of proline and of FC
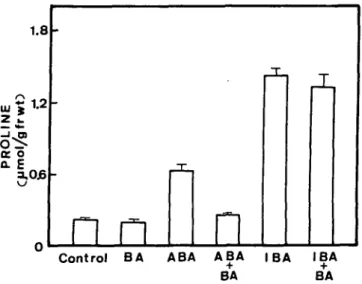
on ABA- and IBA-induced accumulation of proline. Detach-ed rice leaves were incubatDetach-ed in the dark for 8 h in ABA (10//M), IBA (10 mM), F C ( I O ^ M ) , FC(10/m) + ABA (10//M), FC (10/iM) + IBA (10 mM). Bars represent SE; 3 replicates.
Accumulation of proline in rice leaves 271 Table 2 Effects of ABA or IBA with or without
fusicoc-cin (FC) and of FC alone on the pH of the cell sap
Treatment pH of the cell sap
Control FC ABA FC + ABA IBA FC + IBA
Table 3 Effects of ABA or IBA with or without
ben-zyladenine (BA) and of BA alone on the pH of the cell sap
Treatment pH of the cell sap
6.20±0.01 6.07 ±0.01 6.20±0.02 6.08 ±0.02 6.02+0.01 6.05 ±0.02 Control BA ABA BA + ABA IBA BA + IBA 6.32 ±0.02 6.31 ±0.02 6.36±0.04 6.28 ±0.02 6.14±0.03 6.17±0.04
Detached rice leaves were incubated in the dark for 8 h in ABA (10/iM), IBA (10 HIM), FC (10/«rf), FC (10//M) + ABA (10 mM), or FC (10/iM) + IBA (10 mM) (mean ± SE; 3 replicates).
Detached rice leaves were incubated in the dark for 8 h in ABA (10//M), IBA (10 mM), BA (10/*M), BA (10/iM) + ABA (10/iM), or
BA (10J/M) + IBA (10 mM) (mean ± SE; 3 replicates).
the barley leaf system (Pesci 1987, 1988, Pesci and Beffagna 1986). However, the enhancement by potassium chloride on the ABA-induced accumulation of proline in detached rice leaves was much lower than that in the barley system. The phytotoxin fusicoccin (FC) has been shown to stimulate active export of protons (Marre 1979). To our surprise, FC at 10/iM caused not only an increase in levels of proline level but also a decrease in the pH of the cell sap in our system (Fig. 2 and Table 2). Similar results were ob-served when the concentration of FC was only 1 fiM (data not shown). Figure 2 and Table 2 show the effects of IBA and ABA in the absence and presence of FC on levels of proline and on the pH of the cell sap, respectively. IBA-in-duced accumulation of proline was clearly associated with a decrease in the intracellular pH. However, such was not
1.8 -K O o. E a,OjB
-n
TControl BA ABA ABA BA
IBA IBA BA
Fig. 3 Effects of benzyladenine (BA) on levels of proline and of BA on ABA- and IBA-induced accumulation of proline. De-tached rice leaves were incubated in the dark for 8 h in ABA (10//M), IBA (10 mM), BA (10//M), BA (10//M) + ABA (10//M), or BA (10//M) + IBA (10 mM). Bars represent SE; 3 replicates.
the case in ABA-treated segments of rice leaves. FC stimu-lated the ABA-induced accumulation of proline. How-ever, FC had no effect on the IBA-induced accumulation of proline. A synergistic effect on the increase in levels of pro-line was observed when FC and ABA were applied togeth-er. No difference was observed between the pH of cell sap of detached rice leaves treated with FC alone and the pH of detached leaves treated with FC and ABA together.
Kinetin has been shown to reduce the extent of the ac-cumulation of proline induced by ABA (Eder and Huber 1977). More recently, it was reported that benzyladenine (BA) inhibited the accumulation of proline in ABA-treated segments of barley leaves (Stewart et al. 1986). We found that BA did inhibit the ABA-induced accumulation of pro-line in detached rice leaves, but it did not inhibit the IBA-in-duced accumulation of proline (Fig. 3). No difference was observed between the pH of the cell sap of detached rice leaves treated with BA alone and the pH of those treated with BA and ABA together (Table 3).
Discussion
The mechanism of the ABA-induced accumulation of proline in detached rice leaves differs from that of the IBA-induced accumulation of proline. This conclusion is based on the observations that (a) the lowest concentration of ABA that induces the accumulation of proline was 100 times lower than that of IBA; (b) potassium chloride stimulated the accumulation of proline that was induced by ABA, but it did not stimulate the IBA-induced accumula-tion of proline; (c) a synergistic increase in levels of proline was found when FC and ABA were applied together, but FC had not effect on the IBA-induced accumulation of pro-line; (d) BA did inhibit the accumulation of proline by ABA, but it did not inhibit that induced by IBA; (e) the ac-cumulation of proline induced by IBA was associated with acidification of the cell sap, whereas ABA increased levels of proline without any decrease in the pH of the cell sap.
272 I. T. Chou, C. T. Chen and C. H. Kao It is apparent that the effect of ABA is a "hormonal" effect
(ABA is effective at very low concentrations; it acts synergistically with FC and BA; the increase in levels of of proline is relatively independent of the pH of the cell sap), whereas the effect of IBA can be explained by the. decrease of the pH of the cell sap.
Of particulr interest is the finding that FC increased levels of proline in detached rice leaves. Although FC is known to promote secretion of protons from detached rice leaves (Chen and Kao 1988), FC was observed to decrease the pH of the cell sap. The decrease in the pH of the cell sap after application of FC has been reported by other in-vestigators (Bertel and Felle 1985, Brummer et al. 1985, Hager and Moser 1985, Roberts et al. 1981). FC is a diacetate ester and, according to Hager and Moser (1985), it can be hydrolyzed like naphthylacetate and other car-boxyester compounds. Such hydrolysis might explain why FC decreases the pH of the cell sap. Apparently, the ac-cumulation of proline induced by FC is associated with a decrease in the pH of the cell sap.
Pesci and Beffagna (1985) reported that FC completely suppressed the ABA-induced increase in levels of proline during a 7-h treatment period. In contrast to their results, our data indicate that FC promotes the ABA-induced ac-cumulation of proline in detached rice leaves. ABA- and IBA-induced accumulation of proline has been proposed to be mediated by acidification of the cell sap (Pesci and Beffagna 1985). However, our results demonstrate that the accumulation of proline that is induced by IBA, but not by ABA, is associated with acidification of the cell sap. It ap-pears that, in detached rice leaves, factors apart from acidification of the cell sap may also be involved in regulating the accumulation of proline.
At present, it remains unclear whether acidification of the cell sap regulates the accumulation of proline through stimulation of the synthesis of proline, reduction in the utilization of proline, or both. Further studies should pro-vide valuable information on the mechanism of accumula-tion of proline.
This research was supported by the National Science Council of the Republic of China (NSC 77-04O9-B002-11).
References
Bates, L. S. (1973) Rapid determination of free proline for water stress studies. Plant Soil 39: 205-207.
Bertel, A. and Felle, H. (1985) Cytoplasmic pH of root hair cells of Sinapis alba recorded by a pH-sensitive microelectrode. Does fusicoccin stimulate the proton pump by cytoplasmic acidification? J. Exp. Bot. 36: 1142-1149.
Brummer, B., Bertl, A., Potrykus, I., Felle, H. and Parish, R. W.
(1985) Evidence that fusicoccin and indole-3-acetic acid induce cytosolic acidification of Zea mays cells. Fed. Eur. Biochem. Soc. 189: 109-114.
Chen, C. T. and Kao, C. H. (1988) Proton secretion in rice leaves. Bot. Bull. Academia Sinica 29: 315-320.
Chen, C. T., Chou, I. T. and Kao, C. H. (1990) Senescence of rice leaves XX. Changes of proton secretion during senes-cence. Plant Sci. 66: 29-34.
Eder, A. and Huber, W. (1977) About the effects of abscisic acid and kinetin on biochemical changes in Pennisetum typhoides during stress conditions. Z. Pflanzenphysiol. 84: 301-311. Goring, H. and Plescher, F. (1986) Proline accumulation
in-duced by weak acids and IAA in coleoptiles of wheat seedlings. Biol. Plant. 28: 401-406.
Hager, A. and Moser, I. (1985) Acetic acid esters and permeable weak acids induce active proton extrusion and extension growth of coleoptile segments by lowering the cytoplasmic pH. Planta 163: 391-400.
Johnson, C. M., Stout, P. R., Broyer, T. C. and Carlton, A. B. (1957) Comparative chlorine requirements of different plant species. Plant Soil 8: 337-353.
Kao.C. H. (1980) Sensecenceof rice leaves IV. Influence of ben-zyladenine on chlorophyll degradation. Plant Cell Physiol. 21: 339-344.
Marre, E. (1979) Fusicoccin: a tool in plant physiology. Annu. Rev. Plant Physiol. 30: 273-288.
Pesci, P. (1987) ABA-induced proline accumulation in barley leaf segments: dependence on protein synthesis. Physiol. Plant. 71: 287-291.
Pesci, P. (1988) Ion fluxes and abscisic acid-induced proline ac-cumulation in barley leaf segments. Plant Physiol. 86: 927-930.
Pesci, P. and Beffagna, N. (1984) Inhibiting effect of fusicoccin on abscisic acid-induced proline accumulation in barley leaves. Plant Sci. Lett. 36: 7-12.
Pesci, P. and Beffagna, N. (1985) Effect of weak acids on proline accumulation in barley leaves: a comparison between abscisic acid and isobutyric acid. Plant Cell Environ. 8: 129-133. Pesci, P. and Beffagna, N. (1986) Influence of exogenously
sup-plied potassium and sodium salts on the abscisic acid-induced proline accumulation in barley leaf segments. Physiol. Plant. 67: 123-128.
Roberts, J. K. M., Ray, P. M , Wade-Jardetzky, N. and Jardet-zky, O. (1981) Extent of intracellular pH changes during H ex-trusion by maize root-tip cells. Planta 152: 76-78.
Stewart, C. R., Voetberg, G. and Rayapati, P. J. (1986) The effect of benzyladenine, cycloheximide, and cordycepin on wilting-induced abscisic acid and proline accumulations and abscisic acid- and salt-induced proline accumulation in barley leaves. Plant Physiol. 82: 703-707.
Venekamp, J. H. (1989) Regulation of cytosol acidity in plants under conditions of drought. Physiol. Plant. 76: 112-117. Wang, C. Y., Cheng, S. H. and Kao, C. H. (1982) Senescence of
leaves VII. Proline accumulation in senescing excised leaves. Plant Physiol. 69: 1348-1349.