人類血液Haptoglobin之抗氧化角色
156
0
0
全文
(2) 人類血液 Haptoglobin 之抗氧化角色 Antioxidant Role of Human Plasma Haptoglobin. 研 究 生:曾繼鋒. Student:Chi Feng Tseng. 指導教授:毛仁淡. Advisor:Simon JT Mao. 國 立 交 通 大 學 生物科技學系 博 士 論 文. A Thesis Submitted to Department of Biological Science and Technology College of Biological Science and Technology National Chiao Tung University in Partial Fulfillment of the Requirements for the Degree of PhD in Biological Science and Technology April 2006 Hsinchu, Taiwan, Republic of China. 中華民國九十五年四月.
(3) 博碩士論文授權書 本授權書所授權之論文為本人在_交通__大學(學院)_生物科技學系_系所 _____組__94__學年度第_二_學期取得_博_士學位之論文。 論文名稱:_人類血液Haptoglobin之抗氧化角色__ ___________ 指導教授:_毛仁淡__________________________ 1.■同意 □不同意 本人具有著作財產權之上列論文全文(含摘要)資料,授予行政院國家科學委員會科學技 術資料中心(或改制後之機構),得不限地域、時間與次數以微縮、光碟或數位化等各種 方式重製後散布發行或上載網路。 本論文為本人向經濟部智慧財產局申請專利(未申請者本條款請不予理會)的附件之一, 申請文號為:______________,註明文號者請將全文資料延後半年再公開。 2.■同意 □不同意 本人具有著作財產權之上列論文全文(含摘要)資料,授予教育部指定送繳之圖書館及國 立交通大學圖書館,基於推動讀者間「資源共享、互惠合作」之理念,與回饋社會及學 術研究之目的,教育部指定送繳之圖書館及國立交通大學圖書館得以紙本收錄、重製與 利用;於著作權法合理使用範圍內,不限地域與時間,讀者得進行閱覽或列印。 本論文為本人向經濟部智慧財產局申請專利(未申請者本條款請不予理會)的附件之一, 申請文號為:______________,註明文號者請將全文資料延後半年再公開。 3.■同意 □不同意 本人具有著作財產權之上列論文全文(含摘要),授予國立交通大學與台灣聯合大學系統 圖書館,基於推動讀者間「資源共享、互惠合作」之理念,與回饋社會及學術研究之目 的,國立交通大學圖書館及台灣聯合大學系統圖書館得不限地域、時間與次數,以微縮、 光碟或其他各種數位化方式將上列論文重製,並得將數位化之上列論文及論文電子檔以 上載網路方式,於著作權法合理使用範圍內,讀者得進行線上檢索、閱覽、下載或列印。 論文全文上載網路公開之範圍及時間 – 本校及台灣聯合大學系統區域網路: 年 月 日公開 校外網際網路: 年 月 日公開 上述授權內容均無須訂立讓與及授權契約書。依本授權之發行權為非專屬性發行權利。依本授 權所為之收錄、重製、發行及學術研發利用均為無償。上述同意與不同意之欄位若未鉤選,本 人同意視同授權。 研究生簽名: 學號: 9028808 (親筆正楷) (務必填寫).
(4) 國家圖書館博碩士論文電子檔案上網授權書 本授權書所授權之論文為本人在_交通_大學(學院)_生物科技研究所 系所 _____組__94__學年度第_二_學期取得_博_士學位之論文。 論文名稱:__人類血液Haptoglobin之抗氧化角色___________ 指導教授:__毛仁淡 ■同意. _______________________. □不同意. 本人具有著作財產權之上列論文全文(含摘要),以非專屬、無償授權國家圖書館,不限 地域、時間與次數,以微縮、光碟或其他各種數位化方式將上列論文重製,並得將數 位化之上列論文及論文電子檔以上載網路方式,提供讀者基於個人非營利性質之線上 檢索、閱覽、下載或列印。 上述授權內容均無須訂立讓與及授權契約書。依本授權之發行權為非專屬性發行權 利。依本授權所為之收錄、重製、發行及學術研發利用均為無償。上述同意與不同意 之欄位若未鉤選,本人同意視同授權。 研究生簽名: (親筆正楷) 日期:民國 1.. 學號: 9028808 (務必填寫) 年. 月. 日. 本授權書請以黑筆撰寫,並列印二份,其中一份影印裝訂於附錄三之一(博碩士論 文授權書)之次頁﹔另一份於辦理離校時繳交給系所助理,由圖書館彙總寄交國家 圖書館。.
(5) 人類血液 Haptoglobin 之抗氧化角色 學生:曾繼鋒. 指導教授:毛仁淡 博士 國立交通大學. 生物科技學系. 摘. 博士班. 要. 人類血液中之 Haptoglobin (Hp),與血型分類相似,可以分為三種表 現型:1-1、2-1、與 2-2。然而在這些 Hp 表現型之結構與功能的關係,由 於其複雜的結構與困難繁複的純化步驟,目前的瞭解仍十分有限。在此我 們發展一個可以純化每一種 Hp 表現型的簡便方法。首先將血漿通過已結 合可專一辨識 Hp 之單株抗體,將所得到之 Hp 再通過 gel filtration 管柱, 藉由 SDS-PAGE 分析,所得到之 Hp 純度可大於 95%,並可保有其本身所 具有之醣基成分與血紅素結合能力。經由 Circular dichroism 分析,Hp 1-1 (29%) 之 α-helix 組成比例高於 2-1 (22%) 與 2-2 (21%)。這個方法較現有 Hp 的純化方法有了明顯的改善與進步。為了進一步瞭解 Hp 在 in vitro 之 抗氧化活性,thiobarbituric acid-reactive substances (TBARS) assay 用來估 計在脂質過氧化反應中 Hp 的抗氧化活性。Hp 在銅離子所誘發之脂質過 氧化反應中表現了極強之抗氧化能力。此外,在另一種親水性自由基產生 者 2,2’-azobis(2-amidinopropane)-dihydrochloride (AAPH) 所誘發的脂質過氧化 反應中, Hp 亦具有相似之抗氧化特性,因此推測 Hp 可能也扮演著自由 基清除者的角色。為了更進一步研究結構對於其抗氧化特性的影響, I.
(6) carboxymethylation 被用來阻絕在 Hp 中 cysteine 間雙硫鍵的形成,有趣地, 經過修飾後的 Hp 反而較 native Hp 表現出更強之抗氧化能力,因此推論 在 native 的構形中,抗氧化 domain 可能並未完全暴露在外。為了更深入 研究 Hp 在細胞內的抗氧化角色,我們將 Hp 的 cDNA 放入含有 CMV 啟 動子之 pcDNA 3.0 載體中,並轉殖至本身不會表現 Hp 之 Chinese Hamster Ovary (CHO) 細胞中,發現確實可以增加該細胞對於氧化壓力的耐受度, 在添加雙氧水的培養條件下 24 小時,其耐受度較未轉殖之細胞高出 1 倍。 因此 Hp 在 in vitro 與 ex vivo 的研究中皆表現出極佳之抗氧化能力。最後 我們分析了目前普遍使用之抗氧化活性檢測方法,並且闡釋如何研發與設 計防止動脈硬化之強效抗氧化藥物。首先抗氧化藥物必須可以專一地 LDL 結合。第二,必須具有高度之 bioavailability。文中並針對這些抗氧化作用 機制與分析程序之原理與策略進行討論。. II.
(7) Antioxidant Role of Human Plasma Haptoglobin Student::Chi Feng Tseng. Advisor:Dr. Simon JT Mao. Department of Biological Science and Technology National Chiao Tung University. Abstract Similar to blood type, human plasma haptoglobin (Hp) is classified as 3 phenotypes: Hp 1-1, 2-1, or 2-2.. The structural and functional relationship between the Hp phenotypes. has not been studied in detail due to their complicated structures and difficult isolation procedures.. In the present study, we developed a simple protocol that can be used to. purify each Hp phenotype.. Plasma was first passed through an affinity column coupled. with a high affinity Hp monoclonal antibody. The bound Hp was eluted and further chromatographied on a HPLC. The homogeneity of purified Hp 1-1, 2-1, or 2-2 was greater than 95% as judged by SDS polyacrylamide gel electrophoresis. carbohydrate moiety and hemoglobin-binding ability.. It retained the. Circular dichroic spectra showed. that the α-helical content of Hp 1-1 (29%) was higher than that of Hp 2-1 (22%) and 2-2 (21%). The procedures described here represent a significant improvement in current purification methods for each Hp phenotypes. To investigate in vitro antioxidant role of Hp, thiobarbituric acid-reactive substances (TBARS) was used to estimate antioxidant activity of Hp in low-density lipoprotein (LDL) lipid peroxidation.. We demonstrated that. Hp molecule was an extremely potent antioxidant activity in Cu2+-induced LDL peroxidation.. Using. 2,2’-azobis(2-amidinopropane)-dihydrochloride. III. (AAPH),. a.
(8) hydrophilic decomposed radical initiator, it produced a similar antioxidant effect of Hp against LDL oxidation suggesting a free radical-scavenging role of Hp.. To study the. structural effect in its antioxidant activity, carboxymethylation that alters the overall structure of Hp by blocking the formation of disulfide linkages between cysteine residues was used for the evaluation.. Interestingly, carboxymethylated Hp exerting higher. antioxidant potency than that of native Hp indicated that the antioxidant domain of Hp might not be fully exposed. To investigate antioxidant role of Hp on the cellular level, the cDNA of Hp 1-1 was cloned, constructed (containing the pcDNA3.0 vector with CMV promoter) and transfected to Chinese Hamster Ovary (CHO) cells expressing no Hp. These transfected CHO cells were able to express Hp 1-1 and significantly (P<0.001) elevated the tolerance against the oxidative stress. The elevation was about twice-higher than that normal CHO cells when challenged with hydrogen peroxide for 24 h. Thus, Hp plays a provocative antioxidant role as demonstrated in our in vitro and ex vivo studies. Finally, we analyzed commonly used analytical methods for measuring the antioxidant potency and outlined the critical steps as how to evaluate and design a potent antioxidant agent that can be used for the intervention of atherosclerosis.. We conclude that an. antioxidant should be first targeted and incorporated into human LDL.. Second, the. candidate compound should possess high bioavailability. The rationale and strategy for the analytical procedures are discussed.. IV.
(9) 誌. 謝. 在經過五年辛苦帶著甘甜、微笑伴著眼淚的博士求學過程中,特別 要感謝指導教授毛仁淡老師,無論在生活上、實驗上都給予繼鋒悉心地指 導與幫助,讓繼鋒得以成長,博士論文得以完成,在此獻上最誠摯地感謝。 同時也謝謝張正老師,趙雲鵬老師,曾慶平老師,林志生老師,戴鏗碩老 師耐心地批閱斧正,使論文得以更臻完整詳實。. 這些日子以來,實驗室是我第二個家,學長以及學弟妹就像是家人 一般,陪我度過這酸甜苦辣、多樣地五年光陰。阿木學長、文亮、學弟妹 以祥、明誌、佳靜、心怡、怡荏、繡文、媛婷、忠義、彰威、盛龍、惠君、 宏輝、俊瑩、韶嬋、珮真、文昭、立品、佩茹、中曦、冠佑、靜怡,謝謝 大家給了我這麼多充實愉快地回憶,未來的日子裡,一起再從笑傲回味這 一段充滿著喜怒哀樂的記憶吧。也謝謝靖婷、盈蓉、國領、千婷、政男, 很難得地,我們共同從碩士班,博士班一路奮鬥至此,未來也要記住這份 情誼,大家再一起打拼。. 最後我要感謝我的家人及女朋友英潔,謝謝你們在背後默默地支持 著我,讓我可以不斷地鼓起勇氣,去克服生活中接踵而來的挑戰,謹以此 論文獻給你們,這是我們共同的榮耀。. V.
(10) 目. 錄. 中文摘要 --------------------------------------------------------------------------------------------------I Abstract --------------------------------------------------------------------------------------------------III 誌謝 ------------------------------------------------------------------------------------------------------ V 目錄 ----------------------------------------------------------------------------------------------------- VI Overview-------------------------------------------------------------------------------------------------- 1 References ------------------------------------------------------------------------------------------------ 8 Section 1: A novel approach for Hp purification Abstract --------------------------------------------------------------------------------------------------14 Introduction----------------------------------------------------------------------------------------------15 Materials and methods ---------------------------------------------------------------------------------17 Results----------------------------------------------------------------------------------------------------22 Discussion -----------------------------------------------------------------------------------------------24 References -----------------------------------------------------------------------------------------------28 Figure Legends------------------------------------------------------------------------------------------31 Figures ---------------------------------------------------------------------------------------------------34 Tables-----------------------------------------------------------------------------------------------------43 Section 2: Study of antioxidant role of Hp in LDL oxidation and in transfected cells Abstract --------------------------------------------------------------------------------------------------47 Introduction----------------------------------------------------------------------------------------------48 Materials and methods ---------------------------------------------------------------------------------50 Results----------------------------------------------------------------------------------------------------55 Discussion -----------------------------------------------------------------------------------------------58 References -----------------------------------------------------------------------------------------------62 Figure Legends------------------------------------------------------------------------------------------66 Figures ---------------------------------------------------------------------------------------------------69 Tables-----------------------------------------------------------------------------------------------------78 Section 3: Analysis of antioxidant as a therapeutic agent for atherosclerosis Abstract --------------------------------------------------------------------------------------------------80 Introduction----------------------------------------------------------------------------------------------81 Recent antioxidant therapies for atherosclerosis----------------------------------------------------84 Potential antioxidants as antitherosclerotic agents -------------------------------------------------86 Commonly used analytical methods of antioxidant potency --------------------------------------98 Rational design of a synthetic antioxidant as an antiatherosclerotic agent -------------------- 106 VI.
(11) Conclusion--------------------------------------------------------------------------------------------- 115 References --------------------------------------------------------------------------------------------- 114 Figure Legends---------------------------------------------------------------------------------------- 135 Figures ------------------------------------------------------------------------------------------------- 139. VII.
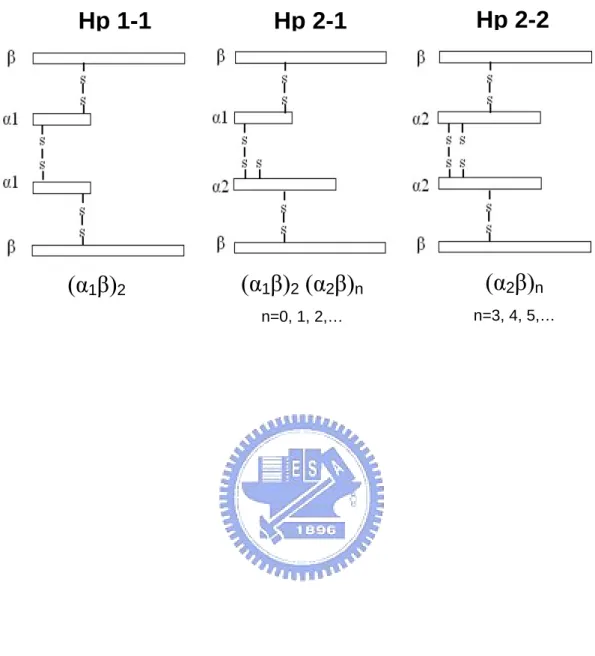
(12) Overview Coronary artery and other vascular diseases (including myocardial infarction, MI) are the leading cause of death in the Western industrialized countries [1-3].. The mortality of the. diseases are the top third-fourth in Taiwan, and has since become the leading killer, despite the overall improvement in mortality of the patients treated with hypocholesterolemic drugs. One concept that has received much recent attention for the pathogenesis of atherosclerosis is the LDL-modification hypothesis [4-6], which postulates that atherosclerotic plaque resulted from the uptake of oxidized LDL by macrophages in the arterial wall followed by smooth muscle cell (SMC) migration and proliferation [7]. In this respect, an understanding of the biochemical events, which impact on the oxidation stress and SMC proliferation into the intima deserves to receive much attention.. Inflammatory phenomena at sites of. atherosclerotic plaques are increasingly thought to be major determinants of the progression and clinical outcome of atherosclerosis disease [7-10].. Therefore, attention is being paid to. systemic markers that may reflect the inflammatory activity in the plaques. Recently we found that haptoglobin (Hp), an acute phase protein elevated sharply during the infection and inflammation, was substantially accumulated in atherosclerotic lesions [].. Nevertheless,. while the pathogenesis of atherosclerosis has been continuously explored, the Hp molecule that may protect against the progression of atherosclerosis has never been reported [11-15]. Hp is classified as three phenotypes, Hp 1-1, 2-1, and 2-2.. The primary translation. product of Hp mRNA is a polypeptide that dimerizes cotranslationally and is proteolytically cleaved while still in the endoplasmic reticulum [16,17].. All the phenotypes share the same. 2 β chains (each with about Mr 40,000 dalton containing 245 amino acids and approximately 30% carbohydrate) [18-20].. A typical structure of homozygous Hp 1-1 is composed of two. identical α 1 chains (each with about Mr 9,000 dalton containing 83 amino acids). Homozygous Hp 2-2 is composed of two identical α 2 chains (each with about Mr 16,500 1.
(13) dalton containing 142 amino acids) as compared to that of heterozygous Hp 2-1 containing one each of α 2 and α 1 [18].. Polymeric form of Hp 2-1 or 2-2 is thought to be associated. with the complications of myocardial infarction [21-22], kidney failure [23-26], and diabetics [21,27,28].. Hp is also a hemoglobin-binding protein ubiquitously present in the plasma of. all vertebrates and believed to participate in hemoglobin transport [29].. The antioxidant. activity of Hp was thought to be related from its binding ability with hemoglobin [15]. Hp may reduce loss of hemoglobin (Hb) and iron through the formation of an Hb-Hp complex which is not filtered through the glomeruli but transported to the liver [30].. The. Hb-Hp complex is cleared from circulation in the liver by recognition of a specific hepatic macrophage receptor CD163 [19,31].. Thus, the in vivo function of haptoglobin was. established as the plasma protein responsible for capture and clearance of excellular Hb from circulation, thereby preventing its toxicity to vasculature components [32].. However,. studies on Hp knockout mice demonstrated that lack of Hp does not impair clearance of Hb from plasma [33].. Indication that uptake of free hemoglobin from circulation is faster than. that of its complex with haptoglobin pointed to other, more urgent functions for this protein [33].. It appears that, by binding hemoglobin, Hp can serve as a vascular antioxidant.. Haptoglobin was shown to completely inhibit the oxidative activity of Hb toward lipids as well as LDL protein. Because oxidative modification of LDL plays a critical role in the pathogenesis of atherosclerosis, Hp can be considered as a central antiatherosclerotic agent.. 2.
(14) The present dissertation is divided as three sections:. 1.. A novel approach for Hp purification. 2.. Study of antioxidant role of Hp in LDL oxidation and in transfected cells. 3.. Analysis of antioxidant as a therapeutic agent for atherosclerosis. Section 1: A novel approach for Hp purification To evaluate the antioxidant role of Hp, in the present thesis, it is essential to evaluate and develop a convenient purification method of Hp. purification of Hp frequently suffer some drawbacks.. The methods currently used for the For example, Rademacher et al. utilize. the chicken hemoglobin-Sepharose affinity column to isolate human Hp [34]; the harsh-elution condition (8 M urea) causes the dissociation of a hemoglobin subunit from the sepharose. Meanwhile, human apolipoprotein A-I appears to be another major contaminant. Wassdal et al. use rabbit hemoglobin-Sepharose; the hemoglobin is still co-eluted from the columns [35].. Travis et al. employ Sephadex G-200 gel filtration, but the purified Hp is. accompanied with large amounts of IgM and α-2 macroglobulin [36].. Morimatsu et al.. provide a modified method using HPLC with anion-exchange, Sephacryl S-300, TSK Phenyl-5PW, and TSK DEAE-5PW columns together; the procedures however are time-consuming, and the yield is relatively low (2.5 mg per 130 ml acute phase serum) [37]. Although Katnik et al. have shown a single-step isolation for Hp using an antibody-affinity column, the phenotypes, final purity, and the biological properties of Hp are not fully reported [38].. Presumably, the purpose of their report was to use isolated Hp for raising monoclonal. antibodies. The similar antibody affinity-column procedure was employed in our laboratory, but the isolated Hp was not pure. Therefore, we established simple two-step procedures for each Hp 1-1, 2-1, and 2-2 3.
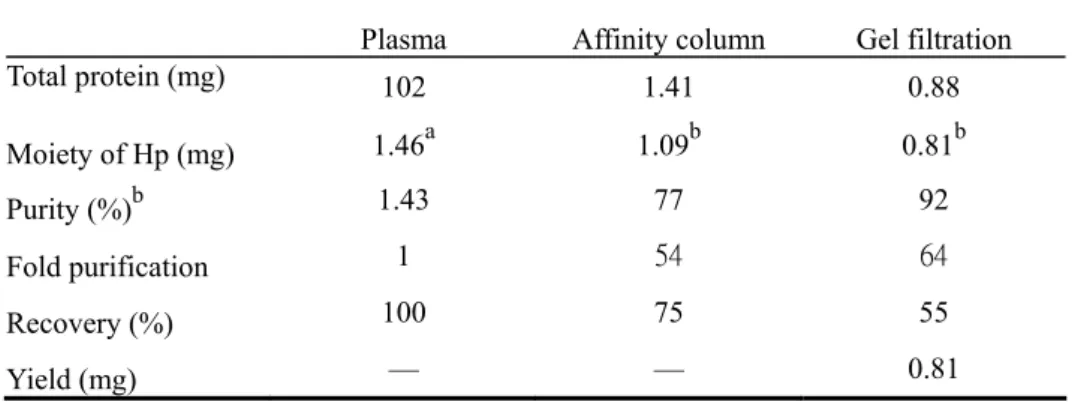
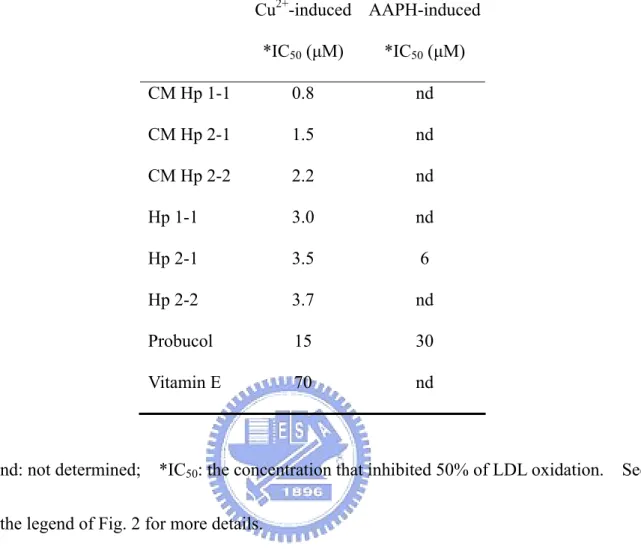
(15) purification using a monoclonal antibody affinity-column followed by a HPLC Superose 12 gel filtration.. Finally, some of the biochemical and physical properties with respect to each. Hp phenotype were characterized and discussed.Antibody-affinity chromatography following a gel filtration column was then developed and used for purification. recovery of purified Hp was at least 92 and 55%, respectively.. The purity and. Each phenotype of isolated. Hp possessed the ability to bind hemoglobin and retained its carbohydrate moiety.. The CD. spectrum of each Hp phenotype was also determined to estimate the secondary structure of Hp. For the first time, according to the best of our knowledge, we showed that the α-helical content of Hp 1-1, 2-1, and 2-2 was 29, 22, and 21%, respectively.. Therefore, Hp 1-1. possessed a more ordered structure than that of polymeric forms of Hp 2-1 and 2-2. In conclusion, due to the structural heterogeneity, it is difficult to purify Hp from human plasma, particularly with the Hp 2-1 and 2-2 polymers.. The present report provides a simple method. for the purification of Hp phenotypes with relatively high yield. Hp 1-1, 2-1, and 2-2 can therefore be prepared and used for the study of structural and functional relationship with the pathogenesis in the diseases of interest.. Section 2: Study of antioxidant role of Hp in LDL oxidation and in transfected cells We found that Hp was an extremely potent antioxidant and the activity of Hp 1-1 was differentially and moderately greater than that of Hp 2-1 and 2-2.. We also demonstrated that. Hp 1-1 cDNA transfected Chinese hamster ovary (CHO) K1 cells (normally not expressing the Hp molecules) significantly resist against oxidative stress.. The role of Hp as an. antioxidant molecule is therefore identified. The clinical significance with respect to the structure and function of Hp phenotype is discussed.. To determine the antioxidant potency. of Hp, TBARS assay using Cu2+-induced LDL oxidation was employed. Hp showed an 4.
(16) extremely potency against LDL oxidation with a dose-dependent manner: The potency was 5 x greater than that of probucol and almost 20 x than vitamin E, in which probucol is known as a highly potent antioxidant compound used in clinics.. Notably, the antioxidant activity of. monomeric Hp 1-1 was moderately higher than that of polymeric Hp 2-1 and 2-2. To study the structural effect in its antioxidant activity, while the cystein was carboxymethylated, it exerted essentially a disordered structure in circular dichroism (CD). change, however, did not attenuate its antioxidant activity. markedly increase in antioxidant activity by about 4 x.. Such conformational. Unexpectedly, it resulted in a. Taking together, the antioxidant. potency expressed as IC50 in ranking was: CM Hp1-1 > CM Hp 2-1 > CM Hp 2-2 > Hp 1-1 > Hp 2-1 > Hp 2-2 > probucol > vitamin E. Subsequently, we tested its ability for scavenging free radicals generated from a water-soluble azo-compound AAPH. The tested antioxidant activity of Hp 2-1 was shown as a dose-dependent fashion with an IC50 about 5 x greater than that of probucol. Presumably, the binding domain of Hp to Hb is dependent on the overall three-dimensional structure of Hp.. We further examined the effect of carboxymethylation of. Hp on the formation of Hp-Hb complex using a HPLC technique.. However, the binding was. totally abolished when Hp was carboxymethylated. Since the native Hp possessed its free radical-scavenging ability and the antioxidant activity of carboxymethylated Hp was superior to native Hp, it might imply that the antioxidant nature of Hp was independent on its binding ability to Hb. To evaluate the ex vivo antioxidant activity of Hp, a CHO-K1 cell line was transfected with a pcDNA3.0 vector containing CMV promoter-driven Hp 1-1 cDNA. The plasmid construct also contained a selection marker, geneticin, for conditionally expressing the Hp 1-1. RT-PCR showed that the un-transfected cells did not express endogenous Hp mRNA.. 5.
(17) Expression of Hp 1-1 protein in the culture medium and CHO cells was confirmed by Western blot and immunocytochemical staining. Hydrogen peroxide (H2O2) was then added to the CHO-K1 cell culture. Cells with and without Hp 1-1transfection were treated with variable dosages of H2O2 for 24 h.. Upon the challenge of H2O2 the relative cell survival. ratios of transfected CHO-K1 cells were statistically and significantly higher (P<0.001) than that of untransfected CHO-K1 cells, especially in the presence of 1 mM H2O2.. Thus, the. expression of Hp elevated the cell tolerance against the H2O2-induced oxidative stress. In conclusion, we demonstrated that Hp was an extremely potent antioxidant molecule and have identified its antioxidant role in cell model.. Blockage of disulfide linkages of Hp. resulted in a loss of its ability to form a complex with Hb and yet exerted almost 4 x greater antioxidant activity than that native Hp.. Thus, in addition to its role in forming Hp-Hb. complex to block the heme group, Hp may participate in an independent antioxidant role for those cells expressing Hp.. Whether or not Hp may be directly beneficial for free-radical. associated atherosclerosis and myocardial infarction will be a subject of interest and challenge.. Section 3: Analysis of antioxidant as a therapeutic agent for atherosclerosis Atherosclerosis and its complications are the major causes of mortality in industrialized countries [1-3]. Research into the oxidation of lipoprotein has yielded many insights into the process underlying the development of atherosclerosis.. Oxidative modification of low. density lipoprotein (LDL) has been suggested as an initial step in the pathogenesis of atherosclerosis [4,6].. However, up until now, investigations of antioxidants have focused on. three main dietary antioxidant vitamins (β-carotene, vitamin C, and vitamin E) [39-41] and some synthetic compounds [42-44].. Among those antioxidants described above, probucol, a 6.
(18) synthetic compound, has been shown to be an extremely potent and effective antioxidant in preventing against the formation of atherosclerosis in both in vitro and ex vivo studies [42,43]. The present review focuses on commonly used analytical methods for measuring the antioxidant potency and outlines the critical steps as how to evaluate and design a potent antioxidant agent that can be used for the intervention of atherosclerosis.. We conclude that. an antioxidant should be first targeted and incorporated into human LDL.. Second, the. candidate compound should possess high bioavailability. From the atherogenesis process and we evaluated those currently-used and potential antioxidant candidates for preventing the formation of atherosclerosis.. The critical. consideration in designing a compound that can be effectively used for antioxidant therapy in atherosclerosis are reviewed as 5 sections: 1) The oxidation hypothesis and atherogenesis induced by oxidized LDL; 2) Recent antioxidant therapies for atherosclerosis; 3) potential antioxidants as antiatherosclerotic agents; 4) commonly used analytical methods of antioxidant potency; 5) rational design of a synthetic antioxidant as an antiatherosclerotic agent.. 7.
(19) References [1]. Gurfinkel E, Lernoud V. (2006) The role of infection and immunity in atherosclerosis. Expert. Rev. Cardiovasc. Ther. 4:131-137.. [2]. Viles-Gonzalez JF, Fuster V, Badimon JJ. (2004) Atherothrombosis: a widespread disease with unpredictable and life-threatening consequences. Eur. Heart J. 25:1197-1207.. [3]. Chisolm GM, Steinberg D. (2000) The oxidative modification hypothesis of atherogenesis: an overview. Free Radic Biol Med. 28:1815-1826.. [4]. Steinberg D, Parthasarathy S, Carew TE, Khoo JC, Witztum JL. (1989) Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 320:915-924.. [5]. Witztum JL. (1994) The oxidation hypothesis of atherosclerosis. Lancet. 344:793-795.. [6]. Navab M, Ananthramaiah GM, Reddy ST, Van Lenten BJ, Ansell BJ, Fonarow GC, Vahabzadeh K, Hama S, Hough G, Kamranpour N, Berliner JA, Lusis AJ, Fogelman AM. (2004) The oxidation hypothesis of atherogenesis: the role of oxidized phospholipids and HDL. J. Lipid Res. 45:993-1007.. [7]. Noguchi N. (2002) Novel insights into the molecular mechanisms of the antiatherosclerotic properties of antioxidants: the alternatives to radical scavenging. Free Radic. Biol. Med. 33:1480-1489.. [8]. Born, G.V.R. (1993) New Horizons in Coronary Heart Disease, Current Science.. [9]. Upston JM, Kritharides L, Stocker R. (2003) The role of vitamin E in atherosclerosis. Prog. Lipid Res. 42:405-422.. [10] Steinberg D. (2006) The pathogenesis of atherosclerosis. An interpretive history of the cholesterol controversy, part IV: the 1984 coronary primary prevention trial ends it--almost. J. Lipid Res. 47:1-14. [11] Arredouani MS, Kasran A, Vanoirbeek JA, Berger FG, Baumann H, Ceuppens JL. 8.
(20) (2005) Haptoglobin dampens endotoxin-induced inflammatory effects both in vitro and in vivo. Immunology. 114:263-271. [12] Wang Y, Kinzie E, Berger FG, Lim SK, Baumann H. (2001) Haptoglobin, an inflammation-inducible plasma protein. Redox. Rep. 6:379-385. [13] Bamm VV, Tsemakhovich VA, Shaklai M, Shaklai N. (2004) Haptoglobin phenotypes differ in their ability to inhibit heme transfer from hemoglobin to LDL. Biochemistry. 43:3899-3906. [14] Tseng CF, Lin CC, Huang HY, Liu HC, Mao SJT. (2004) Antioxidant role of human haptoglobin. Proteomics. 4:2221-2228. [15] Asleh R, Guetta J, Kalet-Litman S, Miller-Lotan R, Levy AP. (2005) Haptoglobin genotype- and diabetes-dependent differences in iron-mediated oxidative stress in vitro and in vivo. Circ. Res. 96:435-441. [16] Hanley JM, Haugen TH, Heath EC. (1983) Biosynthesis and processing of rat haptoglobin. J. Biol. Chem. 258:7858-7869. [17] Wicher KB, Fries E. (2004) Prohaptoglobin is proteolytically cleaved in the endoplasmic reticulum by the complement C1r-like protein. Proc. Natl. Acad. Sci. U S A. 101:14390-14395. [18] Tseng CF, Huang HY, Yang YT, Mao SJT. (2004) Purification of human haptoglobin 1-1, 2-1, and 2-2 using monoclonal antibody affinity chromatography. Protein Expr. Purif. 33:265-273. [19] Kristiansen M, Graversen JH, Jacobsen C, Sonne O, Hoffman HJ, Law SK, Moestrup SK. (2001) Identification of the haemoglobin scavenger receptor. Nature. 409:198-201. [20] Patzelt D, Geserick G, Schroder H. (1988) The genetic haptoglobin polymorphism: relevance of paternity assessment. Electrophoresis. 9:393-397. [21] Suleiman M, Aronson D, Asleh R, Kapeliovich MR, Roguin A, Meisel SR, Shochat M, Sulieman A, Reisner SA, Markiewicz W, Hammerman H, Lotan R, Levy NS, Levy AP. (2005) Haptoglobin polymorphism predicts 30-day mortality and heart failure in patients with diabetes and acute myocardial infarction. Diabetes. 9.
(21) 54:2802-2806. [22] Engstrom G, Lind P, Hedblad B, Wollmer P, Stavenow L, Janzon L, Lindgarde F. (2002) Lung function and cardiovascular risk: relationship with inflammation-sensitive plasma proteins. Circulation. 106:2555-2560. [23] Awadallah S, Hamad M. (2003) A study of haptoglobin phenotypes in patients with chronic renal failure. Ann. Clin. Biochem. 40:680-683. [24] Luo HD, Ramirez SP, Costa MD, Tan CT, Oakley RE, Lee CN, Hsu SI. (2004) Preoperative microalbuminuria, haptoglobin phenotype 2-2, and age are independent predictors for acute renal failure following coronary artery bypass graft. Ann. Acad. Med. Singapore. 33:S15-S16. [25] Burbea Z, Nakhoul F, Rosenberg S, Zoabi R, Skorecki K, Hochberg I, Miller-Lotan R, Benchetrit S, Weissgarten J, Knecht A, Tovbin D, Levy NS, Levy AP. (2004) Role of haptoglobin phenotype in end-stage kidney disease. Nephron. Exp. Nephrol. 97:e71-e76. [26] Miyoshi H, Ohshiba S, Matsumoto A, Takada K, Umegaki E, Hirata I. (1991) Haptoglobin prevents renal dysfunction associated with intravariceal infusion of ethanolamine oleate. Am. J. Gastroenterol. 86:1638-1641. [27] Zvi B, Levy AP. (2006) Haptoglobin phenotypes, which one is better and when. Clin. Lab. 52:29-35. [28] Hochberg I, Roguin A, Nikolsky E, Chanderashekhar PV, Cohen S, Levy AP. (2002) Haptoglobin phenotype and coronary artery collaterals in diabetic patients. Atherosclerosis. 161:441-446. [29] Wicher KB, Fries E. (2006) Haptoglobin, a hemoglobin-binding plasma protein, is present in bony fish and mammals but not in frog and chicken. Proc. Natl. Acad. Sci. U S A. 103:4168-4173. [30] Fagoonee S, Gburek J, Hirsch E, Marro S, Moestrup SK, Laurberg JM, Christensen EI, Silengo L, Altruda F, Tolosano E. (2005) Plasma protein haptoglobin modulates renal iron loading. Am. J. Pathol. 166:973-983. [31] Madsen M, Moller HJ, Nielsen MJ, Jacobsen C, Graversen JH, van den Berg T, 10.
(22) Moestrup SK. (2004) Molecular characterization of the haptoglobin.hemoglobin receptor CD163. Ligand binding properties of the scavenger receptor cysteine-rich domain region. J. Biol. Chem. 279:51561-51567. [32] Asleh R, Marsh S, Shilkrut M, Binah O, Guetta J, Lejbkowicz F, Enav B, Shehadeh N, Kanter Y, Lache O, Cohen O, Levy NS, Levy AP. (2003) Genetically determined heterogeneity in hemoglobin scavenging and susceptibility to diabetic cardiovascular disease. Circ. Res. 92:1193-1200. [33] Lim SK, Kim H, Lim SK, bin Ali A, Lim YK, Wang Y, Chong SM, Costantini F, Baumman H. (1998) Increased susceptibility in Hp knockout mice during acute hemolysis. Blood. 92:1870-1877. [34] Rademacher BE, Steele WJ. (1987) A general method for the isolation of haptoglobin 1-1, 2-1, and 2-2 from human plasma. Anal. Biochem. 160:119-126. [35] Wassdal I, Myrset AH, Helgeland L. (1991) A simple and rapid method for purification of rat haptoglobin for production of antiserum. Scand. J. Clin. Lab. Invest. 51:565-569. [36] Travis JC, Sanders BG. (1972) Haptoglobin evolution: polymeric forms of Hp in the Bovidae and Cervidae families. J. Exp. Zool. 180:141-148. [37] Morimatsu M, Syuto B, Shimada N, Fujinaga T, Yamamoyo S, Saito M, Naiki M. (1991) Isolation and characterization of bovine haptoglobin from acute phase sera. J. Biol. Chem. 266:11833-11837. [38] Katnik I, Jadach J. (1993) Immunoaffinity purification of human haptoglobin using monoclonal antibodies. Arch. Immunol. Ther. Exp. (Warsz). 41:303-308. [39] Jialal I, Fuller CJ. (1995) Effect of vitamin E, vitamin C and beta-carotene on LDL oxidation and atherosclerosis. Can. J. Cardiol. 11:Suppl G:97G-103G. [40] Klouche K, Morena M, Canaud B, Descomps B, Beraud JJ, Cristol JP. (2004) Mechanism of in vitro heme-induced LDL oxidation: effects of antioxidants. Eur. J. Clin. Invest. 34:619-625. [41] Alul RH, Wood M, Longo J, Marcotte AL, Campione AL, Moore MK, Lynch SM. (2003) Vitamin C protects low-density lipoprotein from homocysteine-mediated 11.
(23) oxidation. Free Radic. Biol. Med. 34:881-891. [42] Mao SJT, Yates MT, Parker RA, Chi EM, Jackson RL. (1991) Attenuation of atherosclerosis in a modified strain of hypercholesterolemic Watanabe rabbits with use of a probucol analogue (MDL 29,311) that does not lower serum cholesterol. Arterioscler. Thromb. 11:1266-1275. [43] Mao SJT, Yates MT, Rechtin AE, Jackson RL, Van Sickle WA. (1991) Antioxidant activity of probucol and its analogues in hypercholesterolemic Watanabe rabbits. J. Med. Chem. 34:298-302. [44] Sundell CL, Somers PK, Meng CQ, Hoong LK, Suen KL, Hill RR, Landers LK, Chapman A, Butteiger D, Jones M, Edwards D, Daugherty A, Wasserman MA, Alexander RW, Medford RM, Saxena U. (2003) AGI-1067: a multifunctional phenolic antioxidant, lipid modulator, anti-inflammatory and antiatherosclerotic agent. J. Pharmacol. Exp. Ther. 305:1116-1123.. 12.
(24) Section 1: A novel approach for Hp purification. 13.
(25) Abstract Similar to blood type, human plasma haptoglobin (Hp) is classified as 3 phenotypes: Hp 1-1, 2-1, or 2-2. The structural and functional relationship between the phenotypes, however, has not been studied in detail due to the complicated and difficult isolation procedures. This report provides a simple protocol that can be used to purify each Hp phenotype. Plasma was first passed through an affinity column coupled with a high affinity Hp monoclonal antibody. The bound material was washed with a buffer containing 0.2 M NaCl and 0.02 M phosphate, pH 7.4, eluted at pH 11, and collected in tubes containing 1 M Tris-HCl, pH 6.8. The crude Hp fraction was then chromatographed on a HPLC Superose 12 column in 0.05 M ammonium bicarbonate at a flow rate of 0.5 ml/min. The homogeneity of purified Hp 1-1, 2-1, or 2-2 was greater than 95% as judged by SDS polyacrylamide gel electrophoresis. Essentially, each Hp isolated was not contaminated with hemoglobin and apolipoprotein A-I as that reported from the other methods, and was able to bind hemoglobin. Neuraminidase treatment demonstrated that the purified Hp possessed a carbohydrate moiety, while Western blot analysis confirmed α and β chains corresponding to each Hp 1-1, 2-1, and 2-2 phenotype. The procedures described here represent a significant improvement in current purification methods for the isolation of Hp phenotypes. Circular dichroic spectra showed that the α-helical content of Hp 1-1 (29%) was higher than that of Hp 2-1 (22%) and 2-2 (21%). The structural difference with respect to its clinical relevance is discussed.. 14.
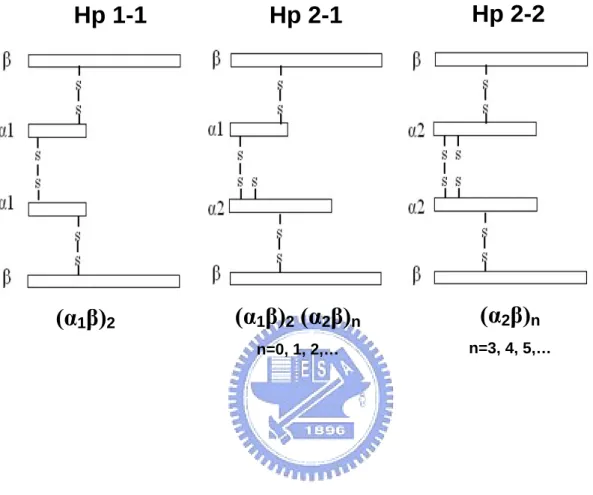
(26) Keywords: Human haptoglobin 1-1, 2-1, 2-2; Affinity purification; α-helix; Monoclonal antibodies. Introduction Hp is known as an acute phase protein, and its plasma level elevates in response to inflection or inflammation. For this reason, Hp is a useful indicator for some infectious diseases [1-3]. It is also a hemoglobin-binding protein present in the plasma of all vertebrates and believed to participate in hemoglobin transport [4]. Human Hp is a tetrameric structure joined by disulfide linkages among the 2 α and 2 β chains [4-6]. Based on the length of α chain, there are three phenotypes of Hp in the population, Hp 1-1, 2-1, and 2-2 (Fig. 1). All the phenotypes share the same 2 β chains (each with about Mr 40,000 dalton containing 245 amino acids and approximately 30% carbohydrate). A typical structure of homozygous Hp 1-1 is composed of two identical α1 chains (each with about Mr 9,000 dalton containing 83 amino acids). Homozygous Hp 2-2 is composed of two identical α2 chains (each with about Mr 16,500 dalton containing 142 amino acids) as compared to that of heterozygous Hp 2-1 containing one each of α2 and α1 (Fig. 1). Likewise, the tetrameric arrangement is also found in other animal species such as rat, rabbit, and pig [7-12]. However the 2 identical αβ units (Hp 1-1), joined by a non-covalent interaction rather than a disulfide. 15.
(27) bridge, are found in dog, cat, and bear [13-14]. Clinically, polymeric form of Hp 2-1 or 2-2 is associated with the complications of myocardial infarction [15], kidney failure [16], and diabetics [17]. Presumably, this was due to the complicated structure of Hp 2-1 and 2-2 as it forms heterogeneous polymers, in which some of the biologically functional groups are not fully expressed on the surface (Fig. 1). The assumption, however, has not been tested because the structural and functional studies are hampered by lack of a straightforward isolation procedure in preparing sufficient Hp phenotypes. The methods currently used for the purification of Hp frequently suffer some drawbacks. For example, Rademacher et al. utilize the chicken hemoglobin-Sepharose affinity column to isolate human Hp; the harsh-elution condition (8 M urea) causes the dissociation of a hemoglobin subunit from the Sepharose [18]. Meanwhile, human apolipoprotein A-I appears to be another major contaminant.. Wassdal et al. use rabbit hemoglobin-Sepharose; the. hemoglobin is still co-eluted from the column [19]. Travis et al. employ Sephadex G-200 gel filtration, but the purified Hp is accompanied with large amounts of IgM and α-2 macroglobulin [20].. Morimatsu et al. provide a modified method using HPLC with anion-exchange,. Sephacryl S-300, TSK Phenyl-5PW, and TSK DEAE-5PW columns together; the procedures however are time-consuming, and the yield is relatively low (2.5 mg per 130 ml acute phase serum) [21]. Although Katnik et al. have shown a single-step isolation for Hp using an antibody-affinity column, the phenotypes, final purity, and the biological properties of Hp are. 16.
(28) not fully reported [22]. Presumably, the purpose of their report was to use isolated Hp for raising monoclonal antibodies [22]. The similar antibody affinity-column procedure [22] was employed in our laboratory, but the isolated Hp was not pure. In the present report, we established simple two-step procedures for each Hp 1-1, 2-1, and 2-2 purification using a monoclonal antibody affinity-column followed by a HPLC Superose 12 gel filtration. Finally, some of the biochemical and physical properties with respect to each Hp phenotype were characterized and discussed.. Materials and methods Materials Goat polyclonal antibody against human Hp was purchased from Sigma (St. Louis, MO, USA). Rabbit anti-Goat IgG was purchased from Chemicon. CNBr-activated Sepharose 4B was purchased from Pharmacia. All other chemicals were purchased from Sigma (St. Louis, MO, USA) and Merck (Darmstadt, Germany) without any further purification. The buffers used in this report were all filtered through a 0.45 μm filter before using.. Preparation of monoclonal antibody against Hp Six monoclonal antibodies: 8B1-3A, W1-11G, 2-3H, G2D-7G, 12B-1 and 4A2-4H, against human Hp were produced and characterized according to the standard procedures established in. 17.
(29) our laboratory [23]. Monoclonal antibody 8B1-3A, which possessed the highest binding affinity to Hp, was selected for preparation of the affinity column. Briefly, 120 ml of cultured medium from the 8B1-3A hybridoma were first precipitated in 50% saturated ammonium sulfate. The precipitate was dissolved in 12 ml of phosphate buffered saline containing 0.02 M phosphate and 0.15 M NaCl, pH 7.4 (PBS). The solution was then dialyzed exhaustively in PBS to remove the remaining ammonium sulfate, followed by a dialysis in coupling buffer containing 0.1 M NaHCO3 and 0.5 M NaCl, pH 8.3.. Preparation of antibody affinity column Dialyzed monoclonal antibody was first coupled to CNBr-activated Sepharose-4B (Pharmacia, Uppsala, Sweden) according to the manufacturer’s procedures. Briefly, 2.86 g of freeze-dried Sepharose (1 g of freeze-dried powder gave about 3.5 ml final volume of gel) were swollen and suspended in 1 mM HCl and immediately washed with 20x volume of the same solution within 15 min on a sintered glass filter [24-26]. The gel was then washed with coupling buffer containing 0.1 M NaHCO3 and 0.5 M NaCl, pH 8.3, and degassed. About 10 ml (18.7 mg/ml) of ammonium-sulfate fraction of monoclonal antibody in coupling buffer were slowly added into the gel (in 15 ml), while gently stirring by a magnetic bar for 1 h at room temperature. After coupling, the gel was washed 10x volume of PBS to remove unbound materials via a sintered glass filter. The gel was then treated with a blocking solution containing. 18.
(30) 0.1 M Tris-HCl and 0.5 M NaCl, pH 8.0, for 2 h at room temperature to saturate the remaining reactive-sites. The degassed gel was then washed with 3 cycles of blocking buffer and a 0.15 M NaCl solution of pH 11.0 (adjusted by ammonium) according to the procedures previously described by us [26]. Finally, the gel was equilibrated in PBS and packed onto a 1.5 x 20 cm column.. Purification of human Hp using antibody affinity-column chromatography Initially, 1 ml of filtered human plasma of each Hp-phenotype batch was loaded onto the antibody affinity-column (10 ml in bed volume) at room temperature. The column was then washed with 50 ml of PBS. The bound materials were further washed with 50 ml of 0.02 M phosphate buffer containing 0.2 M NaCl, pH 7.4, and then eluted with 50 ml of a freshly prepared 0.15 M NaCl solution with pH 11 adjusted by ammonium [26]. Five ml of each fraction was collected in a tube containing 0.25 ml of 1 M Tris-HCl buffer, pH 6.8, to immediately neutralize the pH value. Pooled fractions containing Hp were then concentrated to a final volume of 1 ml using an Amicon centrifugal filter (Millipore).. Further purification of Hp by gel filtration column Concentrated solution with Hp was filtered with a 0.45 μm nylon fiber prior to HPLC. The HPLC system (Waters) consisted of two pumps, an automatic sample injector, and a. 19.
(31) photodiodearray detector. Superose 12 column (1 x 30 cm) (Pharmacia) was used for further Hp purification. The column was pre-equilibrated with 50 mM ammonium bicarbonate. Partially purified Hp (0.8 ml) was applied to the column at a flow rate of 0.5 ml/min. Fractions containing Hp were pooled and concentrated to a final volume of 1 ml using an Amicon centrifugal filter and then lyophilized. The lyophilized Hp was stored at –80℃ until analyzing.. Gel electrophoresis and densitometry Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed according to the Laemmli’s method [27] with some modification in using 5% polyacrylamide (w/v) on the stacking gel as previously described [7]. Samples (typically 5 μg) for SDS-PAGE were preheated at 100 ℃ for 10 minutes in a loading buffer [12 mM Tris-HCl, pH 6.8, 0.4% SDS (w/v), 5% glycerol (v/v), 2.88 mM 2-mercaptoethanol, 0.02% bromphenol blue (w/v)]. For molecular weight calibration, a subset of the following standards was included in each gel: myosin (200 kDa), β-galactosidase (116 kDa), phosphorylase B (97 kDa), serum albumin (66 kDa), ovalbumin (45 kDa), carbonic anhydrase (31 kDa), soybean trypsin inhibitor (21.5 kDa), lysozyme (14.4 kDa), and aprotinin (6.5 kDa). The samples were run for about 1.5 h at 100 V and stained using Coomassie brilliant blue R-250. Densitometric analysis of SDS-PAGE was performed using a Molecular Dynamics densitometer for data acquisition and Image Quant software for integration and analysis.. 20.
(32) Immunoblot analysis Following the separation of proteins by SDS-PAGE, the gel and nitrocellulose- and 3MM filter- papers were soaked in a transfer buffer containing 48 mM Tris-HCl, 39 mM glycine, 0.037% SDS (w/v), and 20% methanol (v/v) at pH 8.3 for 30 min. The gel was then electrotransferred to a nitrocellulose membrane (Pharmacia) at 90 mA for 1 h in a semi-dry transfer cell (Bio-Rad) containing a transfer buffer. The transferred membrane was then immersed in 5% skim milk (w/v) in PBS for 1 hour at room temperature while shaking gently. After three times washing with PBS for 5 min, the membrane was incubated with a primary goat polyclonal antibody against human Hp [1:5000 dilution in PBS washing buffer containing 1% (w/v) skim milk and 0.05% Tween-20 (v/v) for 1 hour] at room temperature and washed three times for 5 min. The membrane was then incubated with 1:10,000 diluted rabbit anti-goat IgG conjugated with horseradish peroxidase in washing buffer for 1 h. In addition, the membrane was washed two times with washing buffer and further washed one time with PBS. Finally, the membrane was developed using 3,3’-diaminobenzidine (DAB) as a substrate for horseradish peroxidase [7, 25].. Circular dichroic spectra The lyophilized Hp was dissolved in 10 mM phosphate buffer at pH 7.4 with a final. 21.
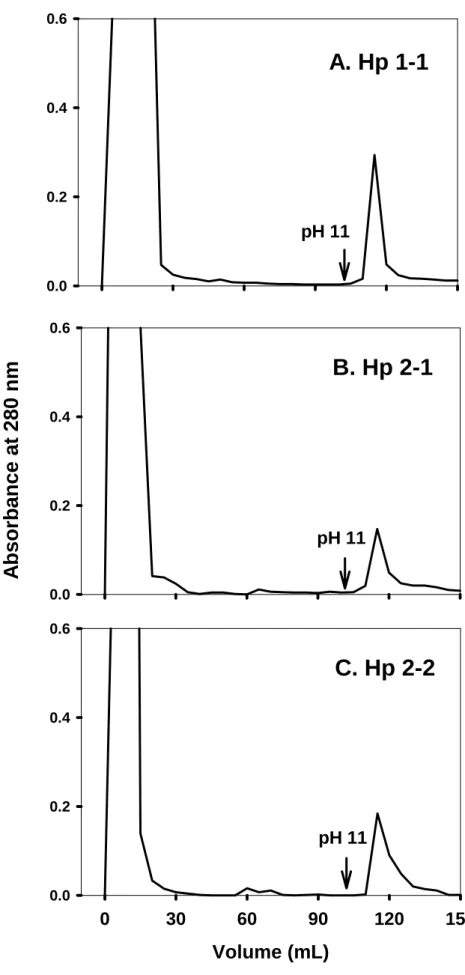
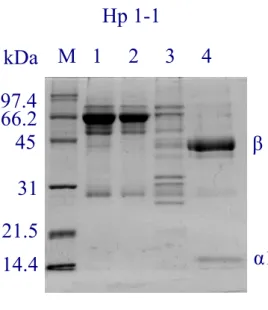
(33) concentration of 0.2 mg/ml. About 300 μl of Hp solution was used to analyze within a cuvette of 1-mm path length. Circular dichroic spectrum was conducted between 190 and 300 nm in a Jasco J-715 spectropolarimetry. The obtained spectrum of each type of Hp was accumulated for 20 times at a scanning rate of 50 nm/min and the % α-helical content was estimated from the mean residue molar elliplicity (θ222). % α-helix = [(θ222 + 3000)/(36000+3000)] X 100 [28].. Results Preparation of monoclonal antibody against Hp Six monoclonal antibodies prepared against Hp were characterized, in which 8B1-3A possessed the highest binding affinity (Ka=5.6 x 109 M-1) and was chosen to prepare an affinity column. The binding capacity estimated was greater than 100 μg of Hp per ml of coupled Sepharose (data not shown).. Purification of human Hp using antibody affinity column chromatography Fig. 2 shows a typical chromatographic profile for Hp 1-1, 2-1, and 2-2 purification on the affinity column. Human plasma was applied to the column followed by an extensive wash with a phosphate buffer containing 0.2 M NaCl. It is worth mentioning, this pre-wash step differed from the conventional method in which 0.12 M NaCl was used. Using 0.2 M NaCl, most of the low-affinity binding proteins were eliminated (Fig. 3). Hp was then eluted at pH 11 and. 22.
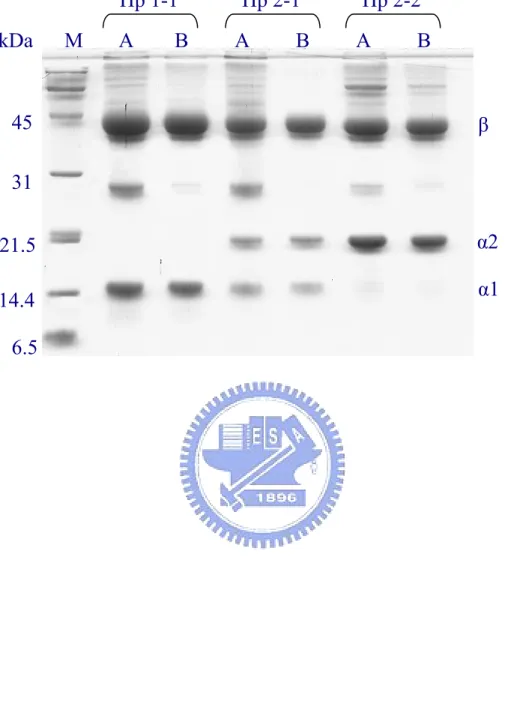
(34) collected in tubes containing 0.25 ml of 1 M Tris-HCl, pH 6.8, to immediately neutralize the pH. The purity of each Hp phenotype was approximately 60-80% in homogeneity as analyzed on SDS-PAGE. Apolipoprotein A-I appeared to be a major contaminant. All the phenotypes of Hp converted to α (α1 or α2 or both) and β subunits in the presence of the reducing reagent (Fig. 3). A typical Western blot analysis showing 3 isolated phenotypes is depicted in Fig. 4. The recovery of Hp at this step accounted for 75-94% of the Hp from the plasma with a final of 51-54 fold purification (Table 1).. Further purification of Hp on HPLC gel-filtration column The obtained Hp 1-1, 2-1, or 2-2 fraction was concentrated and applied onto a gel-filtration Superose 12 column pre-equilibrated with 0.05 M of ammonium bicarbonate, pH 8.0. Chromatographic profiles (Fig. 5) revealed that the solution property of each Hp phenotype was consistent with its molecular form, in which Hp 1-1 was more homogeneous in size with longer elution time than that of Hp 2-1 and 2-2. Purity of each phenotype was then analyzed on SDS-PAGE containing reducing reagent 2-mercaptoethanol. Homogeneity of each phenotype was greater than 95% (Fig. 6). Thus, HPLC Superose column was markedly effective to remove apoA-I contaminant. Western blot analysis in the absence of a reducing reagent demonstrated that Hp 2-1 and 2-2 were all polymeric (Fig. 7), in which Hp 2-2 was devoid of monomer and dimer consistent with. 23.
(35) the proposed structure of Hp (Fig. 1). Thus, our purification procedures did not apparently alter the structural characteristics of Hp phenotypes.. The polymeric structure of isolated Hp and its binding to hemoglobin We further studied the ionic property of isolated Hp 1-1, 2-1, and 2-2 on a native-PAGE; the distinct polymorphism of each phenotype was also observed (Fig. 8). Hp 2-2 was the most basic among the Hp phenotypes. Since hemoglobin (Hb) is able to bind Hp and to form a Hb-Hp complex [10], Fig. 8 demonstrates that the Hb could form Hb-Hp complex with each Hp phenotype we isolated.. Circular dichroic spectra To characterize the secondary structure of each Hp phenotype, we determined the conformation of Hp by CD (Fig. 9). The estimated α-helical content was about 29, 22, and 21% for Hp 1-1, 2-1, and 2-2, respectively (Table 2). Statistically, the α-helical content in Hp 1-1 was significantly higher than that in Hp 2-1 and 2-2 (P < 0.001).. Discussion Purification of human Hp has been hampered for years due to its structural diversity as described above [7, 18-21]. In the present study, plasma without any additional manipulations. 24.
(36) (e.g., ammonium-sulfate precipitation) was utilized for Hp isolation. With the use of a 0.2 M NaCl wash in our procedures, most of the nonspecific low-affinity binding proteins were eluted from the column. Nevertheless, the affinity purified Hp 1-1, 2-1 or 2-2 analyzed on SDS-PAGE was only 60-80% pure (Fig. 3). HPLC Superose 12 column appeared to be superior to the other methods in the second-step purification since apolipoprotein A-I, a major contaminant, and other unknown high molecular-weight proteins were almost eliminated. The solution property of each Hp phenotype on this Superose column was consistent with its molecular forms, of which Hp 1-1 was more homogeneous than the other two species in size. Notably, the elution time of each phenotype was also consistent with the size of Hp, in which the polymeric form of Hp 2-2 and 2-1 was eluted earlier, respectively (Fig. 5). Western blot analysis on the polymeric structure of isolated Hp using 6% SDS-PAGE without reducing reagent (Fig. 7) revealed its structural identity to that originally present in plasma (data not shown). It also confirmed that the isolated Hp 2-1 was heterogeneous in nature containing Hp monomer and dimer, while the polymeric Hp 2-2 contained neither. Subsequently, we show each phenotype of isolated Hp possessed the ability to bind hemoglobin (Fig. 8), although we cannot presently address whether or not the binding potency might be attenuated during the isolation.. Meanwhile, we demonstrated the presence of. carbohydrate moiety in isolated Hp as neuraminidase treatment could remove the terminal sialic acid residues from the Hp with a time-dependent manner similar to our previous study [7]. 25.
(37) (data not shown). Taking together, the Hp isolated from the antibody affinity-column combined with HPLC Superose 12 exhibited advantages over the conventional methods. First, the heterogeneous particles of Hp 2-1 or 2-2 could be obtained in one pool as polymeric forms. Second, the co-eluted hemoglobin from hemoglobin affinity-column [19] was eliminated in the present study. Using an ELISA for hemoglobin assay, we could not detect hemoglobin in the Hp we isolated (data not shown).. Third, the isolated Hp not only retained its ability to bind. hemoglobin (Fig. 8), but also blocked the peroxidase activity of hemoglobin (data not shown). The CD spectrum of each Hp phenotype was determined to estimate the secondary structure of Hp. For the first time, according to the best of our knowledge, we showed that the α-helical content of Hp 1-1, 2-1, and 2-2 was 29, 22, and 21%, respectively. Hp 1-1 possessed a more ordered structure than that of polymeric forms of Hp 2-1 and 2-2. Moreover, we identified that the immunoreactivity of Hp 1-1 was also greater than that of Hp 2-1 and 2-2. For example, the monoclonal antibody used in our affinity column (8B1-3A) exhibited an affinity (Ka) 5.58 x 109, 2.81 x 109, and 2.19 x 109 M-1 against Hp 1-1, 2-1, and 2-2, respectively. The later result suggests that the availability of “functional” surfaces of Hp 1-1 may be greater as compared to Hp 2-1 and 2-2. Such structural differences may explain, in part, the clinical outcome by which Hp phenotype is associated with differential susceptibility to infections, atherosclerosis, and autoimmune disorders [3, 29]. These effects are correlated with a phenotype-dependent. 26.
(38) modulation of oxidative stress and prostaglandin synthesis. In general, patients with Hp 2-2 are more susceptible in developing the severity of the diseases mentioned above. Identification of the biochemical basis for the differences among Hp phenotypes may lead to a rational design in intervening new pharmacological agents, such as mini-Hp, which have been recently proposed [30]. In conclusion, due to the structural heterogeneity, it is difficult to purify Hp from human plasma, particularly with the Hp 2-1 and 2-2 polymers. The present report provides a simple method for the purification of Hp phenotypes with relatively high yield. Hp 1-1, 2-1, and 2-2 can therefore be prepared and used for the study of structural and functional relationship with the pathogenesis in the diseases of interest.. 27.
(39) References. [1]. Raijmakers MT, Roes EM, Morsche RHM te, Steegers EAP, Peters WHM. (2003) Haptoglobin and its association with the HELLP syndrome. J. Med. Genet. 40:214-216.. [2]. Engstrom G, Stavenow L, Hedblad B, Lind P, Eriksson KF, Janzon L, Lindgarde F. (2003) Inflammation-sensitive plasma proteins, diabetes, and mortality and incidence of myocardial infarction and stroke: a population-based study. Diabetes 52:442-447.. [3]. Levy AP, Hochberg I, Jablonski K, Resnick HE, Lee ET, Best L, Howard BV. (2002) Haptoglobin phenotype is an independent risk factor for cardiovascular disease in individuals with diabetes: The Strong Heart Study. J. Am. Coll. Cardiol. 40:1984-1990.. [4]. Kristiansen M, Graversen JH, Jacobsen C, Sonne O, Hoffman H, Law SKA, Moestrup SK. (2001) Identification of the haemoglobin scavenger receptor. Nature 409:198-201.. [5]. Maeda N, Smithies O. (1986) The evolution of multigene families: human haptoglobin genes. Annu. Rev. Genet. 20:81-108.. [6]. Patzelt D, Geserick G, Schroder H. (1988) The genetic haptoglobin polymorphism: relevance of paternity assessment. Electrophoresis 9:393-397.. [7]. Yang SJ, Mao SJT. (1999) Simple high-performance liquid chromatographic purification procedure for porcine plasma haptoglobin. J. Chromatogr. B 731:395-402.. [8]. Hanley JM, Haugen TH, Heath EC. (1983) Biosynthesis and processing of rat haptoglobin. J. Biol. Chem. 258:7858-7869.. [9]. Fraser IH, Smith DB. (1971) Studies on porcine haptoglobin and its complex with human hemoglobin. Can. J. Biochem. 49:141-147.. [10] Baumann H, Jahreis GP. (1983) Regulation of mouse haptoglobin synthesis. J. Cell. Biol. 97:728-736. [11] Chow V, Kurosky A, Murrary RK. (1984) Studies on the biosynthesis of rabbit haptoglobin. J. Biol. Chem. 259:6622-6629. [12] Busby WHJr, Travis JC. (1978) Structure and evolution of artiodactyla haptoglobins. 28.
(40) Comp. Biochem. Physiol. B. 60:389-396. [13] Kurosky A, Hay RE, Bowman BH. (1979) Canine haptoglobin: a unique haptoglobin subunit arrangement. Comp. Biochem. Physiol. B. 62:339-344. [14] Mominoki K, Nakagawa-Tosa N, Morimatsu M, Syuto B, Saito M. (1995) Haptoglobin in Carnivora: a unique molecular structure in bear, cat and dog haptoglobins. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 110:785-789. [15] Engstrom G, Lind P, Hedblad B, Wollmer P, Stavenow L, Janzon L, Lindgarde F. (2002) Lung function and cardiovascular risk: relationship with inflammation-sensitive plasma proteins. Circulation 106:2555-2560. [16] Miyoshi H, Ohshiba S, Matsumoto A, Takada K, Umegaki E, Hirata I. (1991) Haptoglobin prevents renal dysfunction associated with intravariceal infusion of ethanolamine oleate. Am. J. Gastroenterol. 86:1638-1641. [17] Hochberg I, Roguin A, Nikolsky E, Chanderashekhar PV, Cohen S, Levy AP. (2002) Haptoglobin phenotype and coronary artery collaterals in diabetic patients. Atherosclerosis 161:441-446. [18] Rademacher BE, Steele WJ. (1987) A general method for the isolation of haptoglobin 1-1, 2-1, and 2-2 from human plasma. Anal. Biochem. 160:119-126. [19] Wassdal I, Myrset AH, Helgeland L. (1991) A simple and rapid method for purification of rat haptoglobin for production of antiserum. Scand. J. Clin. Lab. Invest. 51:565-569. [20] Travis JC, Sanders BG. (1972) Haptoglobin evolution: polymeric forms of Hp in the Bovidae and Cervidae families. J. Exp. Zool. 180:141-148. [21] Morimatsu M, Syuto B, Shimada N, Fujinaga T, Yamamoyo S, Saito M, Naiki M. (1991) Isolation and characterization of bovine haptoglobin from acute phase sera. J. Biol. Chem. 266:11833-11837. [22] Katnik I, Jadach J. (1993) Immunoaffinity purification of human haptoglobin using monoclonal antibodies, Arch. Immunol. Ther. Exp. (Warsz). 41:303-308. [23] Mao SJT, Rechtin AE, Krstenansky JL, Jackson RL. (1990) Characterization of a monoclonal antibody specific to the amino terminus of the alpha-chain of human fibrin. 29.
(41) Thromb. Haemost. 63:445-448. [24] Chu LF, Lee WC, Yang PC, Chu R, Huang TY, Mao SJT. (1997) One-step HPLC purification procedure for porcine brain 90-kDa heat shock protein. Protein Expres. Purif. 10:180-184. [25] Liau CY, Chang TM, Pan JP, Chen WL, Mao SJT. (2003) Purification of human plasma haptoglobin by hemoglobin-affinity column chromatography. J. Chromatogr. B. 790:209-216. [26] Mao SJT, Miller JP,. Gotto AMJr, Sparrow JT. (1980) The antigenic structure of apolipoprotein A-I in human high density lipoproteins. Radioimmunoassay using surface-specific antibodies. J. Biol. Chem. 255:3448-3453. [27] Laemmli UK. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685. [28] Mao SJT, Sparrow JT, Gotto AMJr, Jackson RL. (1980) The phospholipid-binding and immunochemical properties of amidinated, guanidinated and acetylated apolipoprotein A-II. Biochim. Biophys. Acta. 617:245-253. [29] Bernard DR, Langlois MR, Delanghe JR, De Buyzere ML. (1997) Evolution of haptoglobin concentration in serum during the early phase of acute myocardial infarction. Eur. J. Clin. Chem. Clin. Biochem. 35:85-88. [30] Melamed-Frank M, Lache O, Enav BI, Szafranek T, Levy NS, Ricklis RM, Levy AP. (2001) Structure-function analysis of the antioxidant properties of haptoglobin. Blood 98:3693-3698.. 30.
(42) Figure Legends. Fig. 1. Schematic drawing of proposed structure of human Hp 1-1, 2-1, and 2-2. All three phenotypes share a common structure of β chains (please also see reference 4). The degree of polymerization within the inter-molecular arrangement is shown.. Fig. 2. Typical purification profile of human Hp on antibody affinity-column. One ml of human plasma was applied to an antibody affinity-column (pre-equilibrated with PBS, pH 7.4) followed by a wash in 10 mM phosphate buffer containing 0.2 M NaCl, pH 7.4. The bound Hp was then eluted in a solution at pH 11 and collected in tubes containing 0.25 ml of 1 M Tris, pH 6.8.. Fig. 3. Analysis of isolated Hp from affinity column using 12% SDS-PAGE in the presence of reducing reagent. Lane M: molecular markers (expressed as kDa). Lanes 1-2: plasma before and after flowing through the affinity column, respectively. Lane 3: low-affinity binding proteins washed with 10 mM phosphate buffer containing 0.2 M NaCl, pH 7.4. Lane 4: the eluted Hp as described in Fig. 2.. Fig. 4. Western blot analysis of Hp isolated from antibody affinity-column. Following a 12%. 31.
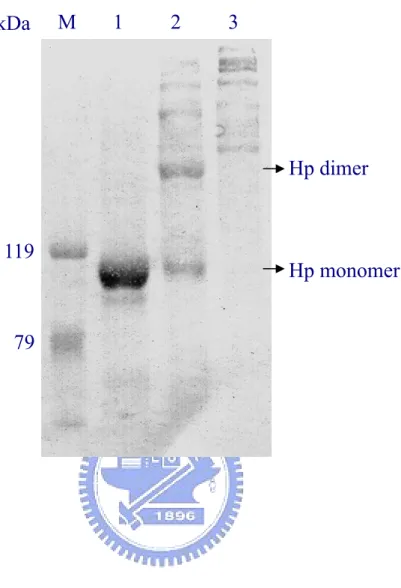
(43) SDS-PAGE, the separated proteins were transferred, blocked, and developed by a goat anti-human Hp. Lane M: molecular markers. Lanes 1-2: plasma before and after flowing through the affinity column, respectively. Lane 3: low-affinity binding proteins washed with 10 mM phosphate buffer containing 0.2 M NaCl, pH 7.4. Lane4: the eluted Hp.. Fig. 5. Typical chromatographic profile of affinity isolated Hp on HPLC Superose 12 column. Isolated Hp 1-1, 2-1, or 2-2 from the affinity column was first concentrated and applied to a HPLC Superose 12 pre-equilibrated in 50 mM of ammonium bicarbonate, pH 8.0. The same solution was used in the mobile phase at a flow rate of 0.5 ml/ml. The filled bar represents the pooled fractions corresponding to isolated Hp.. Fig. 6. Analysis of purified Hp from antibody affinity and HPLC gel filtration column on 15% SDS-PAGE in the presence of reducing reagent. M: molecular markers. A: isolated Hp from affinity column alone. B: purified Hp from an additional separation on HPLC Superose 12 column.. Fig. 7. Western blot analysis of polymeric structure of purified Hp on 6% SDS-PAGE in the absence of reducing reagent. Lane M: molecular markers. Lanes 1-3: purified Hp 1-1, 2-1, and 2-2, respectively. Notably, the Hp 2-1 contains monomeric and dimeric forms of Hp 1-1:. 32.
(44) whereas the Hp 2-2 is devoid of both.. Fig. 8. Analysis of hemoglobin-binding property of purified Hp 1-1, 2-1, and 2-2 on a 7% native-PAGE. Briefly, each Hp phenotype (5 μg) was incubated with and without hemoglobin (Hb) (5 μg) at room temperature for 30 minutes before conducting the native-PAGE. The gel was then stained with Coomassie Blue R-250. Hp of each phenotype was shifted to basic upon the binding of Hb.. Fig. 9. Circular dichroic spectra of Hp 1-1, 2-1, and 2-2. Each Hp in 10 mM phosphate buffer, pH 7.4, at a final concentration of 0.2 mg/ml was monitored by a circular dichroism. Each spectrum represents a mean of 20x determinations.. 33.
(45) Fig 1. Hp 1-1. (α1β)2. Hp 2-1. Hp 2-2. (α1β)2 (α2β)n. (α2β)n. n=0, 1, 2,…. n=3, 4, 5,…. 34.
(46) Fig 2. 0.6. A. Hp 1-1 0.4. 0.2. pH 11 0.0. Absorbance at 280 nm. 0.6. B. Hp 2-1 0.4. 0.2. pH 11 0.0 0.6. C. Hp 2-2 0.4. 0.2. pH 11 0.0. 0. 30. 60. 90. Volume (mL). 35. 120. 150.
(47) Fig 3.. Hp 1-1 kDa M 1. 2. 3. 4. 97.4 66.2 45. β. 31 21.5 α1. 14.4. Hp 2-1 kDa M. 1. 2. 3. 4. 97.4 66.2 45. β. 31 21.5. α2. 14.4. α1 Hp 2-2. kDa M. 1. 2. 3. 4. 97.4 66.2 45. β. 31 21.5 14.4. α2. 36.
(48) kDa M. Hp 1-1 1 2 3. 4. Fig 4.. 119 79 46. β. 31 24 19 α1. kDa M 119 79. Hp 2-1 1 2 3. 4. 46. β. 31 24 α2. 19. kDa. α1. M. Hp 2-2 1 2 3. 4. 119 79 46. β. 31 24 α2. 19. 37.
(49) Fig 5.. 38.
(50) Fig 6. Hp 1-1 kDa. M. A. B. Hp 2-1 A. B. 45. Hp 2-2 A. B. β. 31 α2. 21.5. α1. 14.4 6.5. 39.
(51) Fig 7. kDa. M. 1. 2. 3. Hp dimer. 119. Hp monomer. 79. 40.
(52) Fig 8.. Hp 1-1 Hb. +. —. +. Hp 2-1 —. +. Hp 2-2 —. +. Hb. 41.
(53) [θ]MRW x 10-3 (mdeg x cm2 X dmol-1 x residue-1). Fig 9.. 8 6 4 2 0 -2 -4 -6. Hp 1-1 Hp 2-1 Hp 2-2. -8 -10 -12 180. 200. 220. 240. 260. Wavelength (nm). 42. 280. 300. 320.
(54) Table 1. Analytical recovery and yield of haptoglobin purified from 1 ml of human plasma (A) Hp 1-1 Plasma. Affinity column. Gel filtration. Total protein (mg). 102. 1.41. 0.88. Moiety of Hp (mg). 1.46. a. 1.09. b. 0.81. 1.43. 77. 92. 1. 54. 64. Recovery (%). 100. 75. 55. Yield (mg). —. —. 0.81. Plasma. Affinity column. Gel filtration. 101. 1.80. 0.72. b. b. Purity (%). Fold purification. b. (B) Hp 2-1. Total protein (mg). a. 1.17. 1.10. 0.71. 1.16. 61. 98. 1. 53. 84. Recovery (%). 100. 94. 61. Yield (mg). —. —. 0.71. Plasma. Affinity column. Gel filtration. 100. 1.53. 0.81. b. Moiety of Hp (mg) b. Purity (%). Fold purification. b. (C) Hp 2-2. Total protein (mg) Moiety of Hp (mg) b. Purity (%). Fold purification. a. 1.26. 0.98. 0.76. b. 1.26. 64. 94. 1. 51. 75. Recovery (%). 100. 78. 60. Yield (mg). —. —. 0.76. a. The concentration of each human Hp phenotype was determined using an ELISA currently used in our laboratory. b Determined by densitometer using digital Image Quant software.. 43.
(55) Table 2. α-helical content of each Hp phenotype as determined by CD Hp 1-1 α-helix. 29%. *. Hp 2-1. Hp 2-2. 22%. 21%. *Significant difference as compared to Hp 2-1 or Hp 2-2 (P < 0.001).. 44.
(56) Section 2:. Study of antioxidant role of Hp in LDL oxidation and in transfected cells. 45.
(57) Abbreviations:. AAPH,. 2,2’-azobis(2-amidinopropane)-dihydrochloride;. ABTS,. 2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid); Hp, haptoglobin; Hb, hemoglobin; CAD, cardiovascular disease; CD, circular dichroism; CHO, Chinese hamster ovary; CMV, cytomegalovirus; CM, carboxymethylated, DAB, 3,3’-diaminobenzidine; LDL, low density lipoprotein; HDL, high density lipoprotein; HRP, horseradish peroxidase; MDA, malonaldehyde; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; TBA, 2-thiobarbituric acid; TBARS, thiobarbituric acid-reactive substance. Keywords: Haptoglobin phenotypes / Antioxidant / Hemoglobin / Haptoglobin cDNA / Physiologic role. 46.
(58) Abstract Similar to blood type, human plasma haptoglobin (Hp) is classified as 3 phenotypes: Hp 1-1, 2-1, and 2-2 attributed by their two common alleles 1 and 2.. Clinically, the phenotype 2-2 is. associated with the risk in patients with cardiovascular diseases and diabetes mellitus. In this study, we demonstrated that Hp was an extremely potent antioxidant, which directly prevented LDL from Cu2+-induced oxidation. probucol: one of the most potent antioxidants.. Its potency was markedly superior to The IC50 of antioxidant activity in ranking. was: Hp 1-1> Hp 2-1> Hp 2-2> probucol> vitamin E.. Blockage of disulfide linkages. between Hp subunits, not only abolished the α-helical content but also diminished its ability to form a complex with hemoglobin (Hb). than that of native Hp.. It exerted almost 4 x greater antioxidant activity. To investigate antioxidant role of Hp on the cellular level, the cDNA. of Hp 1-1 was cloned, constructed (containing the pcDNA3.0 vector with CMV promoter) and transfected to CHO-K1 cells. Following the transfection, these CHO cells were able to express Hp 1-1 protein and significantly (P<0.001) elevated the tolerance against the oxidative stress.. The elevation was about twice-higher than that normal CHO cells when. challenged with hydrogen peroxide for 24 h. Thus, Hp plays a provocative antioxidant role as demonstrated in our in vitro and ex vivo studies.. 47.
(59) Introduction Hemoglobin (Hb) is the most abundant and functionally important protein in erythrocytes. However, once Hb released from red blood cells, it becomes highly toxic because of the oxidative nature of iron-containing heme, which participates in the Fenton reaction to produce reactive oxygen species causing cell injury [1-2].. On the other hand, human plasma. haptoglobin (Hp), known as an acute phase protein [3-5], may capture the Hb by forming a high affinity Hp-Hb complex [1, 3-4].. The complex is then metabolized through a. receptor-mediated process including a recent report showing the CD 163 receptor of macrophages [6]. the plasma [7].. Therefore, Hb binding by Hp is essential in rapid clearance of Hb from For this reason, Hp plays a crucial role against Hb-induced oxidative stress. by a mechanism thought to be from its high-affinity binding with Hb and prevent the iron “leaking” from the Hb.. However, thus far, there is no report directly pointing out that Hp. itself is an antioxidant molecule.. The different Hp phenotype 1-1, 2-1, or 2-2 in each respective individual is attributed by two common alleles 1 and 2 located at chromosome 16q22.1.. Structurally, the minimal unit of. Hp (β-α-α-β) is joined by disulfide linkages among the 2 α and 2 β chains [6, 8-9].. Fig. 1. shows that all the phenotypes share the same 2 β chains (each with about MW 40,000 dalton containing 245 amino acids and about 30% carbohydrate).. A typical structure of. homozygous Hp 1-1 (β-α1-α1-β) is composed of only two identical α1 chains (each with. 48.
(60) about MW 9,000 dalton containing 83 amino acids).. Homozygous Hp 2-2 is composed of. two identical α2 chains (each with about MW 16,500 dalton containing 142 amino acids) as compared to that of heterozygous Hp 2-1 containing each α2 and α1 (Fig. 1).. Due to an. extra-thio group in α-2 chain, only Hp 2-1 and 2-2 form large polymers in monomeric, trimeric, tetrameric, pentameric, hexameric, and even larger arrangement through the disulfide-linkages (Fig. 1).. Clinically, diabetic patients with the Hp 1-1 type are markedly resistant to the development of diabetic retinopathy, diabetic nephropathy, and cardiovascular disease (CAD) [10-12].. In a. prospective study, participants homozygous with Hp 2-2 are 5-fold increased in risk for the development of CAD as compared to Hp 1-1, whereas the risk in heterozygous Hp 2-1 are intermediate [13].. In the present study, we show that Hp was an extremely potent antioxidant and the activity of Hp 1-1 was differentially and moderately greater than that of Hp 2-1 and 2-2. We also demonstrated that Hp 1-1 cDNA transfected Chinese hamster ovary (CHO) K1 cells (normally not expressing the Hp molecules) significantly resist against oxidative stress. role of Hp as an antioxidant molecule is therefore identified.. The clinical significance with. respect to the structure and function of Hp phenotype is discussed.. 49. The.
(61) Materials and methods Purification of human Hp Hp phenotype was first identified using plasma supplemented with hemoglobin (Hb) on a native polyacrylamide gel electrophoresis (PAGE), followed by a peroxidase substrate staining [14].. The plasma of each specific Hp phenotype was then chromatographed on an. antibody affinity-column followed by a gel filtration chromatography as previously described by our laboratory [14-15].. The homogeneity of each Hp species employed was greater than. 95%.. Gel electrophoresis Sodium dodecyl sulfate- polyacrylamide gel electrophoresis (SDS-PAGE) containing 15% polyacrylamide (w/v) with a top stacking gel (5% polyacrylamide) was performed for Hp characterization [14, 16].. Samples (typically 5 μg) for SDS-PAGE were preheated at 100℃. for 10 minutes in a loading buffer [12 mM Tris-HCl, pH 6.8, 0.4% SDS (w/v), 5% glycerol (v/v), 0.02% bromphenol blue (w/v)] with/without 2.88 mM 2-mercaptoethanol.. The. samples were run for about 1.5 h at 100 V and stained using Coomassie Brilliant Blue R-250.. Western blot Following the separation of proteins by SDS-PAGE, the gel soaked in a transfer buffer containing 48 mM Tris-HCl, 39 mM glycine, 0.037% SDS (w/v), and 20% methanol (v/v) at. 50.
(62) pH 8.3 was electrotransferred to a nitrocellulose paper (Pharmacia) at 90 mA for 1 h in a semi-dry transfer cell (Bio-Rad).. Immunoblotting and developing were conducted according. the standard procedures previously described [14-16].. Reduction and carboxymethylation of Hp To 1 mg of Hp was added 3.3 mL of 0.01 M Tris-HCl buffer (pH 8.6) containing 5.4 M urea, and 1% (v/v) β-mercaptoethanol. incubated at room temperature.. The reaction mixture was flushed with nitrogen and. After 2 h, 20 mg of iodoacetic acid were added and the. reaction mixture was then maintained at pH 8.6 by the addition of 1 M NaOH for 30 min. Finally, carboxymethylated (CM) Hp was desalted on a Bio-Gel P2 column eluted with 0.1 M ammonium bicarbonate and lyophilized [17].. Preparation of LDL Human low density lipoprotein (LDL) (d. 1.012-1.063 g/mL) was prepared from human plasma by a sequential ultracentrifugation according to the method previously established [17]. Sodium azide (0.01%) was added into plasma prior to ultracentrifugation and into LDL after isolation. Subsequently, freshly prepared LDL was dialyzed against PBS to remove EDTA and used for oxidation assays [1].. LDL oxidation Thiobarbituric acid-reactive substances (TBARS) were used as an index to measure the LDL. 51.
數據
+7
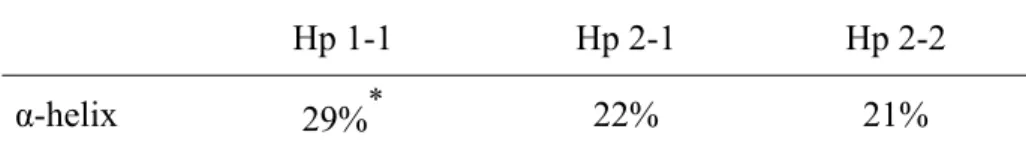

相關文件
² Stable kernel in a goals hierarchy is used as a basis for establishing the architecture; Goals are organized to form several alternatives based on the types of goals and
Nonsmooth regularization induces sparsity in the solution, avoids oversmoothing signals, and is useful for variable selection.. The regularized problem can be solved effectively by
The hashCode method for a given class can be used to test for object equality and object inequality for that class. The hashCode method is used by the java.util.SortedSet
• A narrative poem is a poem that tells a story. Narrative poems can come in many forms and styles. They can be long or short, simple or complex, as long as they tell stories.
It is the author’s hope that the Zuting shiyuan may be effectively used as a supplement for understanding Chan texts, and its contributions be fully valued.. Furthermore, the
To explore different e-learning resources and strategies that can be used to successfully develop the language skills of students with special educational needs in the
Explore different e-learning resources and strategies that can be used to successfully develop the language skills of students with special educational needs in the..
* All rights reserved, Tei-Wei Kuo, National Taiwan University, 2005..
