Original research study
Combined effects of differentiation factor 15 and substance use of alcohol, betel quid and cigarette on risk of head and neck cancer
SL Chiang1, CP Lee2, JG Chang3,4, CH Lee5, KT Yeh6, YS Tsai7, MK Chen8, CH Chen9, YC Ko1,7,10*
* Corresponding author
Email:
[email protected]1 Environment-Omics-Diseases Research Center, China Medical University Hospital, Taichung,
Taiwan
2 Graduate Institute of Medicine, College of Medicine, Kaohsiung Medical University, Kaohsiung,
Taiwan
3 Department of Laboratory Medicine, China Medical University Hospital, Taichung, Taiwan
4 Center of RNA Biology and Clinical Application, China Medical University Hospital, Taichung,
Taiwan
5 Department of Public Health, College of Health Science, Kaohsiung Medical University,
Kaohsiung, Taiwan
6 Department of Pathology, Changhua Christian Hospital, Changhua, Taiwan
7 Center of Excellence for Environmental Medicine, Kaohsiung Medical University, Kaohsiung,
Taiwan
8 Department of Otorhinolaryngology, Head and Neck Surgery, Changhua Christian Hospital,
Changhua, Taiwan
9 School of Dentistry, College of Dental Medicine, Kaohsiung Medical University, Kaohsiung,
Taiwan
10 Graduate Institute of Clinical Medical Science, College of Medicine, China Medical University,
Abstract
Growth differentiation factor 15 (GDF15) is susceptible to areca nut exposure in oral cells and
contributes to the progression of oral tumours. Here, we determined whether independent and
combined effects between GDF15 and substance use of alcohol, betel quid and cigarette (ABC)
influence the incidence of squamous cell carcinoma of the head and neck (SCCHN).
Serum level and genetic variants of GDF15 and substance use habits were evaluated in the risk
of SCCHN using a case-control study with 1191 hospital-based volunteers.
Serum GDF15 level showed an increasing trend among controls and SCCHN patients with
different cancer stages (Ptrend < 0.0001). Furthermore, it was positively correlated with lifetime
consumption of ABC in SCCHN patients (P < 0.05). An AA homozygote of rs1059369 showed
significant association with laryngeal cancer [odds ratio (OR) = 3.43] and had combined effects with
substance use of ABC in addictive interaction [synergy index (SI) = 1.54–2.85]. The CC
homozygotes of rs1054564 and rs1054221 were susceptible to SCCHN (ORs = 2.09 and 2.08,
respectively) and had combined effects with substance use of cigarette and betel quid in the risk of
SCCHN (SI = 1.39–2.01). The risk genotypes of both single-nucleotide polymorphisms were
significantly modified by cigarette smoking or betel chewing in oral cancer risk (SI =1.62–1.93) and
by alcohol drinking in laryngeal cancer risk (SI =3.84 and 3.85); however, no combined effect was
found in the risk of pharyngeal cancer.
GDF15 may influence the incidence and development of SCCHN by combined effects between
Introduction
Squamous cell carcinoma of the head and neck (SCCHN) is one of the most common malignancies
and causes high mortality and recurrence rates worldwide1. We have reported that substance use of
alcohol, betel quid and cigarette (ABC) reveals the great health risks in the Asian region2, and
substance use of ABC is the most common environmental risk factor of SCCHN3–5. Remarkably,
substance use of betel quid/areca nut is related to psychostimulant effects and sociocultural practices,
and it is widely prevalent in South Central Asia and the East Indies6. Globally, there are about 600
million betel quid chewers6; in Taiwan, there are approximately 2 million habitual chewers
(approximately 10% of the population)7. Numerous epidemiological studies also reported an increase
in the incidence of betel quid/areca nut-related oral cancer in Western countries due to the South
Asian and South Pacific immigrants, who have the habit of chewing betel quid/areca nut8–12.
Recently, we indicated that chewing tobacco-free betel quid in conjunction with consumption of
alcohol and/or tobacco impacts early cancer occurrence of the upper aerodigestive tract and
influences tumour site incidence pattern13. In addition, it has been reported that the 5-year survival
rate of SCCHN patients who chew betel is lower than that for those who do not chew betel14. The
substance use of betel quid is indeed a strong and independent risk factor of SCCHN in South
Central Asia and the East Indies.
The dose-dependent effects of ABC consumption are important indexes for evaluating the risks in
carcinogens are present at very low concentrations. Therefore, a number of constitutional factors may
underlie the susceptibility to SCCHN such as genetic predisposition. It has been suggested that an
intrinsic susceptibility to environmental genotoxic exposures attributes to carcinogenesis15 and that
the interaction between intrinsic susceptibility genes and environmental carcinogens can act in
concert to enhance cancer risk16. Several studies have reported a combined effect between
single-nucleotide polymorphisms (SNPs) and substance use of ABC, which results in an increased risk of
SCCHN17–21. Therefore, studying the combined effects between genes and environment can further
clarify on why there is a difference in the development of the onset and severity of SCCHN in
subjects with substance use habits of ABC.
Our previous study demonstrated that mRNA expression level of growth differentiation factor 15
(GDF15) can be upregulated on exposure to arecoline and areca nut extracts (ANEs) in human oral
cells17,22. GDF15 is a member of the transforming growth factor-β (TGFβ) superfamily, and its
biological function is known to regulate tissue differentiation and maintenance. Moreover, GDF15 is
showed to contribute to the tumour microenvironment by inhibiting tumour necrosis factor-α
secretion, which reduces the tumour-killing activity of macrophages23. Intriguingly, both GDF15 as
well as TGFβ act as a tumour suppressor in promoting apoptosis and against cell proliferation in
normal cells, and they act as a tumour promoter in enhancing the epithelial–mesenchymal
transformation, tumour invasion and angiogenesis in cancer cells24,25. Among betel-quid chewers,
pathogenesis of an oral precancerous condition26. In addition, the increased GDF15 expression level
in tissues of oral squamous cell carcinoma (OSCC) associates with higher malignant grade, and
knockdown of GDF15 expression in human tumour cell lines shows a significant decrease in cell
proliferation, colony formation and tumourigenicity27. Recently, it is reported that the novel
biological function of GDF15 in anti-apoptosis, by reduction of caspase-3/7 activity in OSCC cell
lines, may involve in OSCC development28. Because GDF15 potentially increases the risk of OSCC,
in this study, we further investigate whether GDF15 susceptibility genetic variants are associated
with SCCHN.
In the present study, we demonstrate the significant combined effects between GDF15 and substance
use of ABC in the risk of SCCHN. To our knowledge, neither the association between serum GDF15
level and lifetime consumption of ABC nor the combined effects between 3′-UTR SNPs of GDF15
gene and substance use habits of ABC have been investigated in SCCHN.
Methods and materials
Study population
A case-control study was conducted from three medical centres and included 474 male SCCHN
patients [331 oral cavity cancers (69.8%), 94 pharyngeal cancers (19.8%) and 49 laryngeal cancers
(10.3%)] and 717 male controls. This study was approved by the Human Experiment and Ethics
g of ethanol (equal to a 330-mL beer bottle containing 5% ethanol), one pack of betel quid was
defined as chewing 20 grains of betel quid and one pack of cigarettes was defined as smoking 20
cigarettes. Lifetime consumption of alcohol (drinks-years), betel quid (packs-years) and cigarette
(packs-years) was calculated and analysed.
Enzyme-linked immunosorbent assay (ELISA)
Serum specimens were obtained from 427 SCCHN patients (341 oral cancers, 53 pharyngeal cancers
and 33 laryngeal cancers) and 562 controls. The quantification of serum GDF15 level was
determined using human GDF15 ELISA development kit (DuoSet, R & D Systems), and each
sample was analysed in duplicate following the manufacturer’s protocol.
Gene re-sequencing and genotyping
Twenty pairs of SCCHN patients and controls were randomly assigned for identifying GDF15
sequence variants in the Taiwanese population. Genomic DNA was isolated using Gentra PureGene
Blood kit (Qiagen). Four amplicons, spanning −687 of 5′-UTR to +326 of 3′-UTR of the GDF15
gene, were re-sequenced using an ABI 3100 Genetic Analyzer (Life Technologies; also see
Supplementary table). Genetic variations and minor allele frequencies (MAFs) were determined and
analysed, and informative SNPs were further selected and genotyped.
Statistical analyses
(ANOVA) with the Bonferroni multiple comparisons test. The P value for trend was computed using
the slope estimate in a simple regression model. Relationships between serum GDF15 level and log
of ABC lifetime consumption were analysed using Pearson’s correlation. In the diagnostic accuracy
of serum GDF15 level and ABC substance use habits for SCCHN, receiver–operator characteristic
(ROC) curves were plotted by measuring area under curve (AUC). The goodness-of-fit χ2 test was
used to evaluate any deviation from Hardy–Weinberg equilibrium (HWE) in each of the selected
SNPs. Adjusted odds ratios (aORs), 95% confidence intervals (CIs) and exact P values were
calculated by logistic regression models controlling potential confounders. Combined effects
between genes and environment were evaluated using Rothman’s SI for additive interaction model29
and Khoury’s synergy index (SIM) for multiplicative interaction model30. All analyses were
executed using the SAS Statistical Package 9.1.3 (SAS Institute Inc.).
Results
Increased serum GDF15 levels in SCCHN patients
The serum GDF15 level in all SCCHN patients (1.22 ± 0.81 ng/mL), including those with oral (1.16
± 0.75 ng/mL), pharyngeal (1.61 ± 1.08 ng/mL) and laryngeal (1.27 ± 0.67 ng/mL) cancers, was
significantly higher than that of the controls (0.82 ± 0.63 ng/mL; P < 0.05; Figure 1a). A significant
increasing trend in the mean serum GDF15 level was detected in SCCHN patients at different cancer
stages (Ptrend = 0.049) and among controls and HN cancers at different cancer stages (Ptrend < 0.0001)
Correlation between serum GDF15 level and lifetime consumption of ABC
Regarding substance use of ABC, in the study population, the number of years of betel chewing (P =
0.005) and cigarette smoking (P < 0.01) was significantly higher in SCCHN patients than that in
controls, but there was no significant difference in alcohol intake (P = 0.061) (data not shown). The
findings found that an increased serum GDF15 level was significantly and positively correlated with
the lifetime consumption of A (P = 0.017), B (P = 0.012) and C (P = 0.010) in SCCHN patients
(Figure 2). However, such a relationship was insignificant in the controls.
Serum GDF15 level and ABC habits on diagnostic accuracy for SCCHN
The independent or combined effects of substance use habits, A, B, C, AB, AC, BC and ABC, for
SCCHN cancer risk were 3.1 (95% CI, 1.3–7.2), 53.2 (95% CI, 9.8–288.6), 3.0 (95% CI, 1.5–5.8),
62.0 (95% CI, 18.3–210.6), 9.8 (95% CI, 5.3–18.4), 51.7 (95% CI, 26.1–102.1) and 118.1 (95% CI,
65.2–213.8), respectively, after controlling age and ethnicity (data not shown). The results showed
that betel chewing is a strong environmental risk factor of SCCHN in Taiwan compared with alcohol
intake and cigarette smoking, as expected. The AUC of serum GDF15 level was 0.70 for SCCHN,
but the combined model with substance use habits of ABC improved the diagnostic performance to
0.90 (sensitivity = 83.4% and specificity = 83.3%; Figure 2d). The diagnostic performance of the
combined model was significantly greater than that of each independent factor (P < 0.0001; data not
SNP discovery and risk in SCCHN
In total, 19 SNPs were identified in the un-translated and coding regions of GDF15 gene from 20
pairs of patients and controls (Supplementary figure) by comparing the SNP database from
international populations at the HapMap Data Coordination Center (http://www.hapmap.org/); 4
SNPs were located at 5′-UTR (rs12459782, rs57573498, rs17526126 and rs77109188); 4 SNPs were
located at exon 1 (rs1059519, rs6413435, rs1059369 and rs16982331); 7 SNPs were located at exon
2 (rs1059022, rs1804826, rs3746195, rs45586234, rs45535632, rs1058587 and rs11556750) and 4
SNPs were located at 3′-UTR (rs1227733, rs1055150, rs1054564 and rs1054221). However, 8 of the
discovered SNPs (rs57573498, rs77109188, rs16982331, rs1059022, rs3746195, rs45535632,
rs45586234 and rs11556750) presented homozygotes in all subjects. Both rs6413435 and rs1059369
were neighbour SNPs (about 4 base pairs) in exon 1, and the information of rs6413435 was
arbitrarily replaced by rs1059369 in this study. Two SNPs, rs17526126 and rs1227733, failed in
probe design and synthesis (the Custom TaqMan SNP Genotyping Assays, ABI).
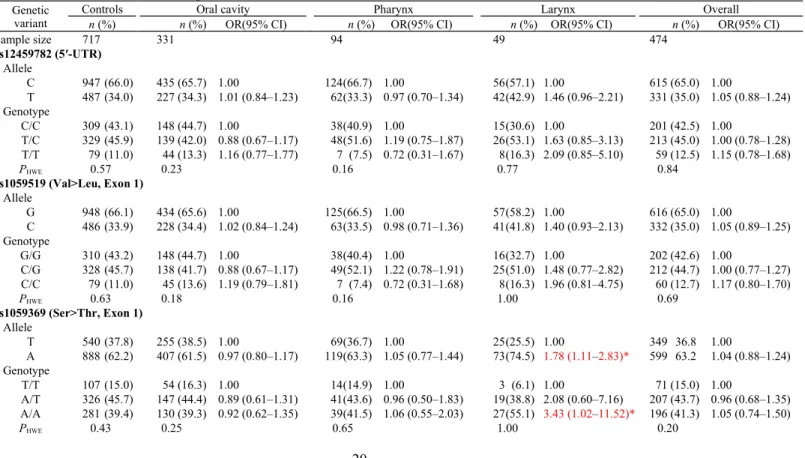
Finally, 8 selected SNPs, rs12459782 (T>C, 5′-UTR), rs1059519 (G>C, exon 1), rs1059369 (T>A,
exon 1), rs1804826 (G>T, exon 2), rs1058587 (C>G, exon 2), rs1055150 (G>C, 3′-UTR), rs1054564
(T>C, 3′-UTR) and rs1054221 (G>C, 3′-UTR), were genotyped, and the MAFs were 0.34 (T), 0.34
(C), 0.38 (T), 0.50 (T), 0.27 (G), 0.35 (C), 0.17 (C) and 0.17 (C), respectively. Each of the selected
in equilibrium in patients or controls (Table 1). Most SNPs located at the GDF15 promoter and
coding regions did not significantly associate with the subsite cancers or overall HN cancers, except
for the A allele and AA homozygote of rs1059369 (exon 1, T>A, Ser>Thr), which significantly
increased the risk of laryngeal cancer (OR = 1.78 and 3.43, respectively). Furthermore, CC
homozygotes of 3′-UTR SNPs, rs1054564 and rs1054221, showed 2.31- and 2.30-fold increases in
risk, respectively, for overall HN cancer mainly owing to oral cavity cancer (OR = 2.20 and 2.18,
respectively) and laryngeal cancer (OR = 5.28 and 5.26, respectively). No genetic effect of GDF15
was found in the risk of pharyngeal cancer.
Gene–environment interaction in the risk of SCCHN
Because rs1059369 (exon 1), rs1054564 (3′-UTR) and rs1054221 (3′-UTR) are significantly
associated with laryngeal and oral cavity cancer, we further evaluated the gene–environment
interactions between these genetic variants and substance use habits of ABC in risk of HN cancers.
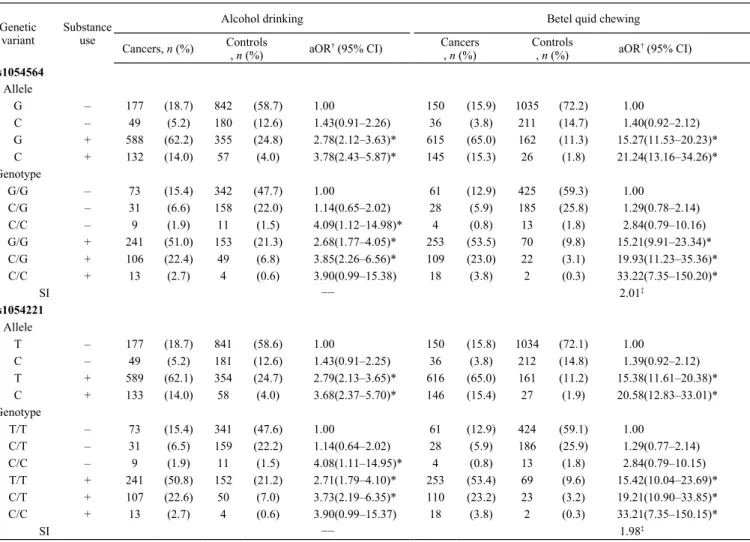
In Table 2, as shown, combined effects between substance use habits of ABC and the A allele/AA
homozygote of rs1059369 were found to increase the risk of laryngeal cancer in addictive interaction
models (SI = 1.26–2.25 and 1.54–2.85, respectively). Remarkably, the A allele/AA homozygote of
rs1059369 was strongly modified by exposure to betel quid chewing in multiplicative interaction
models in the risk of laryngeal cancer (SIM = 2.25 and 2.85, respectively). Moreover, we also found
that the susceptibility genotypes (CC) of 3′-UTR SNPs, rs1054564 and rs1054221, modified the
1.39, respectively) based on an additive interaction model in the risk of overall HN cancer (Table 3).
In subsite cancers of SCCHN, as shown in Table 4, the CC homozygotes of both 3′-UTR SNPs
addictively modified the effects of betel quid chewing (SI = 1.93 and 1.89, respectively) or cigarette
smoking (SI = 1.63 and 1.62, respectively) in oral cancer. Intriguingly, in laryngeal cancer, the CC
homozygotes of both 3′-UTR SNPs also modified the effects of alcohol consumption based on
addictive interaction models (SI = 3.84 and 3.85, respectively) or multiplicative interaction models
(SIM = 1.59 and 1.59, respectively). Nevertheless, the susceptibility genotypes of rs1059369,
rs1054564 and rs1054221 were not modified by exposure to substance use of ABC in pharyngeal
cancer.
Discussion
In Taiwan, the incidence and mortality of SCCHN has increased over the past two decades. We
recently found that 42%–45% of Taiwanese betel quid chewers are dependent on betel quid, and the
adjusted population attributable risk proportion of tobacco-free betel quid chewing accounted for
78.7%, 66.1% and 17.8% of the patients with oral, pharyngeal and laryngeal cancers,
respectively31,32. Alcohol drinking and cigarette smoking are concomitant habits with betel quid
chewing in eastern and South Asian communities2. Subjects with long-term or high frequency of
ABC consumption are at a high risk of developing SCCHN. In the present study, we found that
serum GDF15 level was positively correlated with total life consumption of ABC in SCCHN
substance abuse. GDF15 may act as a tumour suppressor or tumour promoter in epithelial
carcinogenesis, because it is a divergent member of the TGFβ superfamily, which inhibits cell
proliferation and enhances apoptosis in normal cells, whereas it enhances epithelial–mesenchymal
transformation, angiogenesis and tumour invasion in cancer cells24. We showed that areca nut, the
main component of betel quid, upregulates GDF15 gene expression in human oral tumour cells17;
however, no report has shown the effect of alcohol or cigarette on modulating GDF15 expression in
oral models. Studying the effects of ABC in inhibiting GDF15 regular expression in normal status
and inducing GDF15 overexpression in tumour initiation may provide important evidence in the
early development of SCCHN; the pathogenetic mechanisms also remain to be defined.
In this study, significantly increased serum levels of GDF15 were observed in SCCHN as well as in
some human cancers33–35. In addition, the increased levels were significantly associated with clinical
signs of malignancy in SCCHN patients. In OSCC tissues, the overexpression of GDF15 positively
correlated with histopathological malignant grade27. It appears that the dimeric mature form of
GDF15 is largely secreted and cleaved from overexpressed intracellular dimeric precursor in
malignant tumours, and subsequently released into blood circulation in OSCC patients. At the
subsites of SCCHN, not only oral cavity cancer but also pharyngeal and laryngeal cancer showed an
abnormally increased serum level of GDF15. Because elevated serum GDF15 level is positively
correlated with substance use of ABC, we further performed a combined model of ROC curve to
environmental risk factors. This model could provide a detection strategy for screening individual
susceptibility in the risk of early onset of SCCHN from high-risk populations.
To explore whether individual genetic variants of the GDF15 gene are associated with SCCHN, we
further investigated the AA homozygotes of rs1059369 and CC homozygotes of two 3′-UTR SNPs
(rs1054564 and rs1054221), particularly at the subsite cancers of oral cavity and larynx. As per our
findings, however, no genetic effect of GDF15 was found in the risk of pharyngeal cancer. Because
an elevated serum level of GDF15 was observed in this cancer site, we supposed that other signalling
pathways of GDF15 may be involved in the development of pharyngeal cancer such as
overexpression or overactivity of its upstream transcription factors. rs1059369 is a missense SNP
(TCC>ACC) located at exon 1 that has an amino acid residue change at position 48 from Ser to Thr
in the GDF15 propeptide, and it is susceptible in laryngeal cancer. However, in prostate cancer,
rs1059369 did not show a significant association with cancer risk and survival rate of prostate
cancer36,37. Whether the physico-chemical property change of rs1059369 from small size and polar
(Ser) to medium size and polar (Thr) influences the expression of GDF15 pro-form requires further
investigation.
It is notable that the 3′-UTR SNP, rs1054564, but not the other three natural non-synonymous SNPs,
rs1059519 (exon1, Val/Leu, position 9), rs1059369 and rs1058587 (exon2, His/Asp, position 202),
Moreover, a differentially expressed transcript tag ‘GTGCTCATTC’ near sequence positions of
rs1054564 and rs1054221 exhibited a >20-fold increase in the risk of colorectal cancer38. Although
the H6D polymorphism (C>G) is a potential tag SNP for studying cancer risk and survival in prostate
cancer36,39, in the present study, it did not present a significant susceptibility to SCCHN patients.
These findings indicate that genetic variants located at the 3′-UTR region of the GDF15 gene may
modulate its endogenous expression and contribute in the development of some human cancers. To
our knowledge, in the TargetScan database for human miRNAs, the putative mir-1233 and
hsa-mir-1225-3p could be easily targeted to the complementary sequence at wild type (G) of rs1054564,
and hsa-mir-410 could be targeted to the sequence at wild type (T) of rs105422. However, further
studies are needed to validate these genotype-specific putative miRNAs on modulating endogenous
GDF15 expression in the development of SCCHN.
The substance use habits and frequencies of ABC have known to cause the different risks in the
subsite cancers of SCCHN. Nevertheless, all individuals exposed to the same type and consumption
of ABC do not develop SCCHN or even the specific subsite cancers. It is now understood that cancer
development is not only due to exogenous carcinogens but also due to their interactions with
individual susceptibility genes that are involved in tumour initiation, progression or metastasis. In
molecular epidemiological approaches, environmental risk factors and genetic predisposition can, in
combination, additively or multiplicatively confer a range of susceptibilities in risk assessment
(AA), rs1054564 (CC) and rs1054221 (CC) present higher risks of HN cancers, especially when
combined with the habits of alcohol drinking, betel quid chewing or cigarette smoking. Nevertheless,
the precise role of GDF15 in ABC-related SCCHN is not clear at present. Additional studies with the
effect of ABC on changes in cellular microRNAs, which actually modulate the expression of GDF15
or its regulatory transcription factors40–45, are needed for elucidating the molecular mechanisms in
carcinogenesis of ABC-related SCCHN.
Together, the serum GDF15 level can serve as a clinical marker in SCCHN for replacing the
determination in cancerous tissues. The present study is the first report to explore the relationships
and interactions between GDF15 and substance use of ABC in the risk of SCCHN. It is the combined
effects of these inherent SNPs in the GDF15 gene and substance use of ABC that interpret whether
an individual develops early onset of HN cancers. Although we do not know the complete functional
significance of these identified variants, we suggest that they may contribute to ABC-related SCCHN
susceptibility.
Acknowledgements
This work was supported by grants from the Nation Science Council (NSC-100-2314-B-037-037 and
NSC-101-2314-B-039-045-MY3) and Center of Excellence for Environmental Medicine of
References
1. Ferlay J, Bray F, Pisani P, Parkin DM. GLOBOCAN 2002: cancer incidence, mortality and prevalence worldwide. IARC CancerBase No 5, version 20. Lyon: IARC Press; 2004.
2. Lee CH, Ko AM, Warnakulasuriya S, Yin BL, Sunarjo, Zain RB, et al. Intercountry prevalences and practices of betel-quid use in south, southeast and eastern Asia regions and associated oral preneoplastic disorders: an international collaborative study by Asian betel-quid consortium of south and east Asia. Int J Cancer. 2011 Oct;129(7):1741-51.
3. Ko YC, Huang YL, Lee CH, Chen MJ, Lin LM, Tsai CC. Betel quid chewing, cigarette smoking and alcohol consumption related to oral cancer in Taiwan. J Oral Pathol Med. 1995 Nov;24(10):450-3.
4. Lee KW, Kuo WR, Tsai SM, Wu DC, Wang WM, Fang FM, et al. Different impact from betel quid, alcohol and cigarette: risk factors for pharyngeal and laryngeal cancer. Int J Cancer. 2005 Dec;117(5):831-6.
5. Hashibe M, Brennan P, Benhamou S, Castellsague X, Chen C, Curado MP, et al. Alcohol drinking in never users of tobacco, cigarette smoking in never drinkers, and the risk of head and neck cancer: pooled analysis in the International Head and Neck Cancer Epidemiology Consortium. J Natl Cancer Inst. 2007 May;99(10):777-89.
6. Gupta PC, Warnakulasuriya S. Global epidemiology of areca nut usage. Addict Biol. 2002 Jan;7(1):77-83.
7. Ko YC, Chiang TA, Chang SJ, Hsieh SF. Prevalence of betel quid chewing habit in Taiwan and related sociodemographic factors. J Oral Pathol Med. 1992 Jul;21(6):261-4.
8. Warnakulasuriya S. Areca nut use following migration and its consequences. Addict Biol. 2002 Jan;7(1):127-32.
9. Auluck A, Hislop G, Poh C, Zhang L, Rosin MP. Areca nut and betel quid chewing among South Asian immigrants to Western countries and its implications for oral cancer screening. Rural and remote health. 2009 Apr-Jun;9(2):1118.
10. Changrani J, Gany F. Paan and Gutka in the United States: an emerging threat. Journal of immigrant health. 2005 Apr;7(2):103-8.
11. Moles DR, Fedele S, Speight PM, Porter SR, dos Santos Silva I. Oral and pharyngeal cancer in South Asians and non-South Asians in relation to socioeconomic deprivation in South East England. Br J Cancer. 2008 Feb;98(3):633-5.
12. Warnakulasuriya KA, Johnson NW, Linklater KM, Bell J. Cancer of mouth, pharynx and nasopharynx in Asian and Chinese immigrants resident in Thames regions. Oral Oncol. Sep 1999;35(5):471-475.
13. Lee CH, Lee KW, Fang FM, Wu DC, Shieh TY, Huang HL, et al. The use of tobacco-free betel-quid in conjunction with alcohol/tobacco impacts early-onset age and carcinoma distribution for upper aerodigestive tract cancer. J Oral Pathol Med. 2011 Mar;doi: 10.1111/j.1600-0714.2011.01022.x
14. Lo WL, Kao SY, Chi LY, Wong YK, Chang RC. Outcomes of oral squamous cell carcinoma in Taiwan after surgical therapy: factors affecting survival. J Oral Maxillofac Surg. 2003 Jul;61(7):751-8.
15. Li FP, Montesano R. Interactions of cancer susceptibility genes and environmental carcinogens. American Association for Cancer Research (AACR)-International Agency for Research on Cancer (IARC) Joint Conference. Cancer Res. 1994 Aug;54(15):4243-7.
16. Hunter DJ. Gene-environment interactions in human diseases. Nat Rev Genet. 2005 Apr;6(4):287-98.
17. Chiang SL, Chen PH, Lee CH, Ko AM, Lee KW, Lin YC, et al. Up-regulation of inflammatory signalings by areca nut extract and role of cyclooxygenase-2 -1195G>a polymorphism reveal risk of oral cancer. Cancer Res. 2008 Oct;68(20):8489-98.
18. Chen PH, Lee KW, Chen CH, Shieh TY, Ho PS, Wang SJ, et al. CYP26B1 is a novel candidate gene for betel quid-related oral squamous cell carcinoma. Oral Oncol. 2011 Jul;47(7):594-600.
19. Sturgis EM, Castillo EJ, Li L, Zheng R, Eicher SA, Clayman GL, et al. Polymorphisms of DNA repair gene XRCC1 in squamous cell carcinoma of the head and neck. Carcinogenesis. 1999 Nov;20(11):2125-9.
20. Kietthubthew S, Sriplung H, Au WW. Genetic and environmental interactions on oral cancer in Southern Thailand. Environ Mol Mutagen. 2001 Mar;37(2):111-6.
21. Ihsan R, Devi TR, Yadav DS, Mishra AK, Sharma J, Zomawia E, et al. Investigation on the role of p53 codon 72 polymorphism and interactions with tobacco, betel quid, and alcohol in susceptibility to cancers in a high-risk population from North East India. DNA Cell Biol. 2011 Mar;30(3):163-71.
22. Chiang SL, Jiang SS, Wang YJ, Chiang HC, Chen PH, Tu HP, et al. Characterization of arecoline-induced effects on cytotoxicity in normal human gingival fibroblasts by global gene expression profiling. Toxicol Sci. 2007 Nov;100(1):66-74.
23. Bootcov MR, Bauskin AR, Valenzuela SM, Moore AG, Bansal M, He XY, et al. MIC-1, a novel macrophage inhibitory cytokine, is a divergent member of the TGF-beta superfamily. Proc Natl Acad Sci U S A. 1997 Oct;94(21):11514-9.
24. Dumont N, Arteaga CL. Targeting the TGF beta signaling network in human neoplasia. Cancer Cell. 2003 Jun;3(6):531-6.
25. Eling TE, Baek SJ, Shim M, Lee CH. NSAID activated gene (NAG-1), a modulator of tumorigenesis. J Biochem Mol Biol. 2006 Nov;39(6):649-55.
26. Rajalalitha P, Vali S. Molecular pathogenesis of oral submucous fibrosis--a collagen metabolic disorder. J Oral Pathol Med. 2005 Jul;34(6):321-8.
27. Zhang L, Yang X, Pan HY, Zhou XJ, Li J, Chen WT, et al. Expression of growth differentiation factor 15 is positively correlated with histopathological malignant grade and in vitro cell proliferation in oral squamous cell carcinoma. Oral Oncol. 2009 Jul;45(7):627-32.
28. Schiegnitz E, Kammerer PW, Koch FP, Kruger M, Berres M, Al-Nawas B. GDF 15 as an anti-apoptotic, diagnostic and prognostic marker in oral squamous cell carcinoma. Oral Oncol. 2012 Jul;48(7):608-14.
29. Rothman KJ. Synergy and antagonism in cause-effect relationships. Am J Epidemiol. 1974 Jun;99(6):385-8.
30. Khoury MJ, Flanders WD. Nontraditional epidemiologic approaches in the analysis of gene-environment interaction: case-control studies with no controls! Am J Epidemiol. 1996 Aug;144(3):207-13.
31. Lee CH, Ko AM, Yen CF, Chu KS, Gao YJ, Warnakulasuriya S, et al. Betel-quid dependence and oral potentially malignant disorders in six Asian countries. Br J Psychiatry. 2012 Nov;201(5):383-91.
32. Lee CH, Lee KW, Fang FM, Wu DC, Tsai SM, Chen PH, et al. The neoplastic impact of tobacco-free betel-quid on the histological type and the anatomical site of aerodigestive tract cancers. Int J Cancer. 2012 Sep;131(5):E733-43.
33. Brown DA, Ward RL, Buckhaults P, Liu T, Romans KE, Hawkins NJ, et al. MIC-1 serum level and genotype: associations with progress and prognosis of colorectal carcinoma. Clin Cancer Res. 2003 Jul;9(7):2642-50.
34. Koopmann J, Buckhaults P, Brown DA, Zahurak ML, Sato N, Fukushima N, et al. Serum macrophage inhibitory cytokine 1 as a marker of pancreatic and other periampullary cancers. Clin Cancer Res. 2004 Apr;10(7):2386-92.
35. Brown DA, Lindmark F, Stattin P, Balter K, Adami HO, Zheng SL, et al. Macrophage inhibitory cytokine 1: a new prognostic marker in prostate cancer. Clin Cancer Res. 2009 Nov;15(21):6658-64.
36. Lindmark F, Zheng SL, Wiklund F, Bensen J, Balter KA, Chang B, et al. H6D polymorphism in macrophage-inhibitory cytokine-1 gene associated with prostate cancer. J Natl Cancer Inst. 2004 Aug;96(16):1248-54.
37. Cheng I, Krumroy LM, Plummer SJ, Casey G, Witte JS. MIC1 and IL1RN genetic variation and advanced prostate cancer risk. Cancer Epidemiol Biomarkers Prev. 2007 Jun;16(6):1309-11.
38. Buckhaults P, Rago C, St Croix B, Romans KE, Saha S, Zhang L, et al. Secreted and cell surface genes expressed in benign and malignant colorectal tumors. Cancer Res. 2001 Oct;61(19):6996-7001.
39. Hayes VM, Severi G, Southey MC, et al. Macrophage inhibitory cytokine-1 H6D polymorphism, prostate cancer risk, and survival. Cancer Epidemiol Biomarkers Prev. 2006 Jun;15(6):1223-1225.
40. Wollmann W, Goodman ML, Bhat-Nakshatri P, Kishimoto H, Goulet RJ, Jr., Mehrotra S, et al. The macrophage inhibitory cytokine integrates AKT/PKB and MAP kinase signaling pathways in breast cancer cells. Carcinogenesis. 2005 May;26(5):900-7.
41. Li PX, Wong J, Ayed A, Ngo D, Brade AM, Arrowsmith C, et al. Placental transforming growth factor-beta is a downstream mediator of the growth arrest and apoptotic response of tumor cells to DNA damage and p53 overexpression. J Biol Chem. 2000 Jun;275(26):20127-35.
42. Wang A, Arantes S, Conti C, McArthur M, Aldaz CM, MacLeod MC. Epidermal hyperplasia and oral carcinoma in mice overexpressing the transcription factor ATF3 in basal epithelial cells. Mol Carcinog. 2007 Jun;46(6):476-87.
43. Shim M, Eling TE. Protein kinase C-dependent regulation of NAG-1/placental bone morphogenic protein/MIC-1 expression in LNCaP prostate carcinoma cells. J Biol Chem. 2005 May;280(19):18636-42.
44. Baek SJ, Kim JS, Moore SM, Lee SH, Martinez J, Eling TE. Cyclooxygenase inhibitors induce the expression of the tumor suppressor gene EGR-1, which results in the up-regulation of NAG-1, an antitumorigenic protein. Mol Pharmacol. 2005 Feb;67(2):356-64.
45. Baek SJ, Horowitz JM, Eling TE. Molecular cloning and characterization of human
nonsteroidal anti-inflammatory drug-activated gene promoter. Basal transcription is mediated by Sp1 and Sp3. J Biol Chem. 2001 Sep;276(36):33384-92.
Figure 1: Serum GDF15 level in HN cancer. (a) Serum GDF15 levels (ng/mL) in overall and subsites of HN cancer were compared with controls using Student’s t-test or ANOVA with Bonferroni multiple comparisons test. (b) P values for trend (Ptrend) of the serum GDF15
levels were estimated among controls and SCCHN patients with different cancer stages using simple regression models. *P < 0.001, **P < 0.0001.
Figure 2: Linear relationship and ROC curve between ABC substance use and serum GDF15 level in SCCHN. The plots illustrate correlations between the lifetime consumption of (a) alcoholic
beverages (years-drinks), (b) betel quid (years-packs) and (c) cigarettes (years-packs) and serum
GDF15 level in SCCHN patients and controls. R-squared value (R-sq) and P value were calculated
using Pearson’s correlation analysis. (d) Area under ROC curve was measured to evaluate the
predictive models from substance use habits of ABC and serum GDF15 level in SCCHN
development.
Table 1 Genetic variants of the GDF15 gene in the risk of HN cancers
Genetic variant
Controls Oral cavity Pharynx Larynx Overall
n (%) n (%) OR(95% CI) n (%) OR(95% CI) n (%) OR(95% CI) n (%) OR(95% CI)
Sample size 717 331 94 49 474 rs12459782 (5′-UTR) Allele C 947 (66.0) 435 (65.7) 1.00 124(66.7) 1.00 56(57.1) 1.00 615 (65.0) 1.00 T 487 (34.0) 227 (34.3) 1.01 (0.84–1.23) 62(33.3) 0.97 (0.70–1.34) 42(42.9) 1.46 (0.96–2.21) 331 (35.0) 1.05 (0.88–1.24) Genotype C/C 309 (43.1) 148 (44.7) 1.00 38(40.9) 1.00 15(30.6) 1.00 201 (42.5) 1.00 T/C 329 (45.9) 139 (42.0) 0.88 (0.67–1.17) 48(51.6) 1.19 (0.75–1.87) 26(53.1) 1.63 (0.85–3.13) 213 (45.0) 1.00 (0.78–1.28) T/T 79 (11.0) 44 (13.3) 1.16 (0.77–1.77) 7 (7.5) 0.72 (0.31–1.67) 8(16.3) 2.09 (0.85–5.10) 59 (12.5) 1.15 (0.78–1.68) PHWE 0.57 0.23 0.16 0.77 0.84 rs1059519 (Val>Leu, Exon 1) Allele G 948 (66.1) 434 (65.6) 1.00 125(66.5) 1.00 57(58.2) 1.00 616 (65.0) 1.00 C 486 (33.9) 228 (34.4) 1.02 (0.84–1.24) 63(33.5) 0.98 (0.71–1.36) 41(41.8) 1.40 (0.93–2.13) 332 (35.0) 1.05 (0.89–1.25) Genotype G/G 310 (43.2) 148 (44.7) 1.00 38(40.4) 1.00 16(32.7) 1.00 202 (42.6) 1.00 C/G 328 (45.7) 138 (41.7) 0.88 (0.67–1.17) 49(52.1) 1.22 (0.78–1.91) 25(51.0) 1.48 (0.77–2.82) 212 (44.7) 1.00 (0.77–1.27) C/C 79 (11.0) 45 (13.6) 1.19 (0.79–1.81) 7 (7.4) 0.72 (0.31–1.68) 8(16.3) 1.96 (0.81–4.75) 60 (12.7) 1.17 (0.80–1.70) PHWE 0.63 0.18 0.16 1.00 0.69 rs1059369 (Ser>Thr, Exon 1) Allele T 540 (37.8) 255 (38.5) 1.00 69(36.7) 1.00 25(25.5) 1.00 349 36.8 1.00 A 888 (62.2) 407 (61.5) 0.97 (0.80–1.17) 119(63.3) 1.05 (0.77–1.44) 73(74.5) 1.78 (1.11–2.83)* 599 63.2 1.04 (0.88–1.24) Genotype T/T 107 (15.0) 54 (16.3) 1.00 14(14.9) 1.00 3 (6.1) 1.00 71 (15.0) 1.00 A/T 326 (45.7) 147 (44.4) 0.89 (0.61–1.31) 41(43.6) 0.96 (0.50–1.83) 19(38.8) 2.08 (0.60–7.16) 207 (43.7) 0.96 (0.68–1.35) A/A 281 (39.4) 130 (39.3) 0.92 (0.62–1.35) 39(41.5) 1.06 (0.55–2.03) 27(55.1) 3.43 (1.02–11.52)* 196 (41.3) 1.05 (0.74–1.50) PHWE 0.43 0.25 0.65 1.00 0.20
rs1804826 (Synonymous, Exon 2) Allele G 715 (50.4) 311 (47.6) 1.00 97(52.7) 1.00 56(57.1) 1.00 464 (49.6) 1.00 T 705 (49.6) 343 (52.4) 1.12 (0.93–1.35) 87(47.3) 0.91 (0.67–1.24) 42(42.9) 0.76 (0.50–1.15) 472 (50.4) 1.00 (0.85–1.18) Genotype G/G 98 (13.8) 30 (9.2) 1.00 14(15.2) 1.00 10(20.4) 1.00 54 (11.5) 1.00 T/G 519 (73.1) 251 (76.8) 1.50 (0.97–2.32) 69(75.0) 0.88 (0.48–1.63) 36(73.5) 0.65 (0.31–1.35) 356 (76.1) 1.18 (0.82–1.70) T/T 93 (13.1) 46 (14.1) 1.46 (0.85–2.50) 9 (9.8) 0.61 (0.25–1.48) 3 (6.1) 0.29 (0.08–1.08) 58 (12.4) 1.02 (0.64–1.63) PHWE <0.01 <0.01 <0.01 <0.01 <0.01 rs1058587 (Asp>His, Exon 2) Allele G 398 (28.0) 170 (26.2) 1.00 54(29.0) 1.00 31(31.6) 1.00 255 (27.3) 1.00 C 1022 (72.0) 480 (73.8) 1.10 (0.89–1.36) 132(71.0) 0.95 (0.68–1.33) 67(68.4) 0.84 (0.54–1.31) 679 (72.7) 1.04 (0.86–1.25) Genotype G/G 52 (7.3) 22 (6.8) 1.00 9 (9.7) 1.00 6(12.2) 1.00 37 (7.9) 1.00 C/G 294 (41.4) 126 (38.8) 1.01 (0.59–1.74) 36(38.7) 0.71 (0.32–1.56) 19(38.8) 0.56 (0.21–1.47) 181 (38.8) 0.87 (0.55–1.37) C/C 364 (51.3) 177 (54.5) 1.15 (0.68–1.95) 48(51.6) 0.76 (0.35–1.64) 24(49.0) 0.57 (0.22–1.46) 249 (53.3) 0.96 (0.61–1.51) PHWE 0.51 1.00 0.61 0.52 0.63 rs1055150 (3′-UTR) Allele G 946 (66.0) 428 (64.7) 1.00 119(63.3) 1.00 55(56.1) 1.00 602 (63.5) 1.00 C 488 (34.0) 234 (35.3) 1.06 (0.87–1.29) 69(36.7) 1.12 (0.82–1.54) 43(43.9) 1.52 (0.99–2.29) 346 (36.5) 1.11 (0.94–1.32) Genotype G/G 309 (43.1) 142 (42.9) 1.00 32(34.0) 1.00 14(28.6) 1.00 188 (39.6) 1.00 C/G 328 (45.7) 144 (43.5) 0.96 (0.72–1.26) 55(58.5) 1.62 (1.02–2.57)* 27(55.1) 1.82 (0.94–3.53) 226 (47.7) 1.13 (0.88–1.45) C/C 80 (11.2) 45 (13.6) 1.22 (0.81–1.86) 7 (7.4) 0.85 (0.36–1.99) 8(16.3) 2.21 (0.90–5.44) 60 (12.5) 1.23 (0.84–1.80) PHWE 0.67 0.40 0.01 0.56 0.62 rs1054564 (3′-UTR) Allele G 1197 (83.5) 541 (81.7) 1.00 154(82.8) 1.00 70(71.4) 1.00 765 (80.9) 1.00 C 237 (16.5) 121 (18.3) 1.13 (0.89–1.44) 32(17.2) 1.05 (0.70–1.57) 28(28.6) 2.02 (1.28–3.22)* 181 (19.1) 1.20 (0.97–1.48) Genotype G/G 495 (69.0) 225 (68.0) 1.00 64(68.8) 1.00 25(51.0) 1.00 314 (66.4) 1.00 C/G 207 (28.9) 91 (27.5) 0.97 (0.72–1.30) 26(28.0) 0.97 (0.60–1.58) 20(40.8) 1.91 (1.04–3.52)* 137 (29.0) 1.04 (0.81–1.35) C/C 15 (2.1) 15 (4.5) 2.20 (1.06–4.58)* 3 (3.2) 1.55 (0.44–5.50) 4 (8.2) 5.28 (1.63–17.08)* 22 (4.6) 2.31 (1.18–4.52)* PHWE 0.28 0.14 0.73 1.00 0.19 rs1054221 (3′-UTR) Allele T 1195 (83.3) 542 (81.9) 1.00 154(81.9) 1.00 70(71.4) 1.00 766 (80.8) 1.00 C 239 (16.7) 120 (18.1) 1.11 (0.87–1.41) 34(18.1) 1.10 (0.74–1.64) 28(28.6) 2.00 (1.26–3.17)* 182 (19.2) 1.19 (0.96–1.47) Genotype T/T 493 (68.8) 226 (68.3) 1.00 63(67.0) 1.00 25(51.0) 1.00 314 (66.2) 1.00 C/T 209 (29.1) 90 (27.2) 0.94 (0.70–1.26) 28(29.8) 1.05 (0.65–1.68) 20(40.8) 1.89 (1.03–3.47)* 138 (29.2) 1.04 (0.80–1.34) C/C 15 (2.1) 15 (4.5) 2.18 (1.05–4.54)* 3 (3.2) 1.57 (0.44–5.56) 4 (8.2) 5.26 (1.63–17.01)* 22 (4.6) 2.30 (1.18–4.51)* PHWE 0.23 0.13 1.00 1.00 0.18
PHWE, exact P values for HWE were calculated using 10,000 permutations. *P < 0.05.
Table 2 Combined aORs of laryngeal cancer with regard to rs1059369 (Ser>Thr) and substance use habits of ABC
Genetic variant Substance use
Alcohol drinking Betel quid chewing Cigarette smoking
Cases/Controls aOR*(95% CI) Cases/Contr
ols aOR(95% CI) Cases/Controls aOR(95% CI)
Allele T – 5/380 1.00 11/462 1.0 2/298 1.00 A – 25/638 2.80 (1.00–7.84) 29/778 1.7 (0.80–3.59) 8/464 2.73 (0.56–13.23) T + 20/160 4.58 (1.57–13.40)* 14/78 6.4 (2.51–16.41)* 23/242 4.95 (1.10–22.29)* A + 48/250 7.77 (2.85–21.15)* 44/110 14.7 (6.58–32.85)* 65/424 9.30 (2.17–39.91)* SI/SIM 1.26† 2.25†/1.35‡ 1.46† Genotype T/T + A/T – 5/306 1.00 10/373 1.0 2/238 1.00 A/A – 10/203 2.65 (0.81–8.72) 10/247 1.8 (0.68–4.63) 3/143 2.80 (0.45–17.60) T/T + A/T + 17/127 3.79 (1.24–11.63)* 12/60 6.1 (2.14–17.62)* 20/195 4.24 (0.90–19.91)
SI/SIM 1.54‡ 2.85†/1.62‡ 1.63†
*Odd ratios were adjusted with age, ethnicity and another two substance use. *P < 0.05
†Synergy index (SI) was evaluated for additive interaction model by Rothman’s synergy index.
‡Synergy index multiplicative (SIM) was evaluated for multiplicative interaction model by Khoury’s synergy index.
Table 3 Combined aORs of overall SCCHN with regard to GDF15 3′-UTR genetic variants and substance use of ABC Genetic
variant Substanceuse
Alcohol drinking Betel quid chewing
Cancers, n (%) Controls, n (%) aOR† (95% CI) Cancers
, n (%) Controls, n (%) aOR† (95% CI)
rs1054564 Allele G – 177 (18.7) 842 (58.7) 1.00 150 (15.9) 1035 (72.2) 1.00 C – 49 (5.2) 180 (12.6) 1.43(0.91–2.26) 36 (3.8) 211 (14.7) 1.40(0.92–2.12) G + 588 (62.2) 355 (24.8) 2.78(2.12–3.63)* 615 (65.0) 162 (11.3) 15.27(11.53–20.23)* C + 132 (14.0) 57 (4.0) 3.78(2.43–5.87)* 145 (15.3) 26 (1.8) 21.24(13.16–34.26)* Genotype G/G – 73 (15.4) 342 (47.7) 1.00 61 (12.9) 425 (59.3) 1.00 C/G – 31 (6.6) 158 (22.0) 1.14(0.65–2.02) 28 (5.9) 185 (25.8) 1.29(0.78–2.14) C/C – 9 (1.9) 11 (1.5) 4.09(1.12–14.98)* 4 (0.8) 13 (1.8) 2.84(0.79–10.16) G/G + 241 (51.0) 153 (21.3) 2.68(1.77–4.05)* 253 (53.5) 70 (9.8) 15.21(9.91–23.34)* C/G + 106 (22.4) 49 (6.8) 3.85(2.26–6.56)* 109 (23.0) 22 (3.1) 19.93(11.23–35.36)* C/C + 13 (2.7) 4 (0.6) 3.90(0.99–15.38) 18 (3.8) 2 (0.3) 33.22(7.35–150.20)* SI −− 2.01‡ rs1054221 Allele T – 177 (18.7) 841 (58.6) 1.00 150 (15.8) 1034 (72.1) 1.00 C – 49 (5.2) 181 (12.6) 1.43(0.91–2.25) 36 (3.8) 212 (14.8) 1.39(0.92–2.12) T + 589 (62.1) 354 (24.7) 2.79(2.13–3.65)* 616 (65.0) 161 (11.2) 15.38(11.61–20.38)* C + 133 (14.0) 58 (4.0) 3.68(2.37–5.70)* 146 (15.4) 27 (1.9) 20.58(12.83–33.01)* Genotype T/T – 73 (15.4) 341 (47.6) 1.00 61 (12.9) 424 (59.1) 1.00 C/T – 31 (6.5) 159 (22.2) 1.14(0.64–2.02) 28 (5.9) 186 (25.9) 1.29(0.77–2.14) C/C – 9 (1.9) 11 (1.5) 4.08(1.11–14.95)* 4 (0.8) 13 (1.8) 2.84(0.79–10.15) T/T + 241 (50.8) 152 (21.2) 2.71(1.79–4.10)* 253 (53.4) 69 (9.6) 15.42(10.04–23.69)* C/T + 107 (22.6) 50 (7.0) 3.73(2.19–6.35)* 110 (23.2) 23 (3.2) 19.21(10.90–33.85)* C/C + 13 (2.7) 4 (0.6) 3.90(0.99–15.37) 18 (3.8) 2 (0.3) 33.21(7.35–150.15)* SI −− 1.98‡
†Odd ratios were adjusted with age, ethnicity and another two substance use. *P < 0.05 ‡Synergy index (SI) was evaluated for additive interaction model by Rothman’s synergy index.
Table 4 Combined aORs with regard to GDF15 3′-UTR genetic variants and substance use of ABC on the subsites of SCCHN
Genetic variant
Substance use
Alcohol drinking Betel-quid chewing Cigarette smoking
Cases/Con
trols aOR† (95% CI)
Cases/Cont
rols aOR† (95% CI)
Cases/Cont
rols aOR† (95% CI)
Oral cancer rs1054564
G/G – 57/342 1.00 34/425 1.00 20/255 1.00
C/C – 7/11 5.30 (1.35–20.87)* 2/13 2.53 (0.51–12.46) 1/10 2.21 (0.26–18.50) G/G + 168/153 2.08 (1.30–3.32)* 191/70 20.75 (12.68–33.94)* 205/240 2.27 (1.25–4.11)* C/G + 69/49 2.52 (1.37–4.62)* 74/22 23.42 (12.46–44.03)* 82/89 3.01 (1.53–5.91)* C/C + 8/4 1.74 (0.41–7.31) 13/2 42.02 (8.92–198.04)* 14/5 5.04 (1.24–20.43)* SI‡ — 1.93‡ 1.63‡ rs1054221 T/T – 57/341 1.00 34/424 1.00 20/254 1.00 C/T – 22/159 1.26 (0.66–2.43) 17/186 1.36 (0.73–2.54) 9/119 0.93 (0.37–2.32) C/C – 7/11 5.29 (1.35–20.83)* 2/13 2.53 (0.51–12.46) 1/10 2.20 (0.26–18.45) T/T + 169/152 2.12 (1.33–3.38)* 192/69 21.11 (12.90–34.56)* 206/239 2.30 (1.27–4.16)* C/T + 68/50 2.39 (1.31–4.37)* 73/23 22.21 (11.87–41.54)* 81/90 2.89 (1.47–5.69)* C/C + 8/4 1.74 (0.41–7.30) 13/2 41.99 (8.91–197.88)* 14/5 5.03 (1.24–20.41)* SI — 1.89‡ 1.62‡ Pharyngeal cancer rs1054564 G/G – 9/342 1.00 16/425 1.00 5/255 1.00 C/G – 2/158 0.71 (0.14–3.49) 4/185 0.76 (0.24–2.39) 3/118 0.96 (0.21–4.49) C/C – 1/11 4.11 (0.33–51.37) 0/13 — 0/10 — G/G + 55 / 153 5.63 (2.54-12.49)* 48 / 70 8.89 (4.393-17.98)* 59 / 240 2.32 (0.82-6.58) C/G + 24 / 49 7.91 (3.17-19.75)* 22 / 22 14.05 (5.858-33.71)* 23 / 89 3.03 (0.97-9.50) C/C + 2 / 4 4.35 (0.60-31.36) 3 / 2 17.31 (2.465-121.56)* 3 / 5 3.17 (0.47-21.44) SI — — — rs1054221 T/T – 9 / 341 1.00 16/424 1.00 5/254 1.00 C/T – 2/159 0.71 (0.14-3.51) 4/186 0.76 (0.24-2.39) 3/119 0.96 (0.21-4.47) C/C – 1/11 4.11 (0.33-51.57) 0/13 — 0/10 — T/T + 54//152 5.60 (2.52–12.43)* 47//69 8.84 (4.36–17.91)* 58//239 2.30 (0.81–6.51) C/T + 26//50 8.16 (3.30–20.22)* 24//23 14.66 (6.18–34.74)* 25//90 3.13 (1.00–9.76)* C/C + 2//4 4.35 (0.60–31.41) 3//2 17.38 (2.48–121.93)* 3//5 3.15 (0.47–21.34) SI — — — Laryngeal cancer rs1054564 G/G – 7//342 1.00 11//424 1.00 2//254 1.00 C/G – 7//158 2.90 (0.87–9.64) 7//186 1.39 (1.37–1.42) 2//119 1.58 (0.21–11.75) C/C – 1//11 4.84 (0.32–74.26) 2//13 5.38 (5.19–5.58) 1//10 18.10 (1.26–261.14)* G/G + 18//153 3.10 (1.14–8.42)* 14//69 5.96 (5.88–6.04) 23//239 3.79 (0.81–17.79) C/G + 13//49 8.56 (2.82–26.00)* 13//23 20.69 (20.33–21.05) 18//90 12.10 (2.45–59.77)* C/C + 3//4 23.80 (3.59–157.93)* 2//2 — 3//5 17.42 (2.00–151.97)* SI//SIM§ 3.84‡//1.59§ — — rs1054221 T/T – 7//341 1.00 11//425 1.00 4//270 1.00 C/T – 7//159 2.91 (0.88–9.66) 13//283 1.40 (1.37–1.42) 14//283 1.57 (0.21–11.72) C/C – 1//11 4.83 (0.31–74.14) 2//69 5.37 (5.18–5.57) 2//70 18.08 (1.25–260.97)* T/T + 18//152 3.11 (1.14–8.46)* 11//39 6.08 (6.00–6.17) 10//39 3.81 (0.81–17.86) C/T + 13//50 8.48 (2.79–25.79)* 12//45 24.14 (23.73–24.56) 13//45 12.05 (2.44–59.60)* C/C + 3//4 23.85 (3.59–158.20)* 2//2 — 6//10 17.47 (2.00–152.37)* SI//SIM 3.85‡//1.59§ — —
†Odd ratios were adjusted with age, ethnicity and another two substance use. *P < 0.05 ‡Synergy index (SI) was evaluated for additive interaction model by Rothman’s synergy index.
Supplementary figure. SNP discovery of the GDF15 gene from 20 pairs of SCCHN patients and
controls in the study population
Region Genotype frequencies Genotype frequencies
5′ flanking region (promoter) – 473TC (rs1245 9782) MAF- T: 0.32 CC (52.8%) CT (31.5%) TT (16.7%) –396CT (rs5757 3498) CC (100%) – 119CT (rs1752 6126) MAF- T: 0.14 CC (97.3%) CT (2.7%) –20CT(rs7710 9188) CC (100%)
Exon 1 (coding sequence) +57GC (rs1059 519) MAF- G: 0.33 CC (50.0%) CG (35.0%) GG (15.0%) +170GA (rs6413 435) MAF- A: 0.13 GG (97.5%) GA (2.5%) +174A T (rs1059 369) MAF- T: 0.38 TT (40.0%) TA (45.0%) AA (15.0%) +195GT (rs1698 2331) GG (100%)
+56CT (rs1059 022) CC (100%) +143G T (rs1804 826) MAF- T : 0.34 GG (43.8%) GT (43.8%) TT (12.4%) +195C A (rs3746 195) CC (100%) +200G A (rs4558 6234) GG (100%) +281G A (rs4553 5632) GG (100%) +327CG (rs1058 587) MAF- T : 0.19 CC (62.5%) CG (37.5%) +538C A (rs1155 6750) CC (100%)
Region Genotype frequencies Genotype frequencies
Exon 2 (3′-UTR) +685G A (rs122 7733) MAF- T : 0.21 GG (62.5%) GA (32.5%) AA (5%) +689C G (rs105 5150) MAF- T : 0.35 GG (45.0%) GC (40.0%) CC (15.0%) +720G C (rs105 4564) MAF- T : 0.14 GG (75%) GC (22.5%) CC (2.5%) +763T C (rs105 4221) MAF- T : 0.14 TT (75%) TC (22.5%) CC (2.5%)
UTR: untranslated region; MAF: minor allele frequency.
Note: In total, 19 SNPs were discovered on the selected regions of the GDF15 gene by comparing the GDF15 SNP database from international populations at the HapMap Data Coordination Center (http://www.hapmap.org/).
Supplemetary table. PCR conditions for re-sequencing. Ampli
con Base
pair Forward primer Reverse primer Annealing temperature
1 756 5′-AGCACCCTGCTTAGACTGGA-3′ 5′-GGAGCATCTGAGAGC CATTC-3′ 59°C 2 788 5′-CAGCTGTGGTCATTGGAGTG-3′ 5′-CACACCCCCATTGTTT CTCT-3′ 64.5°C
3 820 5′-AAGCCATTCTTCTGCCTCAG-3′ 5′-CACATGGTCACTTGCA CCTC-3′ 55°C 4 789 5′-ACTGCTGGCAGAATCTTCGT-3′ 5′-AACACCTTGCCACTCA TTCC-3′ 57°C