EXPLOITATIVE COMPETITION OF
MICROORGANISMS
FOR TWOCOMPLEMENTARY NUTRIENTS IN
CONTINUOUS
CULTURES*
SZE-BI HSUt, KUO-SHUNG CHENG AND S. P. HUBBELL:I:
Abstract. Thispaper concernstheexploitative competition oftwomicroorganisms for two
complemen-tarynutrients inthecontinuous culture.Consumptionofthe limiting resourcesfollows theHollingType II
functionalresponse or, equivalently,Michaelis-Menten kinetics, generalized to the two-resource situation.
The predicted biologicalconditions whichshould giverise toeachof the possible competitive outcomes are
presentedin detailand analyzedglobally.Amajor conclusion is thateachof thefouroutcomes of classical
Lotka-Volterratwo-species competition theory hasmultiple mechanisticorigins in terms of
consumer-resourceinteractions.Itisalsoshownthat allfourclassical outcomes, including thecasein whichwinning
dependsonthe initialabundancesof thecompetitors, can arise for thispurelyexploitativecompetition.
Moreover,the outcomes of thisexploitativecompetition can bepredicted,inadvanceof actualcompetition, frommeasurementsmadeoneachspeciesgrownbyitselfonthe resources.
1. Introduction. The classical theory of ecological competition between twoor more species, attributed to Lotka
[18]
and Volterra[36],
is an extension of the basiclogistic model of single-species growththatdatesfrom Verhulst
[35].
The dynamical equationsfor thistheoryfor twocompetitors, 1 and 2, areoften written asdN
{1-(N+aN2)}
dN2
(1.1)
d---
rlN1
KI
dt 1(/3N,
+
N)}
r2N2
K2
where
Ni
isthe numberofthe ithcompeting species,ri andK
aretheintrinsicrate of increase andthe carrying capacityoftheithcompetitor, respectively, andaand/3
arethe interaction of "competition" coefficients, expressing the per capita competitive
effect of species 2 on 1, and 1 on 2, respectively. In the absence of competition
(c =/ 0),each populationgrowstoitsrespectivecarrying capacity. Inthe presence
ofcompetition,one orthe otherrivalmaysurvive while itscompetitordiesout,orelse therivalsmaycoexist.These three biologicaloutcomes result from thefour mathemati-cal casesthatcan occurprovided that populations of both speciesarepresentinitially. Competitive stability (coexistence) occurs when a
< K/K2 and/3 <
KE/K1;competi-tive instability (initial numbers of the competitors determine the eventual winner)
occurswhentheseinequalitiesarebothreversed; and competitivedominance
(one
orthe other specieswinsregardless of initialnumbers) occurswhenone but not both of
these inequalities arereversed.
Theclassical theorycanperhaps be called a"phenomenological" theory insofar
asitseeksto describehow the numbers of competing species change, and topredict the eventual outcome of such competition, without ever being specific about which
limiting resources are the focus of competition, nor about how effectively the rival
species forage for, and exploit, these resources. This classical theory has had an immenseand lastingappeal because ofitsgenerality andsimplicity, andithas been the
subject ofaverylarge numberof theoretical studies(cf.Wangersky’sreview
[38]).
Bythesametoken, however,thisgenerality has also madeit difficultforexperimentalists
to interpret and measure the theory’s critical parameters. It has proven especially *Received bythe editorsMarch 31, 1980, and in revised form December 1, 1980.Thiswork was partiallysupported by theNational Science Council of theRepublicof China.
tDepartment of Applied Mathematics, National Chiao-Tung University, Hsin-Chu, Taiwan 300,
Republicof China.
$DepartmentofZoology, University ofIowa, IowaCity,Iowa52240. 422
difficulttoestimatethe competitioncoefficientsindependently of actually growing the potential competitors together, and to determine theirvalues under fieldconditions, althoughconsiderable attentionhas beendevotedtotheseproblems.Usually,
competi-tion coefficients have.been estimated in laboratory competition studies by fitting the dynamical equations
(1.1)
to the growth curves of the species in competition (e.g.,Vandermeer
[34]).
The attempts to estimate competition coefficients have usually focused on various measures of overlap in resource utilization (Schoener[28]),
although information about whether the resources are limiting is usually lacking. Unfortunately, whenever these coefficients canonlybe estimatedfromthe dynamics of populations alreadyincompetition, thevalue ofthe theoryforpredictionisthereby diminished. Thus, the classical theory has been attackedas atautological exercise incurvefittingatbest
(Peters
[23]).
Atworstthefitof(1.1)
todataispoor (Wilbur[39],
Neill
[22],
Richmond et al.[26])
because of avariety of nonlinearities inper capitaratesofgrowthas afunctionof competitordensities.
Overthe past 25 years the elementsof a moremechanistic, resource-based theory of ecological competition have been under experimental andtheoreticaldevelopment byadiverse field
ot
workers.A
listot
all significant contributionsto thiseffortwouldbequite long, butsome
ot
themilestoneshave been papers by Herbertetal.[6],
Powell[24],
Holling[7], [8],
[9],
Miller[21],
Dugdale[2],
Eppley and Coatsworth[3],
Epply and Thomas[4],
Kilham[14],
MacArthur[19],
Stewart and Levin[30], Droop [1],
Koch[15],
LeonandTumpson [17],
Smithetal.[29],
Taylorand Williams[31],
TilmanandKilham
[32],
Tilman[33],
Real[25],
andHsuetal.[10l,
[12].
Thistheoryconsidersthe dynamics of theresourcesexplicitlyin addition tothe population dynamicsofthe competing species.
Moreover,
itpaysparticular attention to thefunctionalresponsesofthe competitors tochanges inresourcedensity.
Incomparisonwiththeclassicalapproachto competition, some thingsaregained andsome arelostby adoptingthisapproach.The disadvantages are that resource-based competitiontheoryisusuallylessgeneral,butalsomoredifficulttoanalyze
mathemati-cally, thanclassicaltheory.
However,
the advantagesarethatthe experimentalist hasaspecificset ofsomewhat more mechanistic questions toask about limiting resources and themannerin whichthe consumersrespond functionally and numericallytothese
resources.Thus,resource-basedtheory directsmoreexplicitattention toresourcesand consumer-resource interactions thanLotka-Volterra theory which, classicallyatleast, focused primarily if notexclusively onthe phenomenological changes in competitor numbers.
We
believe that this approach will spur renewed interest in experimentalstudies of competition, and we also hope that it hastens the development of more predictivetheory.
In
thispaperwepresentaresource-based competition modelwhichdescribes howtwomicroorganisms compete exploitatively for two complementary resources in the chemostat.
A
chemostat isalaboratoryapparatus used for production and physiological studyofmicroorganisms.In
the chemostat model, the limitingnutrient issuppliedat aconstant rate.The inputflowotmedium containsall other factorsforgrowthin excess.The output flow equals the input flow, and carries with it cells, waste product and unusednutrient.Themodelalsoapproximatesconditions forplankton growthinlakes,
with the input of complementary limiting nutrients such assilicaand phosphate from
streamsdraining the surrounding watershed.
2. Competitionforone resource. Before consideringtworesources,itisusefulto reviewbriefly whathappensinthe one-resourcecase.
We
assume that the consumption of a single limiting resource follows the HollingType II
functional response or,equivalently, Michaelis-Menten kinetics, which describes the chemical interaction
between enzyme and substrate. When aspecies grows on a single limitingresource, there is some "break-even" concentration of that resource at which birth rate just balances death rate,which we willcall the subsistenceconcentration.Nowsuppose that
this limiting resource is supplied at a constant rate, corresponding to the constant
carrying capacityenvironmentof classicalcompetitiontheory, and thattwospecies
(or
more)
arecompeting exploitatively forthisresource.Then,provided that theresource is not being suppliedata ratebelow the subsistenceconcentrationof all species, onlyasingle speciesispredictedtosurvive" that specieswhichhasthe smallestsubsistence concentrationof the resource
(Stewart
andLevin[30],
Taylor and Williams[31],
Hsuet al.
[10]).
Recently Hansen and Hubbell[5]
conducted a rigorous test of thisprediction. Thisnicelyintuitiveresultturnsout,however, tobedependentonhaving
a constantresource input. If theresourcehasaperiodic input, for example,seasonally
(Stewart
and Levin[30], Hsu [13]),
or if the resource is a prey species capable of self-reproduction (Koch[15],
Hsu et al.[12]),
then there are conditions when the speciescan
coexistinadynamic, periodicfashion on asingle resource(McGehee
andArmstrong [20]).
Fortheremainder of thispaper,we willbeconcernedwiththe caseofaconstantcarrying capacity environment, correspondingtoLotka-Volterratheory,
butin resource terms.
Thesubsistenceresourceconcentrationis animportant competition criterion,not
onlyinone-resource situations, butinthetwo-resourcetheorydiscussed in thispaper
aswell.Foreach species and limiting resource, thereissuchaconcentration, potentially
different in each case.Itwillbe symbolized by
J,
following Rosenzweig[27].
In
ourprevious workon thissubject10],
11],
wehave usedA for this parameter, but henceforth wewill use J to avoid confusion withthe finite rateof increase.Thesubsistence concentrationcan be calculated from three parameters that are measured
oneach species grown by itself onthe particular resource. Thus, forresources Sand speciesi,
Jsi
canbe found from"(a)
thehalf-saturation constant forresource uptake, Ksi;(b)
theintrinsic rateof increase,ri; and(c)
thedeath rate,Di.
Thehalf-saturationconstant
Ki
corresponds to K, in Michaelis-Menten theory, and represents thatconcentration of resourceat whichconsumption occursathalfthemaximalrate. The
subsistence concentration forspecies growth-limitedby resource Sistheproductof the half-saturation constant and the ratio of the death rate to the intrinsic rate of
increase"
\rsi/
The units of
Ji
arein resource concentration, since theunitsofDi
and rsicancel. The parameter ri isequivalenttormax
for theithspecies, rsi (msi-Di), wheremsi isthemaximalper capitabirthrate onresource S.
For
the detailed biological meaning ofJsiwereferto
[10].
The equations for single-resource competitionamongn species for resourceSare
d__S (sO)_
S)D
Y. ms--2
S N, dt i= yiKsi
+
S(2.2)
dNi
m,i$"Ni
DIN/, i=l,2,...,n. dtKsi
-t-SSisthe concentration ofresource, S)istheconstantinput concentration ofresource,
D
isthe constant rate atwhichnew nutrient is imported, as well as therate at whichnutrientat currentconcentrationisexported.
For
theithorganism,Ni
isthe population density,Ysi
isthe yield of the ith organism produced perunitofresourceS consumed,and
Ks,
m, andD
areasdefinedabove.Itwillbe recognized that(2.2)
isthe system of equations for any continuous culture ofseveral species, growth-limited byasingle nutrientbut supplied with all otherrequirednutrients innonlimitingamounts(TaylorandWilliams
[31], Hsu
et al.[10],
Hsu
[11]).
This is tobeexpected,sincecontinuous cultures were specifically developed as idealized environments having a constantcarryingcapacity. Thus,itshould be noted that a constantcarrying capacity doesnot
result from a fixed quantity of limiting resource, as in batch culture, but from a
steady-stateresourceinput-outputsituation.
In
(2.2),ifnoorganismsarepresent, then the resource equilibratesatS
()atthe point when resource input and outputrates arebalanced.
The one-resource situation can be summed up as follows. With no loss of generality, number the competing
species
such thattheirJ’s
areordered, withJs1 <
Js2
<’<
Jsn.
All species dieoutif theinputconcentrationislessthan thesubsistence concentration for every species, i.e., ifS()<Jsl.
In
this case, limt_ooS(t)= S() andlimt_,oo
N(t)-
O,i-1,...,n.On
the otherhand, ifS()>
Ji for any i,then species 1survivesand outcompetes all rival species.
In
thiscase,limt_,ooS(t)
J,
limt_,oN(t)
y,x(S()-Js)
andlimt_,ooN(t)=
O, 2,...,n.3. Competition[ortwo resources.
In
situationsinvolvingtwo or moreresources,itbecomes necessary for the firsttime to considerhow the resources,onceconsumed,
interact topromotegrowth.
Leon
andTumpson [17]
have distinguishedtwoimportant classes ofresources: complementary andsubstitutable. Complementary resources aresources of differentessentialsubstanceswhich aremetabolically independent
require-mentsfor growth, suchas a carbon source andanitrogen sourcefor a bacterium,or
silica and phosphorus for a diatom. On the other hand, substitutable resources represent alternate sources of the same essential substance, and are metabolically interdependent requirements for growth, such as two carbon sources or two sources forphosphorous.
In
thispaperwe considerjust thecase ofcomplementaryresources.Nowconsider twocomplementary resources,
R
andS.For
eachconsumerspecies thereare twoJ’s,onefor eachresource.TheseJ’sarethesubsistence concentration ofeach resource when the species is growth-limited by that one resource alone. For
species i, call theseconcentrations
Jr
andJ
of resourcesR andS,
respectively. TheseJ
valuesdetermine the position of the zero-growthisocline for species on the S-Rresource plane.
Thezero isoclinefor complementaryresources is a pairof half-lines meeting at
rightanglesatthe point
(J,
.]rri)in theS-R
plane (Fig.1).
Thelines areperpendicular because ofthe independence of the requirements forR
and$.In
thiscase, growthislimitedatany giventimeeitherby
R
orbyS,
butnotby bothR
andSsimultaneously exceptatthecorner).
The curving dashedlinepassingthroughthe cornerintheisoclinerepresents the equation, miS/(Ki
+
.9)
mrR/(K,.
+
R),
where the parametersareaspreviouslydefinedbut subscripted for the appropriate resource. Above the dashedline inthetheS-R plane, species isS-limited, whereas below the dashed line, species is R-limited (Fig.
1).
Thus, for example, when species is S-limited, no increase inresource
R
inthe region above the dashedcurve willhave any effectonincreasing the growth rate of species i; only an increase in resource $ will have this effect. The converse istrue inthe region below the dashedcurve.Before presenting the competition modelsfor twospecieson twocomplementary resources, we should discuss how the functional responses of the consumer species
Jri
/
i-th predoor isocline
=0
/ msiS mriR
/
/ Ks +S Kri+R
FIG.
have been generalized from one to two resources. In the one-resourcecase, the per capita consumption rate, according to the
Type II
functional response, is given by(mri/yri)(R/(Kri+R)) if the resource is
R,
or is given by (rnsi/ys)(S/Ks+S))if the resource is $. Now we generalize the functional response to two complementaryresources.
In
this case, the per capita consumption rate of whichever resource iscurrentlylimitinggrowthis identical tothe one-resource per capita consumption rate, as given above for the appropriate resource. The question thenarises. Atwhatrate is
the nonlimitingresourceconsumed?Thisquestioncanbeansweredwhenweconsider the yield ofconsumerproducedperunitof resourceconsumed. When the yieldfactors,
Yri and Ys are constants, then it follows that there must be a fixed ratio of the growth-essential substances provided by resources
R
and S in a unit of consumer.Moreover,
this also implies that the per capita consumption rate of the nonlimiting resource must be proportional to the per capita consumption rate of the limiting resource. Ifit were not, then theratioofessentialgrowth substancesintheconsumerwould be changing, and the yield factors wouldnolongerbeconstant.The proportion-alityconstant is the ratio ofthe yield constants for the two resources. For example, suppose species is S-limited. Then the per capita consumptionrate of
S,
callitf(S),
is
msi S
(3.1)
f
(S)
ys, Ks,
+
S’
whereas the concurrentper capita consumptionrate of the nonlimiting resourceR is
given by
(3.2)
Ys--2/’
f
l(S)ms____
SY
,.
YrKs
+
SNote that the expression in
(3.2)
does not contain the concentration of the nonlimitingresourceR.
Thus, itshould be noted that:For
complementary resourcesR
andS,
whena species isS-limited, itspercapitaconsumptionrateof
Risindependentof
theconcentrationof
R,
whereas, when thespecies isR-limited,itspercapita consumption rateof
S isindependentof
theconcentrationof
$. The keyto this statementis: "Whenis aspeciesS-limited?".The speciesisonlyS-limitedabove a certain concentration of
resource
R (above
the dashedline inFig.1).
Belowthis concentration ofR,
the per capita consumption rate ofR
does depend on the concentration ofR;
but thisdependence is because the species is now R-limited and no longer S-limited. The
converseargumentapplies when the speciesisR-limited.
4.
Statement
of the model. Given the precedingdevelopment of the biologicalbasisforthe functional response of species exploiting pairsofcomplementary
resour-ces, it is astraightforward matter tostate the two-resource, two-species competition model in the continuous culture.
Forcomplementary resources,
R
andS,
and species 1 and 2 competing exploita-tively forthem,the system of equationsisdS 1 1 .-7
S
S)D
g(S, R)N
g2(S,R
[a Ysl Ys2(4.1)
dR 1 1d---
(R
0R )D
gl(S,
R)NI
g2(S,
R)N2,
Yrl Yr2dNa
dNz
=[g(S,R)-D]N, =[g2(S,R)-D]N, dt dtS(O) >
O,R (0) >
O,N(O) >
O,N2(O) >
O,where
rnsiS
mrtR
)
gS, R
minK-
-
-S
Kr
+
R
(
ms S
mr _R_
g2(S, R
minI-2--S
Kn
+
R
I"
As
noted previously,this model assumes constantcarrying capacities (fixedrateof nutrient input), constant yield factors (unit of consumer produced per unit of resourceconsumed),and
Type II
functionalresponses, generalizedto two complemen-tary resources.In
addition,wehaveassumedType II
numericalresponsesinpercapitabirthrates, with a directdependenceonthe external supplyofresources. Finally,we
have assumed the same dilution rate
D
forS,
R, N, N2.
The parameters in(4.11)
have all been previously defined atvarious pointsin the text, but itiscon-venient to relist them here in oneplace for ease of reference"S
,
R
=
input concentrations of resourceS andR,
respectively.D
input flow rate of medium containingS,
R and also the output flow rate mediumcontaining unusedS, R
and cells N1,N2.
ms, tnr maximalpercapitabirthrateof species onresourceS or
R
alone.Ysi,Yri yield of species perunitof resourceSorR consumed.
K,
Kr
half-saturation constantfor species onresource S orR.We analyze the behavior of solutions of this system of ordinary differential
equations in ordertoanswer the biological question, under whatconditions willneither, one,orboth species surviveordieout?
We
alsoseekto determine thelimiting behavior of the surviving species and the resources.5.
Statement
ofresults.In
this section westatethe principal results of thepaper.The proofs and certain technical lemmas are deferred to 6. The first lemma is a statementthat the system given by
(4.1)
is as "well-behaved" as oneintuits from thebiological problem. Theproof of the lemma is similar tothat in
[10],
and we omitit.LEMMA5.1. Solutions
o[ (4.1)
are positive and bounded.Furthermore, wehave(5.1)
S(t)
S() Ng(t)+o(1)
ast-,i= Ysi
(5.2)
R(t)=R(o)_
Ng(t)+o(1)
ast-->eo.)]ri
The next lemma providesconditions under which the organismscannot survive
given thefixed dilutionrate and thefixedinputrates ofthe nutrients.Beforewestate
the lemma,wenotethe followingtwopairsofequivalentstatements, namely,
msiS
() KsiDs(O)
Ksi-Jr-S(0)
<
D ifandonlyif msi<--Dormsi
D>
mriR
(o)KriD
(o)Kri
-1-R
(o)<
D ifand onlyif mri<=
Dormri
D
>
RLEMMA5.2.
(5.3)
then limt_,oNi(t) O.
msiS
()mriR
(o)Ks
q-s(O)
<
D orKri
-[-R(o)< D,
Lemma5.2statesthe necessaryconditions forspecies
Ni
tosurvive, i.e.,KgD
s(O)
K,.D0<< and
0<<R().
msi D mri D
Since nutrients S and R are complementarily essential to the growth of speciesNi,
there are minimum input concentrations S() and R() both for S and
R
in order tosupport the speciesNi.
COROLLARY5.3.
If
(5.3) holdsfor
1,2, then limt_ooS(t) S(),
limt_,R (t) R(o)andlimt_,oNi (t) O, 1, 2.
We state the principal result in the case of inadequate input concentration of nutrientsin four parts.We areable to determine theglobally asymptoticbehavior of
the solutions in Theorem 5.4. The theorem may be summarized by noting that the unsuccessful competitor doesnot affectthe eventual behaviorof thesurvivor andits
resources.
Beforewestate Theorem 5.4,weintroducethe following important parameters"
Jsi Jri 1,2, msi D mri D
Ci
yiTi
R (0) Jri ..,(o) 1 2. Jsi YriTHEOREM 5.4(i), (ii). Let
(5.3) hold]or
=2 andO<Jsl
<S(0),
O<Jrl
<R().
(i)
If
T1
<
C1, then the trajectoryof
(4.1)
approaches theequilibrium(Es
1)
as -->oe,
where
Nsl*
(Esl)
(JI,R*
sl,Nsl,*0),
Nsl
* ysl(S-Jsl)
R*
R ()sl (o)
Yrl
(ii)
If
T1 <
C1, thenthetrajectoryof
(4’1)
approaches theequilibrium(Erl)aswhere
(Erl)
(Sr*,
Jrl,N’r1,
0),
N*
,1Y,I(R()-
Jl),THEOREM 5.4(iii), (iv). Let
(5.3)
hold]’or
i=1 and O(Js2(S(0),
O<Jr2<R
().
(iii)If
T2
>
C2,thenthe tra]ectoryof
(4.1)
approaches theequilibrium(Es2)
aswhere
(Es2)
(Js2,R*
s2,O, N*s2)
N*
s2 ys2(S -Js2), R R(1
,
(o)Ns2
s2
Yr2
(iv)
If
T2
< C.,
thenthetrajectoryo
(4.1)
approachesthe equilibrium(Er2)
aso,
where
(Er2)
(Sr2,
Jr2,O, Nr*2
N*
r2Yr2(R
--Jr2),
Sr2(0)
,
=s(O)Nr*2
Ys2
Theorem 5.4 statesthat whenlimt_,ooNi(t)=0 the equation
(4.1)
isreduced to asystem of three ordinary differential equations.
In
order to present the biological meaningofparametersT,
Ci, weassume limt_,ooN2(t)=
0.We
mayrewrite T1,C1
as(R()-Jrl)D
1/yrlT1
(s(O)_jsl)
D
C1-
1/yslWhen only species 1 is present,
T1
represents the ratio of the steady-state nutrientregenerationrates atequilibrium under consumption by 1.
Jr1
andJsl
aretheequilib-rium concentrationsofresourcesR and
S,
respectively, understeady-stateconsump-tionby species 1. The parameter
C1
represents thefixedyieldratiofor species 1 growingon resources
R
and S. The units of (1/yrl) are (unitsR consumed/unit
species 1produced); thus
C1
is the ratio, (units Rconsumed/units
Sconsumed)
per unit ofspecies 1produced.
By
comparingT1
with C1, we can determine whether species 1 is S-limited or R-limited. This is becauseC1
represents the invariant ratio in which the essentialnutrients
R
andSareconsumed by species1,whereasT1
represents theratioin whichthesesame resourcesare beingexternallyregeneratedundersteady-stateconsumption
pressurefrom species 1.Therefore,if
T1 >
C1,thegrowthrate ofspecies 1is S-limitedbecause S is regenerating at a steady-state rate slower than
R
with respect to the required consumptionratiofor species 1. Similarly,ifT1
<
C1,the growthrateof species 1 willbe R-limited. The aboveconsiderationsalsoapplytospecies 2forappropriate values ofT2
andC2.
DEFINITION. If
T1
>
C1
(or T1 < C1),
we say that speciesN1
is S-limited(or
R-limited). Similarly, if
T2
>
C2
(or
T2
<
C2), wesay that speciesN2
is S-limited(or
R-limited).
Remark.
We
mayjustify the concept ofR-limited orS-limited.Consider system(4.1) withN2--0. Suppose N1
isR-limited, thenatequilibrium(S*rl,
Jr
1,Nr*l
wehaveD
(mrlJrl)/(Krl
+
Jrl)<
(mslS*
)/(Ks
+
Srl*
and henceS*
>
Js
ThenmrlJrl
N*rl
yrl(R()-
Jr1
Krl+Jrl
DYSl(S
(0)
S
rl
<
Ys(s(0)
Js
1)
andsoT1
<
C1.
Similarly, ifN1
is S-limitedthenC1
<
T.
In order to discuss the interior equilibrium point, we may assume as a basic
hypothesis
(H1)
S() >
Jx,Js2 >
0,R()>Jrx,
Jr2>O.
Under the assumption (HI), the equations of
(4.1)
maybe relabeled without loss ofgenerality,so thatwemay assume either
(H2)
Jl
<Jr2,Jl
<J2
or
(H3)
J
< Jr,
J2<J.
We
note that most of the conditions on parameters for the various cases inTheorems,5.5, 5.6canalso beestablishedby thelinearizationmethod.
Beforewestateour mainresults,Theorems5.5and5.6,weintroducethe following parameters"
T*-R()-Jr2
N*c
ysl(S()-Jsl)(C2-
T*)
Nc
Ys2(S()-JI)(T*-C1)
S(0)
Ys
C1
C1
C2
C1
THEORZM 5.5.
Assume (H1)
and(H2)
hold(see
Figs. 2a, 2b). LetC1
C2.
Thenthestatementsin Theorem 5.4(i) (ii) hold.
R R
Ir2
Jr1
(Esl) GS GS: Globally Stable.Tr2’
.Trl (Erl) I, =-S I,JSl
-Ts2
JslJs2
a. N1isS-limited. b. NisR-limited. FIG.2Theorem 5.5statesthat,if onespecies has the lowerJ’sfor bothnutrients
S
andR,
then that specieswill surviveandits rival will not.Now we consider the situation when each species has the lower subsistence
concentration on one oftheresources, say,
Jr1 <
Jr2,J2 <
J
1.Then theparametersT*,
C1
andC2
become importantinthe competitionoutcomes.First westate thefollowing results describing howonespeciescanoutcompetethe other.
THEOREM 5.6(i).
Assume (H1)
and(H3)
hold. LetC1
C2.
If
T* <
C1, C2, then thestatements in Theorem 5.4(i), (ii) hold(see
Figs. 3a,3b).
THEOREM 5.6(ii). Under the assumptions
of
part (i),if
T*>
C1, C2, then thestatements in Theorem 5.4(iii), (iv) hold
(see
Figs.4a,4b).
J’r2
Jr1
U: Unstable (Er2) (Es) UJs2
-Ts
R Jr2 .Trl FIG.3 (Er2)(E-r1)
Js2 b S GS UI(Esl)
.Ir2 .Irl GS(Er2)
Js2
Is1 Is2 a FIG. 4 (Esl)In
order to explain the biological meaning of Theorem 5.6(i), (ii), we rewriteT*
(R
(o)Jra)D/(S
()Js
1)D,
whichrepresents theratio ofthe steady-stateregener-ationrateof
R
whenN.
isalone and that ofSwhenNt
is alone.We
notethatunder the assumption(H3)
wehaveT* <
T1
andT*
>
Ta.
First weconsider Theorem5.6(i).The assumption
T* < C,
Ca
impliesTa
< Ca;
i.e., speciesNa
isalwaysR-limited.But,
since species
N
has the lower subsistenceconcentration for resourceR,
speciesNt
alwayswins.
Note,
however,thatatthe one-species equilibrium, speciesN
may again beeither S-limited or R-limited. Similarily,we canexplain Theorem 5.6(ii).SpeciesN2
outcompetes speciesN1
forthereasonthat speciesN
is S-limitedandN2
has the lower subsistence concentration for resourceS.Secondly,wedescribe howtwospeciescancoexist.
THEOREM 5.6(iii). Under the assumptions
of
part (i).ff
C1 < T* <
C2, then the "positive" equilibrium(E)=
(J,Jr2,N,
N)
exists and is globally asymptotically stablein thefirst
orthant(see
Fig.5).
In
this case, we note thatC
< T,
andT2
<
C2; i.e.,N1
is S-limited andN2
isR-limited.
But,
species N1,N2
has lower subsistence concentration forR
and Srespectively. Coexistence occursbecause eachspecies has the lower subsistence con-centration forthatresourcewhich,atthetwo-speciesequilibriummixtureofresources,
most limitsthegrowthof itsrival.
Jr2
Jr1
U (Es) GS(E)
Js2 sl (Er2) FIG.5Finally,westatearesult describing theoutcomeof competition dependsoninitial
populations. Thereare two cases. Thefirstone
((a), (d))
isthecase when both speciesare limitedby thesameresource.The secondone((b),
(d))
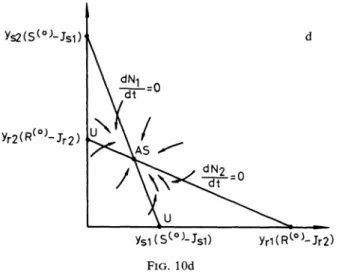
isthecasewhenthe species arelimitedbydifferentresources.THEOREM 5.6(iv). Under the assumptions
of
part(i),//C2 < T*<
C1, then(Ec)
existsandunstable.Furthermore, wehave
four
possibleoutcomes:(a)
If
NI
is S-limitedandN2
isS-limited, then(Es)
and(Es2)
areasymptoticallystable.
(see
Fig.6a).
]’r2
J’rl
AS (Es2)
U AS
AS: Asymptotically Stable
(Ec) (Esl)
Is2
Isl
Jr2
Jr1 AS, Js2 ,(Es2) (Ec) b (Erl);s
.lr2 Jr1 (Er2) (Ec)u
(Esl) ASJs2
]sl C FIG. 6 Jr2Jr1
Er2) (Ec)A=S U"
/s2
]sl (Erl)(b)
If
N1
isR-limitedandN2
isS-limited, then(Erl)
and(Es2)
are asymptotically stable (see Fig.6b).(c)
If
N1
isS-limitedandN2
isR-limited, then (Esl) and(Er2)
areasymptotically stable (see Fig.6c).
(d)
If
Nx
isR-limitedandN2
isR-limited, then(Erx)
and(Er2)
areasymptoticallystable
(see
Fig.6d).This case arises because eachspecies has thelower subsistence concentration for
that resource which, atthe two-species equilibrium mixture of resources, leastlimits
the growth ofitsrival.Thismakes the two-species equilibrium unstable. Theoutcomes
dependonwhether species
Nx
and speciesN2
areindividually S-limited or R-limited atthe one-species equilibrium.6. The
Proof of
Lemma 5.2. From(4.1),
itfollows thatLete
>
0 be chosen such that[
m,(S+
e)
min\K-Z (--o5 )
D,
rnri(R
(o+
e)
_D O,K
+
(Ro+
andfrom(5.1),
(5.2)
chooseto>
0such thatS(t)<-S+
e,R(t)
<-R+
e, >-to.Then, foranappropriateconstantC,
itfollowsthatNi(t)
<=
CNi(O)exp{[min
\Ki
+
(S(
+
e)
mri(g()+e)
-O)] .(t-t0)}.
-D,
Kr
+
(R
o+
e)
Hencelimt_,Ni(t) 0. Q.E.D.
Beforeweprove Theorem5.4(i),(ii), (iii), (iv),Theorem5.5andTheorem5.6,we
present the followingidea which reduces the problem of thefour-dimensionalsystem ofdifferentialequations
(4.1)
toaproblemof a two-dimensionalsystem ofdifferentialequations.
Considerthe
Lyapunov
functioni=1Ysi i=1 Yri
for
(4.1).
Itfollows that+
N
s(O)
N
(o)+
R+
-R _-<0.i=1Ysi i=1Yri
Hence
E
{(S, R, N, N2)"
fr
0}
{
(S,
R, N, N2)"
S) =$+N
YsiR(=R+
,S>-_O,R>-O,N>-O,i=I, 2 i=1YriThen the to-limit set f of thetrajectoryof
(4.1)
lies in E[16],
and it is sufficient tostudy thebehavior ofsolutions ofthe followingtwo-dimensionalsystem:
(6.1)
wheredN1
dN2
NaGI(Nx, Nz), N2G2(N1,N2),
dt dtNI(0)
=>
0,N2(0)
=>
0,i=1)gsi i=1
i=1,2.
Proof
of
Theorem. 5.4. First we prove (i) and (ii). Since(5.3)
holds for i=2,from Lemma 5.2 we have limt_,N2(t)=0. Then the trajectory of
(4.1)
approachesE f3
{(S, R,
N1,N2):N2
0},
and it suffices to consider the equationdN1/dt
N1G
(N1, 0),NI(0)
->0, or, equivalently,(6.2)
dt__=Nl[min
{(msl-D)(S()-Jsl-N1/ysl)
[ -i
Z-
7y--1
(mrl-D)(R()-Jrl-N1/Yrl))]
;-[
+
R(o)_N1/)2r1
NI(0)
=>
0.If
T1
>
C1, i.e.,(R()-Jrl)Yrl >
(S()-Jsl)ysl,
thenN1
0andN1
(S()-Jsl)ysl
arethe(S()
only two possible equilibria of
(6.2).
And limt_,ooNl(t)=Ns*x
ysl -Jx), pro-videdNI(0) >
0 in(6.2).
Since0<Jx
<S(),
O<Jrl
<R(),
from the third equationof(4.1)
it is impossible that limt_,oNl(t)=O. Then there exists (S,R,N1,0)s
withN1 >
0. Thenpart (i)follows directly fromthe invariancepropertyof to-limit setand thefact(EI)
isasymptotically stable.If
TI
<
Ca,thenusing theabovearguments yields theproofof (ii).The proofforpart (iii), (iv)respectivelyis similar to that ofpart (i)and (ii). Q.E.D.
Before weprove Theorem 5.5 andTheorem 5.6,westate a theorem of Markus
[40]
which willbeused repeatedly.DEFINITION.
LetA"
xi =fi(x,t)
andAo:xi =fi(x)(i 1.2,..,
n)
be a first-order systemofordinarydifferentialequations. The real-valuedfunctionsfi(x,t)
andfi(x)for continuous in (x,t)
for x sG, where G is an open subset ofR
n,
and for>
to" they satisfyalocalLipschitzcondition in x.A
is said tobe asymptotic toAoo(AAoo)
in Gif, for eachcompactsetK
_
Gandforeache>
0, thereis a T T(K,e) >
tosuch that[fi(x, t)-fi(x)l <
e for all 1, 2,.,
n, allxK,
all> T.
DEFINITION. The fl-limitset for
x’=
f(x,
t),X(to)
Xoisthe set of to-limitpoints y,wherey limn_,oox(tn)
forsomesequence{tn},
t,c.
THEOREM(Markus).
Let
A-,A
inGandletPbeanasymptotically stable criticalpoint
of
Ao.
Then there isa neighborhoodNof
P
andtimeT
such thatthe l’l limitsetfor
everysolutionx(t)
of
A
whichintersectsNata timelaterthan TisequaltoP.Next,
weneedto describe the isoclinesdNx/dt
O,dN/dt
0of(6.1)
for variouscases. The proofs will be based on these geometric figures. First we note that the
transformation
2
Ni (0) Ni
(6.3)
S S()-i=1
is 1-1 from the S-R plane intothe
NI-N2
plane providedCa
#Cz.
The equations in(6.1)
canberewritten asdN1
dt(6.4)
dN2
N2
dt(ms2_D)(S(O)_jsz
N
zz)
(mra_D)(R(O)_Jr2
Nx
rZZ)
in ysl YrlKs2 +
S
(0) NiNz
K,z
+
R
<o)Na
N2
Ysl Ys2 Yrx
Nx(0)_>-0,
N2(O)O
Theisocline
dN/dt
0, 1, 2, in theNa-Nz
plane can be classified intofour cases bymappingtheisoclinein the $-R plane(see
Fig.1)
under thetransformation(6.3).
Case 1.
Ti
>=
Ca, Cz,i=1,2(see
Fig.7a).
Case 2.
T
<-Ca, Cz, 1,2(see
Fig.7b). Case 3.Ca
--<
T,.
_-<C2, 1, 2 (seeFig. 7c).Case
4.C2
--<
Ti
-<_Ca, 1, 2(see Fig.7d).We
note that theisoclinesof(6.1)
or(6.4)
aresimilartothose in the Lotka-Volterra competition model(1.1),
and hencewe canstudythebehaviorof solutions of(6.1)
by isocline analysisor by pushing trajectory.Proofof
Theorem 5.5.Assume
Tx
>
Ca.
SinceCa
#C2,the nonsingulartransforma-tion
(6.3)
mapstwodisjointisoclinesof(4.1)
inFig. 2aor2bintotwo disjointisoclinesof
(6.1)
in theNa-N2
plane.Combining thetwoisoclines
dNa/dt
O,
dN2/dt
0 of(6.1)
yields seven various figures. The isoclinedNa/dt
0of(6.2)
is ofthe type in Fig. 7aorFig.7c.Onthe other hand, theisoclinedN2/dt
0 has four various forms. These forms are similar inourdiscussion, and we only need to pick one of themin this proof. For example, Fig. 8
isthecombinationof Fig.7a,Fig.7drespectively forisoclines
dNa/dt
O,dN2/dt
0N2
Yr2(R(OLJri
\ \ \ S:Jsi \Ys2(StO)_
Jsi dN \ \ Yrl( (o) -J’ri Ysl(S(OL.Tsi’.
R=J’ri \ FIG. 7aN2 Yr2(R()- Jri Yr(L* N2 Ys2 S()-J’si Yr2(R(J-Jri) Yr2 R
(.o)_.lr
Ys2(S()-.Tsi)
%.
dNi,
-d’i" o dN0
,,
Ys1(S(’L.1"si)
’Yrl
R()-.Tri
N2
N \ \ \dNi
< 0 dldNidt
>0 Yrl(R(’LJri FIG. 7b-dYsI(S( L.1"si
N2 Yr2
(R()-Jrl)
I dN U -aT- <0 Yr2(S )-.It2)dN2
Ys(R()-Js2)
ys (S(.)-Jsl) F]o.8 N(see Fig. 8). If NI(0)>0, N2(0)=>0 then, from isocline analysis, the trajectory
of
(6.2)
approaches(ysl(S(-Jsx),
0).Consider the trajectory (S(t), R(t), N(t),
N2(t))
of(4.1).
First we claim limt_,oN(t)# O.Suppose
limt_,ooN(t)=
0; then from(HI)
and Lemma 5.1 we havelimt_,N2(t)#0.Hence there exists a point (S,
R,
0, N2)elfor some $=>
0,R >
0,N2
>0 where l’l is the o-limit set of the trajectory (S(t), R(t), N(t),N2(t)).
By theinvariance property of theo-limit set, we have
(Es2)e
I) (ifT2
> C2)
or(Er2)
I1 (ifT2
< C2). Compare
the followingtwosystemsof differentialequations"(6.5) dS
(s(O)
S)D
1 gl(S,R)N1
1d--;
Ys--x
ys--
g2S, R N2
dR 1 1d---
(R(o)_R)D
gl(S, R)Nx g2(S, R)N2, YrX Yr2dN2
dt[g2(S, R)
D]N2;
(6.6)
dS(s(o)
S)D
1d--
ys--z
g(S, R)N2,dR
(R
(o)_R)D
g2(S,R)N2, dt yr2dN2
[g2(S, R) D]Nz.
dtObviously, under the assumptionlimt_,ooNx(t)=0we havethat
(6.5)
isasymptoticto(6.6).
Since(Esz)
11 (ifTz
> Cz)
or(Ez) I1 (ifTz
<
Cz)and(’2)
or(’z)
is asymp-totically stablefor system(6.6),
where(/r2)--(Sr2,
.Jr2, Nr*2
),(/s2)-"
(Js2,R*s2,
Ns2:
),Markus’ theorem yields that limt_,oS(t)
S*2,
lim_,oR(t)
J2, lim_.oNE(t)N*
r2 orlimt_,S(t)
Js2,limt_,R (t)
R*
2,limt_,ooNE(t)
Nz.
* Either caseimpliestheunboundedness of
Nx(t). (see
Fig.2a,Fig.2b).This isthedesired contradiction.Hencelim,_,oo
N(t)
#O.Since limt_,oo
Na(t)
O,
itfollows thatthere exists a point(,,/,/Qa,/Qu)
e D,with,
=>
0,/ =>
0,/Qa
>
0,N2
0. The trajectory(Na(t),
N2(t))
with initialvaluesNa(0)
1,
N2(0)=
2
approaches(Ns*,
0).
This and the invariance property of the o-limitset, fe
E,
imply(Esl)efLBut(Ea)
isasymptotically stable.Hencelim,_.oo(S(t),R (t),
Na(t),
N2(t))=
(E,a).Forthe case
Ta
<
C2,similararguments yield that(Era)
isglobally asymptotically stable. Q.E.D.Proof
of
Theorem 5.6(i), (ii).Firstweprovepart(i).The proofofpart(ii)is similar tothat of part (i)andwe omit it. SinceJr
<
Jr2,J2
< J,
1, itfollows thatT*
R()-Jn
s(O)_js
<
Ta
R()-Jrl
T*
R()-Jr2
s(O)_Jsl
S(Jsl
>
T2
S()-Js2
From the assumption
T* <
C1, C2, we haveTz
< Ca,
C2.
Applying the assumptions(H3)
andT*< Ca,
C2
yields four possible cases for the forms of the isoclinesdNa/dt
O, dN2/dt
0of(6.2).
Case
1.Ta >=
Ca,
Cz
(see
Fig. 9awhichcorrespondstoFig.3a).
Case 2.
Ta
<-
Ca,
C
(see
Fig. 9bwhichcorresponds toFig. 3b).Case
3.Ca
<-Ta
<-C2
(see
Fig. 9cwhichcorrespondstoFig. 3a).Case4.
Cz
<-TI
<-Ca
(see Fig. 9d which corresponds to Fig. 3b). Using theargument of Theorem5.5 yields the proofofpart(i). Q.E.D.
N2
Ys2 s(L
Jsl
Yr2(R(oL.Tr2 ASYr1(R()--.Tr2
Ys1(S(L.sl)
Yr2(R(,)-.Tr
Yr2(R()-]r2)
dN1
n Yr1(R()-Jr2) Yr1(RtLJrl) FIG.9a-bYr2 (RCL.l"rl)
Yr2(R(OL]r2)
=0 Yrl( R(o)-Jr2)
Ys2 R(o)_]slYr2(R()-.Tr2
Yrl(.R(OL.Tr2) Yrl(R(O)-.lrl
FIG.9c-dProof of
Theorem 5.6 (iii). We note that under assumption(H3)
we haveT*<
Ta, T*>
Tz.
SinceCa
<
T*< C2
wehave four possible cases whichall corespond toFig. 5"
Case 1.
Ca
<-Ta
<=
Cz,C1
<-_Tz
<-Cz
(see
Fig. 10a). Case 2.Ca
<-Tz
<-Cz,
Ta
>-_Ca,
Cz
(see
Fig. 10b). Case 3.Ca
<-Ta
<=
Cz,Tz
<=
Ca, C/(seeFig. 10c). Case 4.T2
<=
C1, C2,Ta
>=
Ca,
C2
(see
Fig. 10d).First wenote that from linear stability analysis
(Ec)
isasymptotically stable. Using the similar arguments in the proof of Theorem 5.5 yields that limt_,ooNa(t)0,limt_,oo
N2(t)
O. Bythesamearguments as inthe proof of Theorem 5.5 it suffices toshow that there .exists a point (S,
R,
Na,N2)
fl with N>
0,N2 >
0. We have the following three possible cases.Case a.
Na(t)
>-N’c
for all -_>to
forsome to.Since limt_,oNz(t)
0,there exists apoint
(S-,/,/a,/2)
’
with/a
->N’,/2
E for somee>
0.Case b.
Na(t)<-N’
for allt>-to
for some to.In
thiscasewe havetwosubcases: Subcase 1. limt_.Na(t)=c >0 forsome c. Since limt_.ooNE(t)
0, there exists apoint
(S,
R,
c,N2)
flwithNI
c>
0,N2
->e forsome e>
0.Subcase 2.
limt.o Na(t)
doesnotexist.Then thereexist e>
0 andasequence{t,}
with
Na(t,)>
e and(dNa/dt)(t,)=O. We
may choose a subsequence {t,i} such thatYr2
(R()-Jrl)
Yr2 (R(.)-Jr2) N2
l
U’.
ysl(S o)_.is1),ysl
(S(.)_js-
N1
N2
Ys2 S()-.Tsl Yr2 (R()-.Tr2) YsI(S(0)-sl)
Ys1(S(*)--Ts2 NYr2 (R(O)-.]’rl)
Yr2(R(O)-.]’r2) Ysl(S()-Jsl)Yrl(R(*
L.,Tr2
FIG. 10a-c/ \ / dN2 0
s
S(. Yrl R,o)_.Ir2)FIG. lOd
either S(tni)=Jsl for all t,i or R(t,i)=Jrl for all t,. If S(&i)=Jsl for all t,i, then
from
(5.1)
wehave(6.7)
N2(t.,)y[
(s(-j,)
N(t_.,)]
+
o(1)
[
+
o(1)=
+
>-- Ys2 (S()--Jsl)--YslJLet
tn
oo andchoose anappropriatesubsequenceof tn; thereexists apoint (Jsl,R,
Q1,
Q2)
efZwith/Q1
>eand/q2
>=
N*2c.
IfR
(t,)Jl
for all t,, thenfrom(5.2)
wehave(6.8)
N2(t,i)
y,2[
(R()Nx(t")]+o(1)
Yr
[
>--Yr2
(R()-J’r2)-YrtJHencethere existsapoint
(S-,
Jr1,1,/Q2)
elwithJr
>E and.r2
_->N2*c.
Casec.Nl(t)oscillatesaround
N1
N*c,
Then thereexists{&}
with(dN1/dt)(&) <
0 andNI(&)=N*lc.
In
this case we may choose a subsequence{&i}
such that eitherS(&)<-Jsl for all t,i or R(&i)<-Jrl for all t,i. Then from
(5.1)
or(5.2)
we still have inequalities(6.7)
and(6.8).
Hencewecomplete theproofof Theorem5.6(iii). Q.E.D.Proof
of
Theorem 5.6(iv). Fromthe assumption thatC2 < T* <
C1, the "positive" equilibrium(E)
exists. That(E)
is unstable follows directly from the assumptionC2
<
T*
<
C1, and thelinearstability analysis about(E).
The resultsareobvious from Figs. 6a,6b, 6c, 6d. Q.E.D.Remark. In describing Theorems 5.5 and 5.6 we take
C1#
C2
as an essentialassumption. What can happen when
C1
C2?
The meaning forC1
C2
isthat thefixedyieldratiofor species 1 growingonresources
S
andR
isequaltothefixedyieldratiofor species 2 growingonresources
S
andR. In
thiscase, theisoclines in(6.4)
areparallel linesintheN-N2
plane. Sincethe proofs are thesameas or even moresimple than the proofsfor the caseC1
#C2,wemerelystate the results here andomitthe proofs.Letusdefinefor convenience
Nii=
min{ysi(S()-Jsi), yri(R()-Jri)},
andwithoutlossofgenerality assume thatJ
<
Js_.(i) If
Jr
< Jra,
thenN
>
N
and lim (S(t), R(t), NI(t),N.(t))
(S
()t-++
Ysl
(ii) If
Jra
<Jr1
andNll>
N2I,theni,]= 1,2,
lim (S(t), R(t), Nl(t),
Nz(t))
(S
)t--,+
(iii) If
Jra
< Jr1
andNll <
N21, then_,R(O)
N11
N11,0)
lim
(S,(t),R(t),NI(t),N.(t))=(S
)Naa
t-,+
Ys2
(iv) If
Jr2
(Jrl andNll
N21,then,R(O)
Nll
Nll,0)
___,R(O)
N22
--
0N22
)
lim (SCt), R(t), Nl(t),N2Ct))
(Js2,Jr1,N*,
N
),whichdependson initialconditions.
N*
andN’
satisfy the following equationN* N*
$(o)N
--+--
-Js2
or--+--=
R()-Jrl.
Ysl Ys2 Yrl Yr2
This means that all points on the line
N1/Ysl +N2/Ys2-
S()-Js2
on theN1-N2
plane(onthe nonnegativeoctant)areequilibrium points.
7. Discussion.
In
this paper, we have explored the behavior of an exploitative competition model which describeshowtwospecies compete fortwocomplementaryresources.
The analysis has revealedthateachofthe classicaloutcomes oftwo-speciesLotka-Volterra competition theory can arise in two or three different ways when resource dynamics and consumer-resource interactionare explicitlyconsidered.
Leon and
Tumpson [17]
discuss the competition between two species for twocomplementaryorsubstitutableresources.Interested readersmayfindthe mathemati-calanalysis and biological discussion forsubstitutableresourcesin apaper ofWaltman,
Hubbell and
Hsu
[37].
The equationsin(4.1)describeshowtwospecies compete fortwocomplementary
nutrients inthechemostat. Tilmanand Kilham
[32]
and Tilman[33]
have performed interesting competition studies in semicontinuous cultures between two freshwater diatoms, Asterionellaformosa
Hass.,
and Cyclotella meneghinianaKutz
forthecom-plementary resourcesphosphate and silicate.They did otreport any casesin which
theoutcomesweredependenton initialnumbers.
However,
theydid find abroad regionof coexistence over a range of ratios of silicate/phosphate in the influentsupply to semicontinuouscultures of thetwo diatomspecies.
We
havetaken the data providedinTilman[33]
tosee ifthere isany possibility ofacasein whichtheinitialnumberofAsterionella orCyclotella woulddeterminethe outcome of competition.Let
JPA
amdJSA
be theJcriteria forAsterionella onphosphateand silicate, respectively, and let
JPc
andJsc
be the corresponding J criteria forCyclotella. If we assume thatallcelldeath was duetowashout fromthe culture in the
effluent, then themaximum deathratetheystudiedexperimentally was 0.5/day,i.e.,
D
0.5/day.Then thevaluesof theJcriteriaareJPA
----0.025/xM
(micromole),JSA
3.28/zM,
JPc
0.417/zM
andJsc
0.90/zM.
ThusJPA
< JPc,
sothat Asterionella hasalowersubsistenceconcentrationonphosphatethanCyclotellabymorethananorder of magnitude, but
Jcs
<
JAS, SOthat Cyclotella hasalower subsistence concentration onsilicatethan Asterionella.Next,
it is necessary to computeT*,
CA,
andCc,
whereCA
andCc
are the CcriteriaforAsterionella and Cyclotella, respectively,
T*
(p(O_
JPc)(S(0)--
JSA)
where
p(O
and S( arethe input phosphate andsilicate concentrations, respectively, andthe point(JsA,JPC)
istheintersectionofthe Asterionella and Cyclotella isoclines onthe silicate-phosphateresourceplane. Of the rangeofvaluesofp(O
andS(tested by Tilman, we chose p(O=10/zM.
and S(=100M.
This gives a value forT*=
9.9x10-2.
Finally, it is necessaryto compute the C criteria forthe two diatoms. The yield
constants for Asterionella are reported by Tilman
[33]
to be"YPA=
2.18X
10
cells//zM
on phosphate,andYSA--2.51
106
cells//zM
onsilicate.There-fore,
CA
(1/ypA)/(1/YSA) 1.15 10-2.
The yieldconstants forCyclotella areYPc
2.59107
cells/tzM
on phosphate,Ysc=4.20106
cells//.tM
on silicate. Thus,Cc
(1/Ypc)/(1/Ysc) 1.62x 10-1.
With this information, wecan answer the question ofwhether there canexist a
casein whichthe winningdiatomspecies(AsterionellaorCyclotella)isdeterminedby theinitialcell density of each diatom.
We
note thatJPA
<
JPc
andJSA
>
JSA.
Next,
we notethatCA
<
T* <
Cc.
Thiscorrespondstoa caseof coexistence, Theorem.5.6(iii),afact that Tilman
[33]
confirmed experimentally.In
order for there to be a case inwhichtheinitial diatomdensitydeterminestheoutcomeinthiscompetitive systemfor
these
J’s,
itwould be necessary that the inequalitiesamongCA,
T*,
andCc
betotally reversed:CA
>
T*>
Cc.
This,in turn, would require substantial changesin the yieldconstants forphosphate and silicate in thesetwo diatomspecies.Sinceonly thecriterion
variable
T*
involvesparameters under experimentalcontrol, thereis no possiblity ofacase in which initialcelldensitiesaffect the competitiveoutcomebetweenAsterionella and Cyclotella.
We
note thatit ispossibleforT* < CA,
Cc
orT* > CA,
Cc
such thatT*
is an experimental parameter.In
either of these cases, onlyone species survivesandcoexistence doesnotresult.
REFERENCES
[1] M. R. DROOP, Thenutrientstatusofalgalcellsin continuousculture, J.MarineBiol.Assoc. U.K.,54
(1974),pp. 825-855.
[2] R.C. DUGDALE,Nutrientlimitation in seadynamics:identificationand significance,Limnol.Oceanogr. 12(1974),pp. 685-695.
[3] R. W.EPPLEYANDJ. L. COATSWORTH,UptakeofnitrateandnitritebyPitylumbrightwelliimKinetics
and mechanisms,J.Phycol., 4(1968), pp. 151-158.
[4] R. W.EPPLEYANDW.H. THOMAS,Comparisonof half-saturationconstantsforgrowth andnitrate
uptakeofmarinephytoplankton,J.Phycol., 5(1969),pp. 375-379.
[5] S.R.HANSENANDS.P.HUBBELL,Single-nutrient microbialcompetition: agreementbetween
experi-mental and theoreticallyforecastoutcomes(subm.toScience,1979).
[6] D. HERBERT, R.ELSWORTHANDR. C.TELLIIKIG,Thecontinuouscultureofbacteria: atheoretical
and experimentalstudy,J. Gen.Microbiol.,14(1956),pp. 601-622.
[7] C. S.HOLLING, Thecomponentsofpredationasrevealed byastudyofsmallmammal predationofthe Europeanpinesawfly, Canad. Entomol., 91(1959),pp. 293-320.
[8]
,
Thefunctionalresponse ofpredatorstoprey density and itsrole in mimicryand populationregulation,Mem.Entomol.Soc. Canada,45 (1965),pp. 3-60.
[9]
,
Thefunctionalresponseofinvertebratepredatorstopreydensity,Mem.Entomol.Soc. Canada,48(1966), pp. 1-86.
[10] S.-B.Hsu,S.P.HUBBELLANDP.WALTMAN, Amathematicaltheoryofsingle-nutrient competition
incontinuouscultureso[microorganisms, this Journal, 32(1977), pp. 366-383.
[11] S.-B.Hsu,Limiting behavior]:orcompeting species, this Journal, 34(1978), pp. 760-763.
[12] S.-B.Hsu, S. P. HUm3ELLANDP.WALTMAN, A contributiontothe theoryofcompetingpredators,
Ecological Monographs,48(1978),pp.337-349.
[13] S.-B. Hsu, Acompetitionmodel]’oraseasonally fluctuatingenvironment,J.Math.Biology,toappear.
14] P.KILHAM, Ahypothesisconcerning silicaandthefreshwaterplanktonic diatoms,Limnol.Oceanogr.,
16(1971),pp. 10-18.
[15] A.L. KOCH, Competitive coexistenceo[twopredators utilizing thesamepreyunderconstant
environ-mentalconditions,J.Theoret. Biol.,44(1974),pp.373-386.
[16] J. P.LASALLE, The StabilityofDynamicalSystems,CBMS Regional Conference Series inApplied
Mathematics25, Societyfor Industrial andApplied Mathematics, Philadelphia, 1976.
[17] J.
m.
LEON,AND D. B. TUMPSON, Competition between twospecies]’ortwocomplementaryortwo substitutableresources, J. Theoret.Biol., 50(1975),pp. 185-201.[18] A. J.LOTKA, Elementso[MathematicalBiology,WilliamsandWilkins,Baltimore,MD,1925.
[19] R. H. MACARTHrOa,GeographicalEcology,HarperandRow, NewYork, 1972.
[20] R. MCGEHEE AND R.
m.
ARMSTRONG, Some mathematical problems concerning the ecologicalprincipleo]’competitiveexclusion, J.DifferentialEquations,23(1977), pp. 30-52.
[21] R. S.MILLEa, Patternandprocess in competition,Adv.Ecol.Res.,4(1976),pp.1-47.
[22] W. E.NEILL, Thecommunity matrixand interdependenceofthecompetitioncoecients,Amer.Nat., 108(1974),399-408.
[23] R. H. PETERS,Tautologyin evolutionandecology,Amer. Nat.,110(1976),pp.1-12.
[24] E.O.POWELL,Criteriaforgrowthofcontaminants andmutantsincontinuousculture, J. Gen.Microbiol.
18(1958),pp. 259-268.
[25] L. A.REAL, Thekineticsof functionalresponse,Amer. Nat.,111(1977),pp.287-300.
[26] R. C. RCHMOND, M. E. GILPIN, S. PEREZ SALAS AND E. J. AYALA, A search foremergent competitivephenomena: the dynamicsofmultispecies Drosophilasystems,Ecology,56(1975), pp. 709-714.
[27] M.L. ROSENZWEG,Evolutionofthe predator isocline,Evolution,27(1973),pp. 84-94.
[28] T. W. SCHOENER, Somemethods]’orcalculatingcompetitivecoefficients fromresource utilization spectra,
Amer.Nat.,108(1974),pp. 320-340.
[29] O. SMITH, H. H. SHUGART, R. V. O’NEILL, R. S. BOOTH AND D. C. McNAUGHT, Resource
competitionandan analytical modelofzooplankton feedingonphytoplankton,Amer. Nat., 109 (1975),pp.571-591.
[30] F. M. STEWART, AND B. R. LEVIN, Partitioning of resources and the outcome of interspecific
competition:amodel andsomegeneralconsiderations,Amer. Nat.,107(1973),pp. 171-198. [31] P. A. TAYLOR,ANDJ. L.WILLIAMS, Theoreticalstudies onthecoexistenceofcompeting speciesunder
continuous-flowconditions,Canad. J.Microbiol.21(1975),pp.90-98.
[32] D. TILMAN,ANDS. S. KILHAM, Phosphate andsilicategrowth and uptakekineticsofthe diatoms,
AsterionellaformosaandCyclotella meneghiniana, J. Phycol., 12(1976),pp.375-383.
[33] D.TILMAN, Resourcecompetitionbetween planktonic algae:anexperimental and theoretical approach,
Ecology, 58(1977),pp. 338-348.
[34] J. H. VANDERMEER,Thecompetitive structureofcommunities anexperimental approach withprotozoa,
Ecology,50(1969),pp. 361-371.
[35] P. H.VERHULST,Noticesurlaloi quela populationpursuitdanssonaccroissement,Correspond.Math.
Phys., 10(1838),pp. 113-121.
[36] V. VOLTERRA,Variationsandfluctuationsofthe numberofindividualsofanimalspecies livingtogether,
AnimalEcology,R. N.Chapman, ed.,McGraw-Hill,NewYork, 1926.
[37] P. E. WALTMAN, S. P. HUBBELLAND S.-B. HSU, Theoreticalandexperimental investigation of
microbial competition in continuouscultures,Proc.ofConf.onMathematicalModelling,
Carbon-dale,II.,1979.
[38] P. J. WANGERSKY,Lotka-Volterra population models.Ann. Rev.Ecol.Syst.,9(1978),pp. 189-218.
[39] H.WILBER, Competition,predation, and thestructureofthe Ambystoma-Rana Sylvaticacommunity,
Ecology 53 (1972),pp. 3-21.
[40] L. MARKUS, Asymptotically autonomous differential systems, in Contributions to the Theory of
NonlinearOscillation,vol.3,PrincetonUniversityPress,Princeton,N.J.,1956, pp. 17-29.