Tongue Cancer Cells Through
Mitochondria-Dependent and -Independent Pathways
Siu-Wan Ip,1* Sheng-Hui Lan,1* An-Cheng Huang,2Jai-Sing Yang,3 Ya-Yin Chen,4,5Hui-Ying Huang,1Zen-Pin Lin,6Yuan-Man Hsu,7 Mei-Due Yang,8Chang-Fang Chiu,9,10Jing-Gung Chung7,11 1
Department of Nutrition, China Medical University, Taichung 404, Taiwan 2
Department of Nursing, ST. Mary’s Medicine Nursing and Management College, Yilan 266, Taiwan
3
Department of Pharmacology, China Medical University, Taichung 404, Taiwan 4
School of Chinese Medicine, China Medical University, Taichung 404, Taiwan 5
Department of Chinese Internal Medicine, China Medical University Hospital, Taichung 404, Taiwan
6
Department of Holistic Wellness, Mingdao University, Changhua 523, Taiwan 7
Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan
8
Department of Surgery, China Medical University Hospital, Taichung 404, Taiwan 9
Graduate Institute of Cancer Biology, China Medical University, Taichung 404, Taiwan 10
Division of Hematology and Oncology, China Medical University Hospital, Taichung 404, Taiwan
11
Department of Biotechnology, Asia University, Wufeng, Taichung 413, Taiwan
Received 29 April 2010; revised 26 July 2010; accepted 27 July 2010
ABSTRACT: Although there have been advances in the fields of surgery, radiotherapy, and chemotherapy of tongue cancer, the cure rates are still not substantially satisfactory. Capsaicin (trans-8-methyl-N-vanillyl-6-nonenamide) is the major pungent ingredient of hot chili pepper and has been reported to have an antitumor effect on many human cancer cell types. The molecular mechanisms of the antitumor effect of capsaicin are not yet completely understood. Herein, we investigated whether capsai-cin induces apoptosis in human tongue cancer cells. Capsaicapsai-cin decreased the percentage of viable cells in a dose-dependent manner in human tongue cancer SCC-4 cells. In addition, capsaicin produced DNA
Correspondence to: J.-G. Chung; e-mail: [email protected] Contract grant sponsor: Department of Health, Executive Yuan, Republic of China (Taiwan).
Contract grant number: DOH99-TD-C-111-005.
*These authors contributed equally to this work.
Published online in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/tox.20646
fragmentation, decreased the DNA contents (sub-G1 phase), and induced G0/G1 phase arrest in SCC-4 cells. We demonstrated that capsaicin-induced apoptosis is associated with an increase in reactive oxy-gen species and Ca21generations and a disruption of the mitochondrial transmenbrane potential (DCm).
Treatment with capsaicin induced a dramatic increase in caspase-3 and -9 activities, as assessed by flow cytometric methods. A possible mechanism of capsaicin-induced apoptosis is involved in the activation of caspase-3 (one of the apoptosis-executing enzyme). Confocal laser microscope examination also showed that capsaicin induced the releases of AIF, ATF-4, and GADD153 from mitochondria of SCC-4 cells.#2010 Wiley Periodicals, Inc. Environ Toxicol 00: 000–000, 2010.
Keywords: capsaicin; human tongue SCC-4 cells; apoptosis; mitochondria; caspases-3
INTRODUCTION
Cancer is the major cause of death in the world. In the United States, one in every four deaths is from cancer. Oral and pha-ryngeal cancers number over 300,000 cases annually with male outnumbering female worldwide (Pintos et al., 2008). Based on the 2008 report from the Department of Health, R.O.C. (Taiwan), it was indicated that 9.6 individuals per 100,000 die annually from oral cancer in Taiwan. In the oral cavity, oral tongue squamous cell carcinoma (SCC) has been shown to be associated with the highest rate of metastasis when compared with other tumor sites (Sano and Myers, 2007). Although cures for oral cancer including surgery, radiotherapy, and current chemotherapeutic options are still inadequate, there is still focus on identifying new agents and novel targets for treating oral cancer (Ichimiya et al., 2005). The best strategy for treatment of cancer from chemothera-peutic agents is to induce apoptosis of cancer cells. It is well documented that some of the phytochemicals present in me-dicinal herbs exerts antitumorigenic activity via inducing apo-ptosis in cancer cells.
Capsaicin, a homovanillic acid derivate, is an active pharmaceutical ingredient of red pepper, genus Capsicum, and has been shown to induce apoptosis of many tumors cells (Hail, 2003; Ito et al., 2004; Amantini et al., 2007). Several reports have shown that reactive oxygen species (ROS) act as the principal signaling molecules in capsaicin-induced apoptosis (Lee et al., 2004; Zhang et al., 2008; Huang et al., 2009), but other reports also showed that cap-saicin treatment reduced endogenous ROS levels in several cell lines (Oh et al., 2004; Baek et al., 2008). It was reported that p53 is important for capsaicin-induced growth arrest and apoptosis in human myeloid leukemia NB4 cells (Ito et al., 2004). Recently, it was reported that capsaicin induces apoptosis in prostate cancer cells including wild-type p53-expressing LNCaP cells, p53-null PC-3 cells, and p53-mutant DU-145 cells (Mori et al., 2006). It was also reported that capsaicin induced apoptosis in cultured cells derived from human cutaneous SCC via the increase in ROS generation and disruption of the mitochondrial trans-membrane potential (Hail and Lotan, 2002). Therefore, the exact molecular mechanisms of capsaicin-induced apopto-sis in human oral or tongue cancer cells still remain unclear. The purpose of this study is to investigate a series
of experiments regarding the apoptogenic action of capsai-cin on human tongue cancer cells inclusive of cytotoxicity, cell-cycle arrest, and proteins involved in apoptosis. We offer more information to show that capsaicin induced apo-ptosis via mitochondrial-dependent and ER stress pathway in human tongue cancer SCC-4 cells.
MATERIALS AND METHODS
Chemicals and Reagents
Capsaicin, dimethyl sulfoxide (DMSO), propidium iodide (PI), Triton X-100, and trypan blue were purchased from Sigma Chemical Co. (St. Louis, MO). RPMI-1640, fetal bo-vine serum (FBS), penicillin–streptomycin, trypsin–EDTA, and glutamine were purchased from Gibco BRL (Invitrogen, Grand Island, NY). Caspase-3, -8, and -9 activity assay kits were obtained from OncoImmunin (Gaithersburg, MD).
Cell Line
The human SCC-4 cell line (human oral SCC) was obtained from the Food Industry Research and Development Insti-tute (Hsinchu, Taiwan). The cells were cultured in RPMI-1640 medium containing 10% FBS, 2 mM L-glutamine,
100 U/mL penicillin and 100 lg/mL streptomycin in 75-cm2 tissue-culture flasks at 378C under a humidified 5% CO2 and 95% air atmosphere as we have previously reported (Chen et al., 2009, 2010).
Determinations of Viability and Cell-Cycle
Distribution of SCC-4 Cells
The SCC-4 cells were plated into 12-well plates at a density of 2 3 105cells/well and maintained for 24 h. Cells were treated with 0, 200, 250, 300, and 400lM capsaicin, while only adding DMSO (solvent) for the control regimen and grown at 378C, 5% CO2, and 95% air for 12, 24, 36, and 48 h. For viability determination, after treatment, cells were harvested, and the number of viable cells was determined by the flow cytometric assay cited previously (Tan et al., 2006; Ji et al., 2009). For cell cycle and sub-G1 phase determination, the harvested cells were washed with phos-phate-buffered saline (PBS) and were fixed in 70% ethanol
at2208C overnight then resuspended in PBS containing 40 lg/mL PI and 0.1 mg/mL RNase and 0.1% Triton X-100 in a dark room for 30 min at 378C and analyzed by flow cytometry (Becton-Dickinson, San Jose, CA) equipped with an argon ion laser at 488-nm wavelength as described previously (Ji et al., 2009).
Determinations of DNA Damage
and DNA Fragmentation
Approximately 23 105cells/mL of SCC-4 cells in 12-well plates were treated with 200, 250, and 300lM of capsaicin for 24 h. Cells in each treatment were stained with 4,6-diami-dino-2-phenylindole dihydrochloride (DAPI) and then were examined and photographed using a fluorescence microscope as previously described (Ji et al., 2009). Approximately 13 106cells/well of SCC-4 cells were treated with 0, 200, 250, 300, and 400lM of capsaicin and incubated for 24 h. Cells from each treatment were harvested by centrifugation, and the DNA was isolated for DNA gel electrophoresis. It was then followed by EtBr staining and then examined and photo-graphed under UV light as described previously (Chen et al., 2009; Yang et al., 2009).
Determinations of ROS, Ca
21Production
Levels, and Mitochondrial Membrane
Potential (
DC
m) in SCC-4 cells
Approximately 23 105cells/mL of SCC-4 cells in 12-well plate were treated with 300lM capsaicin for 0, 3, 6, 12, and 24 h. The cells from each treatment were harvested and washed twice by PBS then resuspended in 500 lL of 2,7-dichlorodihydrofluorescein diacetate (10 lM) (DCFH-DA contains fluorescence staining for ROS) and Indo 1/AM (3 lg/mL) (dye contains fluorescence for staining of Ca21) and DiOC6(1lmol/l) (dye contains fluorescence for staining of MMP). All samples were then incubated at 378C for 30 min to detect percentage of changes in ROS, Ca21, andDCm before flow cytometry as described previously (Lin et al., 2009; Lu et al., 2010).
Determinations of Protein Levels Associated
with Cell-Cycle Arrest and Apoptosis in SCC-4
Cells After Exposure to Capsaicin
Approximately 1 3 106 cells/mL of SCC-4 cells were treated with 300lM capsaicin for 0, 12, 24, 36, and 48 h, and then cells were harvested. The harvested cells were lysed in a radioimmunoprecipitation assay buffer contain-ing PhosSTOP (Roche Applied Science). The Western blot-ting analysis was performed as previously described (Lin et al., 2009; Lu et al., 2010) using monoclonal anti-P21, -P16, -cyclin D, -Cyclin E, -CDK2, -CDK6, -AIF, -cytochrome c, -pro-caspase-9, -active-caspase-9,
-pro-cas-pase-3, -PARP, -Bax, -Bid, -ATF6-a, -IRE1-a, -GRP78, -pro-caspase-12, -Fas ligand, and -act-caspase-8 (Santa Cruz Biotechnology). Then all samples were stained by secondary antibody. The signal was detected with an enhanced chemiluminescence Western blot analysis system (GE Healthcare) (Lin et al., 2009; Lu et al., 2010).
Confocal Laser Scanning Microscopy for
Protein Translocation
Approximately 53 104cells/well of SCC-4 cells were cul-tured on four-well chamber slides for treatment without or with 300lM capsaicin for 24 h. Then cells on the slides were fixed in 4% formaldehyde in PBS for 15 min, permeabilized with 0.3% Triton-X 100 in PBS for 1 h with blocking of non-specific binding sites using 2% BSA as described previously (Chen et al., 2009; Kuo et al., 2009). Primary antibodies to AIF, ATF-4, and GADD153 (1:100 dilution) (green fluores-cence) were used to stain the fixed cells overnight, washed twice with PBS before being stained with secondary antibody (FITC-conjugated goat anti-mouse IgG at 1:100 dilution), and followed by DNA staining with PI (red fluorescence). All samples were microphotographed using a Leica TCS SP2 Confocal Spectral Microscope (Chen et al., 2009).
Statistical Analysis
The results are showing in mean6 SD, and the difference between the capsaicin-treated and control groups were ana-lyzed by Student’s t test, a probability of P\ 0.05 being considered significant.
RESULTS
Capsaicin Affected the Total Human Tongue
Cancer Viability and Cell-Cycle Distribution
of SCC-4 Cells
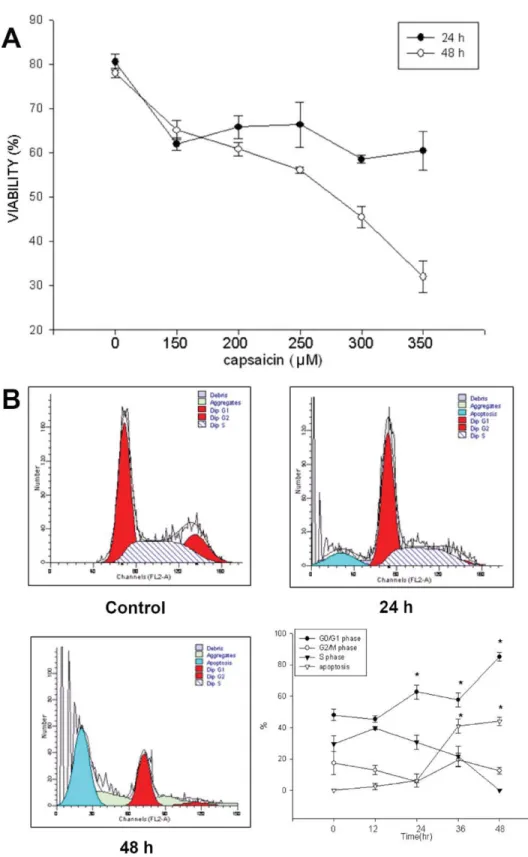
The results of flow cytometric analysis are shown in Figure 1(A,B), and they indicate that increasing the dose of capsaicin and/or time of incubation led to decrease the percentage of viable cells. The capsaicin at 350lM significantly decreased the viable cells by almost 65% [Fig. 1(A)]. The cell-cycle dis-tribution of SCC-4 cells after treatment with different doses of capsaicin for 0, 12, 24, 36, and 48 h is depicted in Figure 1(B). The cell cycle tended to be arrested at theG0/G1 stage by dose-dependent [Fig. 1(B)] after exposure to capsaicin. The sub-G1 peaks, indicating the proportions of apoptosis, increased in a dose-dependent manner when the concentra-tion of capsaicin added increased [Fig. 1(B)].
Capsaicin Induced DNA Damage and
Apoptosis in SCC-4 Cells
To investigate and confirm that capsaicin induced apoptosis via DNA damage, the SCC-4 cells were treated with
Fig. 1. Capsaicin affects the total human oral cancer viability and cell-cycle distribution of SCC-4 cells. Cells were incubated with or without 0, 150, 200, 250, 300, or 350lM cap-saicin for 24 h and 48 and then were harvested for determining the percentage of viable cells and the distribution of cell cycle by flow cytometry as described in Materials and Methods section. Data represent mean6 SD of three experiments. *P \ 0.05. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
capsaicin and isolated for DAPI staining and DNA frag-mentation determination. The results are presented in Fig-ure 2(A,B), which indicated that capsaicin induced DNA condensation [Fig. 2(A)] and DNA damage and fragmenta-tion at 400-lM capsaicin treatment [Fig. 2(B)].
Capsaicin Affected the Productions of ROS
and Ca
21and the Levels of Mitochondria
Membrane Potential (
DC
m) in SCC-4 Cells
To investigate whether or not capsaicin induced apoptosis is involved in mitochondrial dependent, SCC-4 cells were treated with 300 lM capsaicin before cells were isolated for examining the levels ofDCm, and the results are shown in Figure 3(A), which indicated that capsaicin decreased the levels ofDCmin SCC-4 cells. To address the possibility that the capsaicin-induced apoptosis could be related to contributions from the mitochondrial pathway, the changes in ROS and Ca21 productions were further examined, and the results are shown in Figure 3(B,C). In contrast, adminis-tration of capsaicin resulted in a right shift of the DCF and dihydroethidine fluorescence curves, indicating the increase in ROS [Fig. 3(B)] and Ca21[Fig. 3(C)] generations. These effects are time-dependent manners.
The Effects of Capsaicin on the Associated
Protein Levels of Apoptosis in Human Tongue
Cancer SCC-4 Cells
To reconfirm that the cytotoxic effect of capsaicin on SCC-4 cells, as shown from flow cytometric assay, came from G0/G1 arrest and apoptosis, SCC-4 cells were cultured with 300lM of capsaicin for 0, 12, 24, 36, and 48 h before being isolated for determination of protein levels and assayed by Western blotting. The results are shown in Figure 4(A–E). The figures indicated that capsaicin increased the levels of p21 and p16 [Fig. 4(A)] and decreased the levels of cyclin D, cyclin E, CDK2, and CDK6 [Fig. 4(A)] leading to the G0/G1 phase arrest in SCC-4 cells. Capsaicin increased the levels of AIF, cytochrome c, active-caspase-9 [Fig. 4(B)], Bax [Fig. 4(C)], ATF6-a, IRE1-a, GRP78 [Fig. 4(D)], Fas ligand, and active-caspase-8 [Fig. 4(E)], but decreased the levels of pro-caspase-3 [Fig. 4(B)] and Bid [Fig. 4(C)] that led to apoptosis in SCC-4 cells.
The Effects of Capsaicin on the Location of
AIF, ATF-4, and GADD153 in SCC-4 Cells
To confirm the translocation and levels of AIF, ATF-4, and GADD153 in SCC-4 cells after exposure to capsaicin, the SCC-4 cells were exposed to 300lM of capsaicin for 24 and 48 h and stained and examined and photographed by confocal laser microscopy. The results showed in Figure 4 indicate that capsaicin promoted the levels of AIF, ATF-4, and GADD153 that reconfirm the results from Western blotting analysis.
DISCUSSION
Much evidence has shown that capsaicin induces apoptosis in many human cancer cells. However, the molecular
Fig. 2. Capsaicin induced DNA damage and apoptosis in SCC-4 cells. Cells were treated with different concentrations of capsaicin (0, 200, 250, 300, and 400lM) for 24 and 48 before being harvested for DAPI staining (A) and for DNA gel electrophoresis (B) as described in Materials and Methods section. The arrow bar ‘‘:’’ revealed the DNA condensation (an apoptotic characteristic). Scale bar, 50lm. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
mechanism that triggers capsaicin-induced apoptosis in human tongue cancer cells is unclear. Herein, we demon-strate that capsaicin-induced apoptosis is mediated in part through the mitochondria based on the observations: (a) we have found that capsaicin induced cytotoxic effects of
SCC-4 cells in a dose- and time-dependent manner. cin-induced cell death is correlated with ROS. (b) Capsai-cin treatment affected the levels of cell-cycle associated proteins, which led to G0/G1 arrest and induced apoptosis through mitochondrial-dependent pathways. (c) Capsaicin
Fig. 3. Capsaicin affected the productions of reactive oxygen species (ROS) and Ca21, the levels of mitochondria membrane potential (DCm) in SCC-4 cells. Cells were treated
with 300lM of capsaicin for 0, 3, 6, 12, and 24 h before being collected and stained with DiOC6for theDCmlevels determined (A), stained by 2,7-dichlorodihydrofluorescein
diace-tate for ROS levels determined (B) and stained by Indo 1/AM for Ca21levels determined (C) as described in Materials and Methods section. The % in Y-axis of the bar diagram refers to % cell count with fluorescence *P \ 0.05. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
induced AIF release from mitochondria. (d) Capsaicin ex-posure also leads to dysfunction of mitochondria and induc-tion of caspase-9 and caspase-3. All these findings clearly indicate that capsaicin-induced dysfunction of mitochon-drial might be one of the critical events leading to apoptosis.
In this study, capsaicin does promote the ROS and Ca21 levels; therefore, our results also showed that capsaicin
treatment induces ER stress, which elicited a rise in intra-cellular Ca21, and subsequent mitochondrial membrane depolarization (decreased the levels ofDCm), followed by mitochondrial release of cytochrome c and AIF, then conse-quent activation of caspase-9 and -3. On the basis of these observations, we may suggest that Ca21-mediated signaling is involved in capsaicin-induced apoptosis, and this is also in agreement with other reports (Lee et al., 2004; Wu et al.,
Fig. 4. The effects of capsaicin on the associated protein levels of apoptosis in human tongue cancer SCC-4 cells. Cells were incubated with or without 300lM capsaicin for 0, 12, 24, 36, and 48-h incubations, and the cells were collected for Western blotting as described in Materials and Methods section. A: P21, P16, cyclin D, Cyclin E, CDK2 and CDK6; B: AIF, cytochrome c, pro-caspase-9, active-caspase-9, pro-caspase-3, PARP; C: Bax and Bid; D: ATF6-a; INEI-a, GRP78 and pro-caspase-12; E: Fas ligand and act-cas-pase-8 and Bid.
2006; Huang et al., 2009). It was reported that AIF is a mi-tochondrial intermembrane flavoprotein (Vahsen et al., 2004). Other reports already demonstrated that apoptosis is induced when AIF is translocated from the mitochondria to the nucleus to induce caspase-independent peripheral chro-matin condensation and large-scale DNA fragmentation (Susin et al., 1999; Ye et al., 2002). Our results also showed that capsaicin increased the Bax level but decreased the Bcl-2 levels. It was reported that mitochondrial outer brane permeabilization by proapoptotic Bcl-2 family mem-bers is involved in the release of AIF from the mitochondria (Otera et al., 2005). Increasing evidence have been found regarding the cancer chemotherapeutic agents modulating and interfering with mitochondrial function for promoting permeability of the mitochondrial membrane and cell death (Costantini et al., 2000). It means that capsaicin induced ap-optosis partly through caspases-independent pathways. Our results also showed that capsaicin promoted the levels of GADD153 and Ca21. This is in agreement with other reports, which demonstrated the generation of oxidative stress in capsaicin-treated cells (Lee et al., 2004; Vahsen et al., 2004; Otera et al., 2005; Wu et al., 2006). In the SCC-4 cells, we have measured the production of ROS upon capsaicin exposure and antioxidant NAC partially
recovered capsaicin-induced cell death (data not shown). The levels of 300 lM of capsaicin in the present study in vitro is higher than other phytochemicals, but it is lower
Fig. 5. Capsaicin promoted the release of AIF, ATF-4, and GADD153 in SCC-4 cells. The cells were exposed to 300lM of capsaicin for 24 h and then were stained and examined and photographed by confocal microscopy as described in Materials and Methods sec-tion. Mitochondria and nuclei were counterstained with PI (red color), respectively. Control and capsaicin treatment: AIF, ATF-4, and GADD153. Scale bar, 20lm. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Fig. 6. The proposed possible signal pathways of capsaicin induced cell-cycle arrest and apoptosis in human tongue cancer SCC-4 cells. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
than that of other reports, which showed that actual daily intake of capsaicin in some countries that use more chili peppers is much higher (estimated at 2.8 mg/kg body wt) (Lopez-Carrillo et al., 1994; Otera et al., 2005). Apparently, further investigations are needed in the future.
These observations from the present studies may suggest that capsaicin-induced apoptosis is likely mediated through the generation of oxidative stressed on the ROS and Ca21 production. We found that capsaicin treatment induced the expression of ER stress-related proteins such as IRE1, GADD153/Chop, and GRP78/Bip (Figs. 4 and 5). It is well known that the oxidative stress could potentially damage mi-tochondria, which can again generate oxidative stress. Our results also showed that capsaicin increased the levels of Bax and p21, p16, and decreased the expression of Bcl-2 and the altered ratio of Bax/Bcl-2 proteins could further affect the in-tegrity of mitochondria. It is well known that after cyto-chrome c is released from mitochondria into cytosol, it is fol-lowed by the sequential activation of caspase-9 and caspase-3 in various cancer cells (Shin et al., 2003; Ito et al., 2004; Jin et al., 2005; Wu et al., 2006; Zhang et al., 2008). Our earlier reports also showed that capsaicin induced apoptosis via increasing intracellular Ca21levels and leading to loss of the DCm(Wu et al., 2006). Consistent with this report, we found that capsaicin treatment increased the intracellular Ca21level and induced the loss of theDCmin SCC-4 cells.
In conclusion, our findings can be summarized as shown in Figure 6, which indicated that (1) capsaicin inducedG0/ G1 phase arrest via the decreasing of cyclin D and E, CDK2, and CDK6, but increased the levels of p21 and p16; (2) capsaicin induced apoptosis through the mitochondria-dependent and -inmitochondria-dependent pathways, and ROS plays an important role for ER stress before leading to apoptosis in SCC-4 cells.
REFERENCES
Amantini C, Mosca M, Nabissi M, Lucciarini R, Caprodossi S, Arcella A, Giangaspero F, Santoni G. 2007. Capsaicin-induced apoptosis of glioma cells is mediated by TRPV1 vanilloid receptor and requires p38 MAPK activation. J Neurochem 102:977–990.
Baek YM, Hwang HJ, Kim SW, Hwang HS, Lee SH, Kim JA, Yun JW. 2008. A comparative proteomic analysis for capsaicin-induced apoptosis between human hepatocarcinoma (HepG2) and human neuroblastoma (SK-N-SH) cells. Proteomics 8:4748–4767.
Chen JC, Lu KW, Tsai ML, Hsu SC, Kuo CL, Yang JS, Hsia TC, Yu CS, Chou ST, Kao MC, et al. 2009. Gypenosides induced G0/G1 arrest via CHk2 and apoptosis through endoplasmic reticulum stress and mitochondria-dependent pathways in human tongue cancer SCC-4 cells. Oral Oncol 45:273–283. Chen YY, Chiang SY, Lin JG, Yang JS, Ma YS, Liao CL, Lai TY,
Tang NY, Chung JG. 2010. Emodin, aloe-emodin and rhein induced DNA damage and inhibited DNA repair gene
expres-sion in SCC-4 human tongue cancer cells. Anticancer Res 30:945–951.
Costantini P, Jacotot E, Decaudin D, Kroemer G. 2000. Mito-chondrion as a novel target of anticancer chemotherapy. J Natl Cancer Inst 92:1042–1053.
Hail N Jr. 2003. Mechanisms of vanilloid-induced apoptosis. Apo-ptosis 8:251–262.
Hail N Jr, Lotan R. 2002. Examining the role of mitochondrial res-piration in vanilloid-induced apoptosis. J Natl Cancer Inst 94:1281–1292.
Huang SP, Chen JC, Wu CC, Chen CT, Tang NY, Ho YT, Lo C, Lin JP, Chung JG, Lin JG. 2009. Capsaicin-induced apoptosis in human hepatoma HepG2 cells. Anticancer Res 29:165–174. Ichimiya Y, Fuwa N, Kamata M, Kodaira T, Furutani K,
Tachi-bana H, Tomita N, Hidano S. 2005. Treatment results of stage I oral tongue cancer with definitive radiotherapy. Oral Oncol 41:520–525.
Ito K, Nakazato T, Yamato K, Miyakawa Y, Yamada T, Hozumi N, Segawa K, Ikeda Y, Kizaki M. 2004. Induction of apoptosis in leu-kemic cells by homovanillic acid derivative, capsaicin, through oxidative stress: Implication of phosphorylation of p53 at Ser-15 residue by reactive oxygen species. Cancer Res 64:1071–1078. Ji BC, Hsu WH, Yang JS, Hsia TC, Lu CC, Chiang JH, Yang JL,
Lin CH, Lin JJ, Suen LJ, et al. 2009. Gallic acid induces apo-ptosis via caspase-3 and mitochondrion-dependent pathways in vitro and suppresses lung xenograft tumor growth in vivo. J Agric Food Chem 57:7596–7604.
Jin HW, Ichikawa H, Fujita M, Yamaai T, Mukae K, Nomura K, Sugimoto T. 2005. Involvement of caspase cascade in capsai-cin-induced apoptosis of dorsal root ganglion neurons. Brain Res 1056:139–144.
Kuo TC, Yang JS, Lin MW, Hsu SC, Lin JJ, Lin HJ, Hsia TC, Liao CL, Yang MD, Fan MJ, et al. 2009. Emodin has cytotoxic and protective effects in rat C6 glioma cells: Roles of Mdr1a and nuclear factor kappaB in cell survival. J Pharmacol Exp Ther 330:736–744.
Lee YS, Kang YS, Lee JS, Nicolova S, Kim JA. 2004. Involve-ment of NADPH oxidase-mediated generation of reactive oxy-gen species in the apototic cell death by capsaicin in HepG2 human hepatoma cells. Free Radic Res 38:405–412.
Lin SY, Lai WW, Ho CC, Yu FS, Chen GW, Yang JS, Liu KC, Lin ML, Wu PP, Fan MJ, et al. 2009. Emodin induces apoptosis of human tongue squamous cancer SCC-4 cells through reactive oxygen species and mitochondria-dependent pathways. Anti-cancer Res 29:327–335.
Lopez-Carrillo L, Hernandez Avila M, Dubrow R. 1994. Chili pepper consumption and gastric cancer in Mexico: A case-con-trol study. Am J Epidemiol 139:263–271.
Lu HF, Wang HL, Chuang YY, Tang YJ, Yang JS, Ma YS, Chiang JH, Lu CC, Yang JL, Lai TY, et al. 2010. Danthron induced apoptosis through mitochondria- and caspase-3-de-pendent pathways in human brain glioblastoma multiforms GBM 8401 cells. Neurochem Res 35:390–398.
Mori A, Lehmann S, O’Kelly J, Kumagai T, Desmond JC, Pervan M, McBride WH, Kizaki M, Koeffler HP. 2006. Capsaicin, a compo-nent of red peppers, inhibits the growth of androgen-independent, p53 mutant prostate cancer cells. Cancer Res 66:3222–3229.
Oh SH, Lee BH, Lim SC. 2004. Cadmium induces apoptotic cell death in WI 38 cells via caspase-dependent Bid cleavage and calpain-mediated mitochondrial Bax cleavage by Bcl-2-inde-pendent pathway. Biochem Pharmacol 68:1845–1855.
Otera H, Ohsakaya S, Nagaura Z, Ishihara N, Mihara K. 2005. Export of mitochondrial AIF in response to proapoptotic stimuli depends on processing at the intermembrane space. EMBO J 24:1375–1386.
Pintos J, Black MJ, Sadeghi N, Ghadirian P, Zeitouni AG, Viscidi RP, Herrero R, Coutlee F, Franco EL. 2008. Human papilloma-virus infection and oral cancer: A case-control study in Mon-treal, Canada. Oral Oncol 44:242–250.
Sano D, Myers JN. 2007. Metastasis of squamous cell carcinoma of the oral tongue. Cancer Metastasis Rev 26:645–662. Shin CY, Shin J, Kim BM, Wang MH, Jang JH, Surh YJ, Oh U.
2003. Essential role of mitochondrial permeability transition in vanilloid receptor 1-dependent cell death of sensory neurons. Mol Cell Neurosci 24:57–68.
Susin SA, Lorenzo HK, Zamzami N, Marzo I, Snow BE, Brothers GM, Mangion J, Jacotot E, Costantini P, Loeffler M, et al. 1999. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature 397:441–446.
Tan TW, Tsai HR, Lu HF, Lin HL, Tsou MF, Lin YT, Tsai HY, Chen YF, Chung JG. 2006. Curcumin-induced cell cycle arrest and apoptosis in human acute promyelocytic leukemia HL-60
cells via MMP changes and caspase-3 activation. Anticancer Res B 26:4361–4371.
Vahsen N, Cande C, Briere JJ, Benit P, Joza N, Larochette N, Mastroberardino PG, Pequignot MO, Casares N, Lazar V, et al. 2004. AIF deficiency compromises oxidative phosphorylation. EMBO J 23:4679–4689.
Wu CC, Lin JP, Yang JS, Chou ST, Chen SC, Lin YT, Lin HL, Chung JG. 2006. Capsaicin induced cell cycle arrest and apoptosis in human esophagus epidermoid carcinoma CE 81T/VGH cells through the elevation of intracellular reactive oxygen species and Ca21productions and caspase-3 activation. Mutat Res 601:71–82. Yang JS, Chen GW, Hsia TC, Ho HC, Ho CC, Lin MW, Lin SS, Yeh RD, Ip SW, Lu HF, et al. 2009. Diallyl disulfide induces apoptosis in human colon cancer cell line (COLO 205) through the induction of reactive oxygen species, endoplasmic reticulum stress, caspases casade and mitochondrial-dependent pathways. Food Chem Toxicol 47:171–179.
Ye H, Cande C, Stephanou NC, Jiang S, Gurbuxani S, Larochette N, Daugas E, Garrido C, Kroemer G, Wu H. 2002. DNA bind-ing is required for the apoptogenic action of apoptosis inducbind-ing factor. Nat Struct Biol 9:680–684.
Zhang R, Humphreys I, Sahu RP, Shi Y, Srivastava SK. 2008. In vitro and in vivo induction of apoptosis by capsaicin in pancre-atic cancer cells is mediated through ROS generation and mito-chondrial death pathway. Apoptosis 13:1465–1478.