Elsevier Scientific Publishers Ireland Ltd.
S E N E S C E N C E O F R I C E L E A V E S . XX. C H A N G E S O F P R O T O N S E C R E T I O N D U R I N G
S E N E S C E N C E
CHIEN TEH CHEN0 I R E N E T. CHOU and CHING HUEI KAO*
Department of Agronomy, National Taiwan University, Ta~pe~ Taiwan (Repubhc of China~
(Received December 20th, 1988) (Revision received J u n e 29th, 1989) (Accepted A u g u s t 31st, 1989)
The relation between H ÷ secretion and senescence of detached rice leaves was investigated. The decrease of H* secretion measured in KCl-mannitol medium by detached rice leaves preceded the commencement of senescence under light or dark condition. Light, which r e t a r d e d senescence of detached leaves, slowed the loss of H* secretion and the decrease of cell sap pH. Fusicoccin promoted H ÷ secretion and r e t a r d e d senescence of detached leaves. Vanadate, which was found to decrease cell sap pH, significantly accelerated senescence of detached leaves under light and dark conditions. Isobutyric acid applied at pH 5.5 caused acidification of cell sap and acceleration of senescence as well. In contrast, isobutyric acid applied at pH 7.0 and citric acid at pH 5.5 did not r e s u l t in acidification of the cell sap and had no effect on senescence. Benzyladenine- and abscisic acid- p r e t r e a t e d leaf segments had higher and lower H ÷ secretion activity, respectively, than w a t e r - p r e t r e a t e d ones. In conclusion, H ° secretion activity of detached leaves may play a regulatory role in senescence.
Key words: cell sap pH; leaf senescence; Oryza satw~" proton secretion
Introduction
Exchange of H ÷ b e t w e e n plant cells and their
surroundings is associated with a range of met-
abolic processes, and especially with solute
transport. Several lines of evidence indicate
that auxin-induced cell elongation of coleoptiles
and etiolated seedling stems is mediated by
proton secretion [1]. H ÷ secretion has also been
shown to play an important role in regulating
light-induced cell expansion in bean leaves [2].
Relatively little attention has been paid to the
role of H ÷ secretion in regulating leaf senesc-
ence. Gepstein [3] reported that cessation of H ÷
secretion parallelled the acceleration of oat leaf
senescence. However, this is the only r e p o r t
relating H ÷ secretion to leaf senescence so far.
It is not known w h e t h e r other leaf s y s t e m s
show a similar relationship. For this reason we
investigated this relationship in rice leaves.
M e t h o d s and Materials
Rice
(Oryza sativa
L. cv. Taichung Native 1)
seedlings were grown as previously described
[4]. Rice seedlings were grown in a greenhouse
with natural light at 30°C day/25°C night.
The apical 3-cm segments excised from the third
leaves of 12-day-old seedlings were used. A
group of 10 segments were floated in a Petri
dish containing distilled w a t e r or t e s t solution.
All t e s t solutions and distilled w a t e r were
adjusted to pH 5.5. Incubation was carried out
at 27°C in darkness or in light (20 W m -2)
provided by a Grolux fluorescent lamp. Leaf
samples
were
collected
to
determine
chlorophyll level and H ÷ secretion at 1-day
intervals or at various other times.
Chlorophyll was extracted and determined
as described before [5]. Chlorophyll was
expressed as A~5 per 10 segments after extrac-
tion in 10 ml of 80% (v/v) ethanol.
The procedure for measuring H ÷ secretion
by detached rice leaves was, in general, based
on that of Gepstein [3]. The leaf segments were
cut into 0.2-cm pieces. However, we did not
scrub leaf segments before cutting them into
small pieces; scrubbing may result in wounding
0168-9452/90/$03.50 © 1990 Elsevier Scientific Publishers Ireland Ltd.effects [7]. F o u r h u n d r e d and fifty 0.2-cm seg-
ments (weighing about 150 mg) w e r e incubated
in 5 ml of a solution containing 250 mM manni-
tol and I mM KC1 at 27°C and w e r e shaken at a
speed of 70 cycles/min. Unless otherwise indi-
cated, H ÷ secretion was m e a s u r e d u n d e r light
(20 W m -2)
provided by a Grolux fluorescent
lamp. The secretion of protons from leaf seg-
ments was e x p r e s s e d as the change in pH:
- A pH =
- ( f i n a l pH -
initial pH) in the
medium during a 4-h incubation. The initial pH
was adjusted with 1 mM NaOH to be b e t w e e n
pH 6.5 and 6.8.
F o r the m e a s u r e m e n t of the cell sap pH, leaf
segments were t r a n s f e r r e d at t h e end of treat-
ments directly into syringes and were frozen in
liquid nitrogen. A f t e r thawing the cell sap was
pressed out from the syringe and the pH was
measured.
All e x p e r i m e n t s w e r e r e p e a t e d one or more
times with similar results. The data r e p o r t e d
here are from single e x p e r i m e n t s .
Results
To investigate the relation b e t w e e n H ÷
secretion and senescence of detached rice
leaves, the H ÷ secretion m e a s u r e d should be
shown to be an active process r a t h e r than a
t r a n s i e n t passive exchange of H+/K ÷ and not to
be due to cut or damaged cells. Our previous
work [8] d e m o n s t r a t e d t h a t the H ÷ secretion
m e a s u r e d was indeed an active process. The
reversibility of pH changes e x t e r n a l to leaf
segments shifted from light to darkness, or
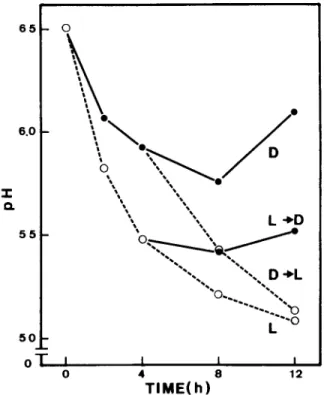
vice-versa, as is shown in Fig. 1. This indicates
t h a t the H ÷ secretion m e a s u r e d is unlikely to be
due to cut or damaged cells.
The senescene of rice leaves was followed by
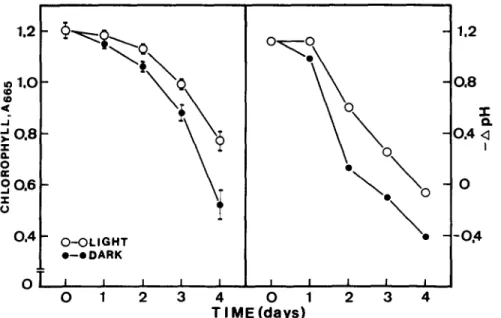
measuring the decrease of chlorophyll. F i g u r e 2
shows the time courses of chlorophyll level and
H ÷ secretion of detached leaves which had been
floating on distilled w a t e r in light or darkness
for various times. The decrease of chlorophyll
level in darkness and light was significantly
evident at 2 and 3 days, respectively, a f t e r leaf
detachment. It is also clear from Fig. 2 t h a t
light was effective in r e t a r d i n g senescence of
"I"
6 5 - 0
6 . 0 -•
/ ~ ~ D
°
e~L ÷D
5 5
oT
,
~
i
,
0
4
8
12
TIMElh)
Fig. 1. Changes of pH in external medium by rme leaf segments in continuous darkness (D), continuous light (L), and after t r a n s f e r from light to dark (L ~ D) or from dark to light (D ~ L). Leaf segments (0.2 cm) were incubated in standard medium (250 mM mannitol and 1 mM KC1).
rice leaf segments. The rates of H ÷ secretion
significantly decreased at 1 and 2 days after
incubating in darkness and light, respectively.
Light also substantially slowed the loss of H ÷
secretion.
Theoretically, the decrease in rate of H ÷
secretion m a y result in a lowering of intracellu-
lar pH. If the decrease of H ÷ secretion plays an
important
role in regulating
rice leaf
senescence, then decrease of the cell sap p H
before the c o m m e n c e m e n t of chlorophyll degra-
dation is to be expected. Significant decrease of
the cell sap p H was observed at 1 day after
incubating leaf segments in light or darkness
(Table I). It is also clear from Table I that in the
light there was less change in the cell sap pH.
It has long been recognized that cytokinins
are effective in retarding the senescence of
most, if not all,
leaves. The effect of benzyladen-
ine, a synthetic cytokinin, on senescence of
1.2 © 1,O ¢D ¢D - I
0.8
z a. 0 ES o.e
.T- U 0.4oT
O - O L I G H T e--eDARK I I I I 0 1 2 3\
I I I I I i 4 0 1 2 3 4 T I M E ( d a y s ) 1.2 0 . 8 "t" O.-0.4
< i- 0
- - 0 . 4Fig. 2. C h a n g e s of chlorophyll content and H ÷ secretion of detached rice leaves incubated in distilled water under dark and light conditions. A t 24-h intervals, detached leaves were cut into 0.2-cm pieces which w e r e then transferred and incubated in
standard m e d i u m (250 m M mannitol and 1 m M KCI) to m e a s u r e H ÷ secretion. Chlorophyll was expressed as mean ± S.E., 4 repetitions.
detached leaves is presented in Table II. It was
found that benzyladenine effectively retarded
senescence (loss of chlorophyll) in detached
leaves under both light and dark conditions.
A m o n g the known promoters of senescence,
abscisic acid has been most often studied. Gep-
stein and Thimann [9] claimed that abscisic acid
is an endogenous factor in leaf senescence.
Abscisic acid significantly promoted senesc-
ence of detached rice leaves under dark and
light conditions (Table II). If the decrease in
rate of H ÷ secretion plays an important role in
senescence of detached rice leaves, then H ÷
secretion activity of benzyladenine- and abs-
cisic acid-treated detached leaves would be
Table I. E f f e c t of light and d a r k on t h e cell sap p H of rice
leaf segments. Mean ± S.E., 4 repetitions.
Treatment Cell sap pH Initial 6.24 ± 0.03 Dark, 24 h 6.05 ± 0.01 Light, 24 h 6.18 ± 0.01
expected to be higher and lower, respectively,
than those treated with water alone. As indi-
cated in Table II, this is indeed the case.
Several lines of evidence indicate that a plaso
malemma A T P a s e acts as an electrogenic pump
in higher plants [10]. Recently, Gepstein [3]
demonstrated that vanadate, a specific inhibi-
Table II. E f f e c t of benzyladenine and abscisic acid on chlo-
rophyll content and H ÷ secretion of rice leaf segments. Rice leaf s e g m e n t s w e r e incubated in benzyladenine (10 pM) or
abscisic acid (10 ~M) in light o r d a r k n e s s for 4 days. L e a f segments were then collected to measure H ÷ secretion and
chlorophyll content. Chlorophyll was expressed as mean ± S.E., 4 repetitions.
Addition Chlorophyll - A pH (A~5/10 segments)
None, dark 0.61 -+ 0.03 - 0.20
Benzyladenine, dark 0.92 ± 0.02 0.47
Abscisic acid, dark 0.38 ± 0.03 - 0.45
None, light 0.81 ± 0.02 0.13
Benzyladenine, light 0.99 ± 0.01 0.70
o°91
°°'51-
O(~.v- I :1 I i I
0
5x1(34
163
5xlo 3
VANADATE,M
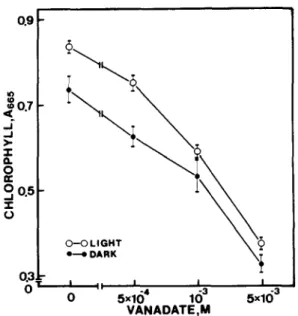
Fig. 3. Effect of different concentrations of ammonium v a n a d a t e (pH 5.5) on chlorophyll content in detached rice leaves after 4 days in the light or 3 days in darkness. The controls for light and darkness contained 0.25 mM sodium phosphate buffer, 5 mM NH,C1 and 1 mM KCI. Data points are means ± S.E., 4 repetitions.
tor of plasma membrane ATPase, blocked H *
secretion and promoted senescence of detached
oat leaves. Our previous work also showed t h a t
H ÷ secretion by rice leaf segments originated
from an ATP-driven H ÷ pump located in the
plasmalemma and t h a t vanadate significantly
Table I l L Effect of ammonium v a n a d a t e on the cell sap pH of rice leaf segments. Leaf s e g m e n t s were incubated in ammonium chloride (5 raM, pH 5.5) or ammonium v a n a d a t e (5 raM, pH 5.5) for 8 h. Mean ± S.E., 4 repetitions.
T r e a t m e n t Cell sap pH Dark NH,CI 6.09 ± 0.01 NH,VO 3 5.90 _+ 0.02 Lsght NH4CI 6.04 ± 0.03 NH4VO 3 5.94 ± 0.01
phyU content of rice leaf segments. Before measuring H ÷ secretion, freshly excised leaves (apical 3-cm segments) were cut into 0.2 cm pieces and incubated in standard medium (250 mM mannitol and 1 mM KC1) with or without fusicoecin for 4 h u n d e r light or dark condition. For chloro- phyll determination, leaf segments (3 cm) had been incubated 4 days in the absence or the presence of fusicoc- cin in light or darkness. Controls contained phosphate buffer (10 raM, pH 5.5). Chlorophyll was expressed as mean
± S.E., 4 repetitions.
T r e a t m e n t Chlorophyll - A pH (A~/10 segments)
Experiment I
Dark control 0.40 ± 0.01 0.64 Fusicoccin (10 ~uM), dark 0.58 ± 0.01 2.90
Experiment II
Light control 0.54 + 0.01 Fusicoccin (0.1 t~I), light 0.60 ± 0.01
1.06 2.02
inhibited H + secretion [8]. The effect of vanda-
date on senescence of detached rice leaves
measured as chlorophyll loss is presented in
Fig. 3. Vanadate effectively promoted senesc-
ence of detached leaves under both light and
dark conditions. Vanadate was also found to
decrease the cell sap pH of rice leaf segments
incubated in light or darkness (Table III). Curl-
ing and loss of turgor of leaf segments were not
evident even at 5 mM vanadate.
Fusicoccin is commonly used to induce H ÷
secretion via plasmalemma ATPase [11]. Fusi-
coccin showed not only promotion of H ÷ secre-
tion but also retardation of chlorophyll loss in
detached leaves under light and dark condi-
tions (Table IV).
Recently, Pesci and Beffagna [12] demon-
strated that exogenously supplied isobutyric
acid induced an acidification of the cell sap in
detached leaves of barley. Therefore, it was of
great interest to investigate the effect of isobu-
tyric on the cell sap pH and senescence of rice
leaf segments. Treatment with isobutyric acid
at pH 5.5 in darkness or light resulted in a
decrease of the cell sap pH, reaching a value
which was about 0.3 pH unit below that of
untreated leaves within 24 h (Table V). As
Table V. Effects of isobutyric acid and citric acid on the cell sap pH and chlorophyll content of rice leaf segments. Leaf seg- m e n t s were incubated in isobutyric acid (10 mM) or citric acid (10 mM) in light or darkness. Cell sap pH was measured at 24 h after incubation. Chlorophyll content was d e t e r m i n e d at 4 and 3 days in light and darkness, respectively. Mean ± S.E., 4 repetitions.
T r e a t m e n t Chlorophyll Cell sap pH
(Am/10 segments) Dark W a t e r (pH 5.5) 0.80 ± 0.08 6.22 ± 0.01 W a t e r (pH 7.0) 0.83 ± 0.05 6.25 ± 0.02 Isobutyric acid (pH 5.5) 0.53 ± 0.03 5.96 ± 0.02 Isobutyric acid {pH 7.0) 0.77 ± 0.02 6.25 ± 0.02 Citric acid (pH 5.5) 0.72 ± 0.01 6.26 ± 0.03
Light
W a t e r (pH 5.5) 0.85 + 0.01 6.26 ± 0.02 W a t e r (pH 7.01 0.82 ± 0.03 6.30 ± 0.02 Isobutyric acid (pH 5.5) 0.75 ± 0.02 5.94 ± 0.01 Isobutyrie acid {pH 7.0~ 0.81 ± 0.03 6.29 ± 0.02 Citric acid (pH 5.5) 0.87 ± 0.08 6.28 ± 0.02expected, isobutyric acid applied at pH 5.5
promoted senescence of detached rice leaves
(measured as chlorophyll loss) under both light
and dark conditions (Table V). On the contrary,
isobutyric acid applied at pH 7.0 had no effect
on the cell sap pH and senescence. Since
isobutyric acid is a weak acid with pK 4.84, the
lack of effects on the cell sap pH and senescence
by isobutyric acid applied at pH 7.0 can be
attributed to fewer molecules of isobutyric acid
penetrating the plasmalemma. When rice leaf
segments were t r e a t e d with the less permeable
citric acid (pK 1 = 3.71) at the same concentra-
tion and pH as isobutyric acid (10 raM, pH 5.5) in
light or darkness, the cell sap pH was not
decreased and senescence was not promoted
(Table V).
Discussion
Since the decrease in rate of H ÷ secretion in
rice leaves is rapid following excision, one may
argue that the H ÷ secretion is probably the
result of permeability change. This possibility
is ruled out by our previous finding that solute
leakage increased only after the commence-
ment of chlorophyll loss [13], during which time
leaf segments had much lower H + secretion
activity. No difference of respiration rate at an
early stage of incubation
was observed
b e t w e e n light- and dark-treated leaf segments
[6]. It seems unlikely that the inhibition of the
decrease in rate of H ÷ secretion by light is due
to respiratory C02 dissolved in the external
solution.
Gepstein [3] suggested that H ÷ secretion
played a regulatory role in oat leaf senescence.
Our results, in general, support this suggestion.
This conclusion is based on the observations
that: (a) the decrease in rate of H ÷ secretion
preceded the commencement of senescence of
detached
leaves
under
light
and
dark
conditions; (b) the decrease of the cell sap pH
occurred at 24 h after incubating leaf segments
in light or darkness, which was prior to chloro-
phyll loss; (c) light, which r e t a r d e d the
senescence of detached leaves, inhibited the
decrease of H ÷ secretion and the decrease of the
cell sap pH; (d) benzyladenine-treated detached
leaves had higher H ÷ secretion activity than
water-treated leaves, whereas abscisic acid-
treated detached leaves had lower H ÷ secretion
secretion, retarded senescence of detached
leaves under both light and dark conditions; (f)
vanadate, which is known to inhibit H ÷ secre-
tion, decreased the cell sap pH and accelerated
senescence of detached leaves in light or dark-
ness; (g) isobutyric acid applied at pH 5.5 acidi-
fied the cell sap and accelerated senescence of
detached leaves under both light and dark con-
ditions; (h) isobutyric acid applied at pH 7.0 and
citric acid at pH 5.5 had no effect on the cell sap
pH and senescence of detached leaves in light
or darkness.
Gepstein [3] speculated that the role of H ÷
pumping in non-expanding cells might be pri-
marily to prevent the acidosis of the cytoplasm
rather than to acidify the cell wall. According
to this view, this mechanism may retard the
process of senscence by decreasing the activity
of acid hydrolases which are known to be
involved in leaf senescence [14]. This mecha-
nism also explains our early findings that
proteinase and carboxypeptidase, which have
low pH optima, showed increased activities
during senescence of rice leaves [15].
Previously we have reported that proline
accumulated in detached rice leaves during
senescence [16]. Recent findings by GSring and
Plescher [17] and Pesci and Beffagna [12,18]
suggested that a decrease of intracellular pH
caused stress-, abscisic acid- and isobutyric
acid-induced proline accumulation in wheat
coleoptiles and barley leaves. Based on the
results reproted in this investigation, it seems
that proline accumulation during senescence of
detached rice leaves is most likely due to
decreased H + secretion.
Acknowledgement
This research was supported by a grant from
the National Science Council of the Republic of
China.
1 R.E. Cleland and D.L. Rayle, Auxin, H÷-excretion and cell elongation. Bot. Mag. Tokyo (Special Issue), 1 (1978) 125-139.
2 E. Von Volkenburgh and R.E. Cleland, Proton excretion and cell expansion in bean leaves. Planta, 148 (1980) 273- 278.
3 S. Gepstein, Light-induced H" secretlon and the rela- tion to senescence of oat leaves. Plant Physiol., 70 (1982) 1120-1124.
4 W.P. Hurng, T.L. Lin, S.S. Ren, J.C. Chen, R.R. Chen and C.H. Kao, Senescence of rice leaves. XVIII. Changes of stomatal aperture during senescence. Plant Cell Physiol., 29 (1988) 2 7 - 31.
5 C.H. Kao, Senescence of rice leaves. IV. Influence of benzyladenme on chlorophyll degradation. Plant Cell Physiol., 21 (1980) 1255-- 1262.
6 C.H. Kao, Senescence of rme leaves. XIV. Changes of respiration during senescence of detached leaves. Bot. Bull. Acad. Sin., 26 {1985) 21 - 30.
7 G. Gimdhar and K.V. Thlmann, Interaction between senescence and wounding m oat leaves. Plant Physiol.,
78 (1985) 29--33.
8 C.T. Chen and C.H. Kao, Proton secretion by rme
leaves. Bot. Bull. Acad. Sin., 29 (1988)315-320.
9 S. Gepstem and K.V. Thimann, Changes in the abscisic
acid content of oat leaves during senescence. Proc.
Natl. Acad. SCL U.S.A., 77 (1980) 2050 -- 2053.
10 R.M. Spanswick, Electrogenic ion pumps. Annu. Rev. Plant Physiol., 32 (1981) 267 -- 289.
11 E. Marr4, Fusicoccin; a tool m plant physiology. Annu. Rev. Plant Phymol., 32 (1981) 2 6 7 - 289.
12 P. Pesci and N. Beffagna, Effects of weak acids on pro-
line accumulation in barley leaves: a comparison between abscisic acid and isobutyric acid. Plant Cell
Environ., 8 (1985) 129 - 133.
13 Y.Y. Shyr and C.H. Kao, Senescence of rice leaves. XV. Solute leakage and inorganic phosphate uptake. Bot. Bull. Acad. Sin., 26 (1985) 171 -- 178.
14 K.V. Thimann, Senescence. Bot. Mag. Tokyo (Special Issue), 1 (1978) 17--26.
15 S.H. Cheng and C.H. Kao, The role of proteolyhc
enzymes in protein degradation during senescence of
rice leaves. Physiol. Plant., 62 (1984) 231 --237. 16 C.Y. Wang, S.H. Cheng and C.H. Kao, Senescence of
rme leaves, VII. Proline accumulation m senescing
excised leaves. Plant Physiol., 69 (1982) 1348 - 1349.
17 H. Goring and F. Plescher, Proline accumulahon
induced by weak acids and IAA in coleoptile of wheat seedlings. Biol. Plant., 28 (1986) 401 -- 406.
18 P. Pesci and N. Beffagna, Inhibiting effect of fusicoccm on abscmic acid-reduced proline accumulation m barley