Inhibitory effects of maslinic acid upon human esophagus, stomach and
pancreatic cancer cells
Sheng-lei Yana, Mei-chin Yinb,c,*
aDivision of Gastroenterology, Department of Internal Medicine, Chang Bing Show-Chwan Memorial Hospital, Changhua County, Taiwan
bDepartment of Nutrition, China Medical University, Taichung City, Taiwan cDepartment of Health and Nutrition Biotechnology, Asia University, Taichung City,
Taiwan
running title: anti-cancer effects of maslinic acid
*To whom correspondence should be addressed: Dr. Mei-chin Yin, Professor, Department of Nutrition, China Medical University, 91, Hsueh-shih Rd., Taichung City, Taiwan TEL: 886-4-22053366 ext. 7510, FAX: 886-4-22062891
Email: [email protected] 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
Abstract
Apoptotic, anti-invasive and anti-migratory effects of maslinic acid (MA) at 4, 8, or 16 M in human esophageal squamous cancer cell line, OE33; gastric cancer cell line, SGC-7901; and pancreatic cancer cell line, Panc-1, were examined. MA treatments inhibited OE33 and SGC-7901 cells growth at 21-66% and 32-75%, respectively; but lowered Panc-1 viability at 13-27% only. MA treatments increased cleaved caspase-3 and Bax expression, and raised caspase-3 and caspase-8 activities in OE33 and SGC-7901 cells. MA treatments also increased DNA fragmentation and decreased reactive oxygen species production in these two cell lines. MA treatments declined invasion and migration in OE33 and SGC-7901 cells, and lowered vascular endothelial growth factor and transforming growth factor-beta1 levels in these cells. MA suppressed hypoxia-inducible factor-1alpha, matrix metalloproteinase (MMP)-2 and MMP-9 expression in OE33 and SGC-7901 cells. These findings indicated that this triterpene was a potent agent against esophagus and stomach cancers.
Keywords: Maslinic acid; GI tract cancer cells; Apoptosis; Invasion; Migration
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
1. Introduction
Esophagus, stomach, pancreatic, liver and colon cancers are common gastrointestinal (GI) tract cancers (Khamly, Jefford, Michael, & Zalcberg, 2006). The development and application of agent with inhibitory effects upon these GI tract cancer cells benefits the prevention and/or therapy for these cancers.
Maslinic acid (MA) is a pentacyclic triterpenic acid naturally occurring in many herbs and plant foods such as glossy privet fruit (Ligustrum lucidum Ait.), hawthorn fruit (Crataegi Pinnatifidae Fructus) and olive (Cui et al., 2006; Yin, Lin, Mong, & Lin, 2012). It is reported that this triterpene could inhibit the proliferation of human colon cancer cell lines, HT-29 and Caco-2 (Reyes, Centelles, Lupiáñez, & Cascante, 2006; Juan, Planas, Ruiz-Gutierrez, Daniel, & Wenzel, 2008). Our previous study found that MA at 2-16 M declined invasion and migration in three hepatic cancer cell lines via suppressing mRNA expression of angiogenic factors including hypoxia-inducible factor (HIF)-1alpha, vascular endothelial growth factor (VEGF) and urokinase-type plasminogen activator (Lin, Huang, Mong, Chan, & Yin, 2011). These previous studies implied that this triterpene was a potent cytotoxic agent for GI tract cancers. However, it is unknown that MA could retard the growth of other GI tract cancer cells such as esophagus, stomach or pancreatic cancer cells. Human esophageal squamous cancer cell line, OE33; gastric cancer cell line, SGC-7901; and pancreatic cancer cell line, Panc-1, have been widely used for anti-cancer studies to investigate the apoptotic effects and possible mechanisms of certain compounds (Li et al., 2012; Milano et al., 2013; Zhang et al., 2013). In our present study, these cancer cell lines were used to examine the apoptotic effects, anti-invasive and anti-migratory activities of MA at various doses.
Cancer cells apoptosis could be induced by increasing the expression of apoptotic factors such as Bax and cleaved caspase-3, and/or decreasing the expression of anti-1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
apoptotic factors such as Bcl-2 (Manikandan, Murugan, Priyadarsini, Vinothini, & Nagini, 2010; Prasad, Vaid, & Katiyar, 2012). In addition, activated caspase cascade is another important pathway to promote cell apoptosis (Frejlich et al., 2013). So far, less information is available regarding the influence of MA upon caspase activity, expression of apoptotic or anti-apoptotic molecules in these cancer cell lines. On the other hand, VEGF, transforming growth factor (TGF)-beta1, matrix metalloproteinase (MMP)-2 and MMP-9 are involved in GI tract cancer cell adhesion, invasion and migration, which contribute to cancer metastasis (Augoff et al., 2009; Xue et al., 2013). HIF-1alpha regulates the essential adaptive responses of cancer cells to hypoxia; and reactive oxygen species (ROS) could be generated by mitochondria under hypoxic condition (Pugh & Ratcliffe, 2003). Increased HIF-1alpha expression and ROS overproduction favor the metastasis of gastric and pancreatic cancers (Park et al., 2003; Shimojo et al., 2013). Therefore, if MA could mediate these factors such as MMPs, HIF-1alpha or TGF-beta1 in OE33, SGC-7901 or Panc-1 cells, its anti-invasive or anti-migratory actions upon these cancer cells could be explained.
2. Materials and methods 2.1. Chemicals
MA (98%) was purchased from Aldrich Chemical Co. (Milwaukee, WI, USA). Medium, plates, antibiotics and chemicals used for cell culture were purchased from Difco Laboratory (Detroit, MI, USA). All chemicals used in these measurements were of the highest purity commercially available.
2.2. Cell culture
OE33, SGC-7901 and Panc-1 cell lines were obtained from American Type Culture Collection (ATCC, Rockville, MD, USA). Cells were cultured in RPMI 1640 medium, 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
containing 10% fetal bovine serum (FBS), 100 units/mL of penicillin and 100 units/mL of streptomycin (pH 7.4) at 37°C in 5% CO2. The culture medium was changed every three days, and cells were subcultured once a week. A phosphate buffer saline (PBS, pH 7.2) was added to adjust the cell number to 105/mL for various experiments and analyses. 2.3. Experimental design
Stock solution of MA was prepared in dimethyl sulfoxide (DMSO) and diluted with medium. An equal volume of DMSO (final concentration <0.5%) was added to the controls. Test cells (105/mL) were treated with MA at 4, 8 or 16 M for 48 hr at 37°C. Control group contained no MA.
2.4. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
MTT assay was performed to examine cell viability. Briefly, test cells were incubated with 0.25 mg MTT/mL for 3 hr at 37°C. The amount of MTT formazan product was determined by measuring absorbance at 570 nm (630 nm as a reference) using a microplate reader (Bio-Rad, Hercules, CA, USA). Cell viability was expressed as a percent of control group.
2.5. Measurement of DNA fragmentation
Cell death detection ELISA plus kit (Roche Molecular Biochemicals, Mannheim, Germany) was used to quantify DNA fragmentation. After incubation with MA at various concentrations, cells were lysed for 30 min at room temperature and followed by centrifugation at 200 xg for 10 min. Then, 20 L supernatant was transferred onto the streptavidin-coated plate, and 80 L freshly prepared immunoreagent was added to each well and incubated for 2 hr at room temperature. After washing with PBS, the substrate solution was added and incubated for 15 min. The absorbance at 405 nm (reference wavelength 490 nm) was measured using a microplate reader. DNA fragmentation was expressed as the enrichment factor using the following equation:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
enrichment factor = (absorbance of the sample) / (absorbance of the control), sample: cells treated with MA; control: cells without MA treatment.
2.6. Measurement of caspases activity
Activity of caspase-3 and -8 was detected by using fluorometric assay kits (Upstate, Lake Placid, NY, USA) according to the manufacturer’s protocol. In brief, control or treated cells were lysed in 50 mL cold lysis buffer and incubated in ice for 10 min. Fifty L cell lysates were mixed with 50 mL reaction buffer and 5 mL fluorogenic substrates specific for caspase-3 or -8 in a 96-well microplate. After incubation at 37°C for 1 hr, fluorescent activity was measured using a fluorophotometer with excitation at 400 nm and emission at 505 nm. Data were expressed as percentage of the control, and the control group was designated as 100%.
2.7. Analysis of VEGF, TGF-beta1 and ROS levels
Cells were pre-incubated with MA at various concentrations for 48 hr. After washing by PBS, samples were incubated with 10 ng/mL TNF-alpha for 18 hr. VEGF and TGF-beta1 levels in cell culture supernatant were determined using ELISA kits (R&D Systems, Minneapolis, MN, USA). The dye DCFH2-DA was used to measure ROS level. Briefly, cells were washed and suspended in RPMI 1640 medium. After incubating with 50 M dye for 30 min and washing with PBS, the cell suspensions were centrifuged at 412 xg for 10 min. Then, the medium were removed and cells were dissolved with 1% Triton X-100. Fluorescence changes were measured at an excitation wavelength of 485 nm and an emission wavelength of 530 nm using a fluorescence microplate reader.
2.8. Western blot analysis
Cells were suspended in lysis buffer, and protein content was determined by Bio-Rad protein assay reagent (Bio-Rad Laboratories Inc. Hercules, CA, USA). Sample at 40 μg protein was applied to 10% SDS-polyacrylamide gel electrophoresis, and transferred to a 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
nitrocellulose membrane (Millipore, Bedford, MA, USA) for 1 hr. After blocking with a solution containing 5% nonfat milk for 1 hr, membrane was incubated with mouse cleaved caspase-3 (1:1000), Bcl-2 (1:2000), Bax (1:1000), HIF-1alpha, anti-MMP-2, anti-MMP-9 (1:1000) monoclonal antibody (Boehringer-Mannheim, Indianapolis, IN, USA) at 4ºC overnight, and followed by reacting with horseradish peroxidase conjugated antibody for 2 hr. The detected bands were quantified by Scion Image analysis software (Scion Corp., Frederick, MD, USA), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a loading control. The blot was imaged by autoradiography, and quantified by densitometric analysis. Results were normalized to GAPDH, and given as arbitrary units (AU).
2.9. Invasion and migration assay
Cell invasion and migration were measured in transwell chambers by fibronectin- and matrigel-coated polycarbonate filters, respectively. In brief, cells (105/100 L) were seeded into the upper chamber in 200 L of serum-free medium containing MA at various doses; the lower chamber was filled with 0.66 mL of RPMI 1640 media containing 10% of FBS as a chemoattractant. After 6 hr incubation for migration assay or 16 hr incubation for invasion assay at 37ºC, the cells on the upper surface of the filter were removed by a cotton swab. The migrated or invaded cells to the lower surface of the filter were stained with 0.2% crystal violet in 10% ethanol. Four independent fields of invasive or migratory cells per well were photographed under the microscope to determine the cell numbers. Data were calculated as percentage of the control group.
2.10. Statistical analysis
The effect of each treatment was analyzed from eight different preparations (n=8). Data were reported as means standard deviation (SD), and subjected to analysis of variance (ANOVA). Differences among means were determined by the Least Significance 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Difference Test with significance defined at P<0.05.
3. Results
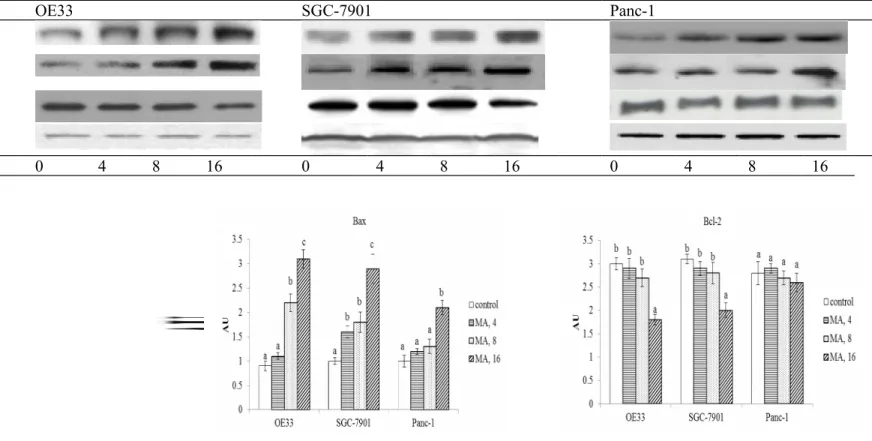
MA treatments at three doses inhibited the growth of OE33 and SGC-7901 at 21-66% and 32-75%, respectively (Figure 1, P<0.05). However, MA only at 8 and 16 M decreased Panc-1 viability in the range of 13-27%. As shown in Figure 2, MA treatments increased cleaved caspase-3 expression in OE33, SGC-7901 and Panc-1 cells; and dose-dependent manner was presented in OE33 and SGC-7901 cells (P<0.05). MA at 8 and 16 M raised Bax expression in OE33 cells; and at 4, 8 and 16 M raised Bax expression in SGC-7901 cells (P<0.05). Bax expression in Panc-1 cells was increased only by 16 M MA treatment (P<0.05). MA only at 16 M decreased Bcl-2 expression in OE33 and SGC-7901 cells (P<0.05); and MA did not affect Bcl-2 expression in Panc-1 cells (P>0.05). MA treatments dose-dependently raised caspase-3 and caspase-8 activities in OE33 and SGC-7901 cells (Figure 3, P<0.05). MA only at 8 and 16 M increased caspase-3 and caspase-8 activities in Panc-1 cell line (P<0.05). As shown in Table 1, MA treatments dose-dependently increased DNA fragmentation and decreased ROS production in OE33 and SGC-7901 cells (P<0.05). MA at three doses lowered ROS formation in Panc-1 cells (P<0.05); but only at 8 and 16 M increased DNA fragmentation in this cell line (P<0.05).
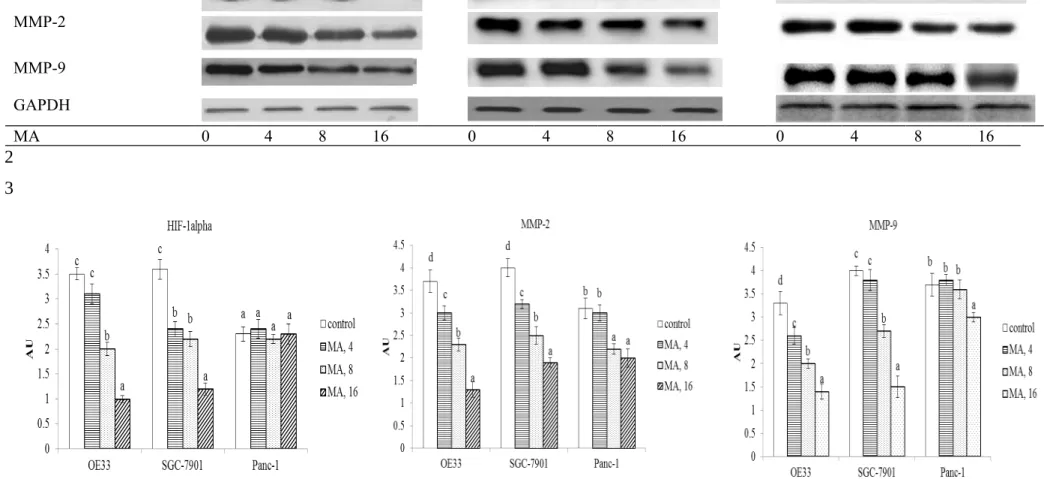
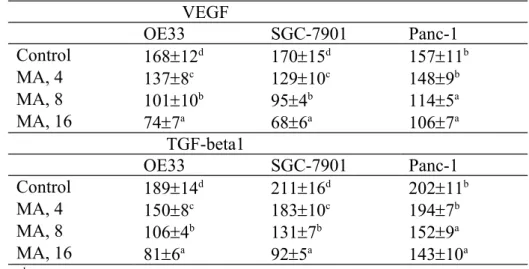
MA at three doses reduced cell invasion and migration in OE33 and SGC-7901 cells; and only at 8 and 16 M declined cell invasion and migration in Panc-1 cells (Figure 4, P<0.05). MA dose-dependently lowered VEGF and TGF-beta1 levels in OE33 and SGC-7901 cells (Table 2, P<0.05); and only at 8 and 16 M decreased VEGF and TGF-beta1 levels in Panc-1 cells (P<0.05). As shown in Figure 5, MA at 8 and 16 M down-regulated 42.9-71.4% HIF-1alpha expression in OE33 cells; and at three doses suppressed 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
33.1-67.5% 1alpha expression in SGC-7901 cells (P<0.05). MA did not affect HIF-1alpha expression in Panc-1 cells. MA dose-dependently lowered MMP-2 expression at 18.9-62.2% and 20.1-51.6% in OE33 and SGC-7901 cells, respectively; and only at 8 and 16 M reduced 27.2-39.4% MMP-2 expression in Panc-1 cells (P<0.05). MA also dose-dependently lowered 24.3-54.5% MMP-9 expression in OE33 cells; and at 8 and 16 M reduced 32.5-62.7% MMP-9 expression in SGC-7901 cells (P<0.05). MMP-9 expression in Panc-1 cells was decreased 23.4% by 16 M MA treatment only (P<0.05).
4. Discussion
The cytotoxic effects of MA against colon and liver cancer cells have been reported (Juan, Planas, Ruiz-Gutierrez, Daniel, & Wenzel, 2008; Lin, Huang, Mong, Chan, & Yin, 2011). Our present study further found that this triterpene effectively inhibited the growth of OE33 and SGC-7901 cells, esophagus and stomach cancer cells. Our data revealed that MA suppressed protein expression of cleaved caspase-3, Bax, HIF-1alpha and MMPs; increased DNA fragmentation; lowered ROS, VEGF and TGF-beta1 production in OE33 and SGC-7901 cells, which consequently caused apoptosis and declined invasion and migration for these cancer cells. These findings supported that MA was a potent agent against esophagus and stomach cancers. Although MA also decreased viability and ROS level in Panc-1 cells, its impact upon several examined factors in this cell line was mild. It seems that MA was a weak agent against pancreatic cancer.
Apoptosis occurs through the death receptor pathway and/or mitochondrial pathway. We found MA at test doses markedly up-regulated cleaved caspase-3 and Bax expression in OE33 and SGC-7901 cells; but only at high dose declined Bcl-2 expression in these two cell lines. Obviously, MA caused apoptosis in these cancer cells through mitochondrial pathway; and its apoptotic action was mainly due to its mediation upon apoptotic factors 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
rather than anti-apoptotic factor. Caspases include two subfamilies: upstream initiator caspases such as caspases-8, which are involved in regulatory events; and downstream effector caspases such as caspases-3, which are directly response to the change in cell morphological events (Frejlich et al., 2013). Our present study found that MA treatments effectively enhanced the activity of caspase-8, an upstream initiator, and caspase-3, a downstream effector. These findings indicated that MA was able to activate both upstream and downstream caspases in test cancer cell lines, which in turn contributed to the observed apoptosis in these cancer cell lines. In addition, caspase-8 and caspase-3 are also responsible for death signals transduction in death receptor pathway (Hu et al., 2013). Thus, MA induced apoptosis in these cancer cells might be partially through the death receptor pathway.
DNA fragmentation is a marker of cell death. Our data regarding cell viability and DNA fragmentation in OE33 and SGC-7901 cells were consistent, and these results suggest that MA could penetrate into these cancer cells and destroy plasma integrity, which consequently led to cell apoptosis. It is known that ROS overproduction enhances oxidative stress and stimulates VGEF production, which promotes esophagus or stomach cancer progression (Park et al., 2003; Hong et al., 2011; Kajdaniuk, Marek, Folty, & Kos-Kudła, 2011). MA is a compound with anti-oxidative activity (Allouche, Beltrán, Gaforio, Uceda, & Mesa, 2010). It seems reasonable to observe lower ROS levels in MA-treated cancer cells. Thus, the retarded growth and metastasis in MA-treated OE33 and SGC-7901 cells could be partially ascribed to this agent decrease oxidative stress in those cells. However, it is interesting to find that MA at test doses also attenuated oxidative stress in Panc-1 cells, but this triterpene exhibited mild inhibitory effects toward this cancer cell line. It is possible that oxidative stress was not a top priority upon Panc-1 cell growth or invasion. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
HIF-1alpha is a crucial activator responsible for gastro-esophageal cancer progression because it regulates the essential adaptive process for cancer cells to hypoxia, the major pathophysiological condition (Griffiths et al., 2007). The suppressive effects of MA upon HIF-1alpha expression in OE33 and SGC-7901 cells suggested that this triterpene could weaken the adaptability of these cells to hypoxic microenvironment, which in turn diminished the survival and metastasis of these cells. Furthermore, HIF-1alpha is a transcriptional factor to enhance VEGF expression, and promote tumor neovascularization in GI tract cancers (Song et al., 2009; Zhu et al., 2011). Since MA treatments down-regulated HIF-1alpha expression in OE33 and SGC-7901 cells, the lower VEGF generation in these two cell lines could be explained. In addition, TGF-beta1 overproduction promotes angiogenesis and metastasis of esophagus and stomach cancer cells (Noma et al., 2008; Fuyuhiro et al., 2011). In our present study, MA treatments also decreased TGF-beta1 formation in OE33 and SGC-7901 cells, which consequently declined the ability of these cells to invade and migrate. These findings revealed that MA could provide anti-angiogenic, anti-fibrogenic and anti-metastatic actions in those cancer cells through suppressing HIF-1alpha, VEGF and TGF-beta1. Matrix metalloproteinases are endopeptidases, and able to degrade extracellular matrix components, which allows cancer cells to access the vasculature and lymphatic systems. MMP-2 and MMP-9 have attracted more attention because both could degrade type IV collagen, the basic component of the basement membrane (Hamano et al., 2003). Increased expression of MMP-2 and MMP-9 in esophagus and stomach cancer cells has been reported (Zheng et al., 2006; Groblewska, Siewko, Mroczko, & Szmitkowski, 2012). Our present study found that MA treatments down-regulated MMP-2 and MMP-9 expression in OE33 and SGC-7901 cells, which subsequently restricted the process of invasion and migration. These findings indicated that MA declined invasion and migration of these two cancer cell lines via lowering 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
available MMP-2 and MMP-9. Although MA treatments at 8 and 16 M also limited MMPs expression and decreased TGF-beta1 formation in Panc-1 cells, these treatments failed to affect HIF-1alpha expression in this cell line, and finally exhibited mild anti-invasive and anti-migratory effects upon this cell line. Apparently, HIF-1alpha played a predominant role in retarding pancreatic cancer cell invasion and migration.
MA is a triterpene naturally occurring in many plant foods. Based on its natural property, this agent seems safe for human. However, further in vivo study is necessary to verify its efficiency and safety. In addition, it should be pointed out that MA is a lipophilic compound, and the supplement of this agent at high doses may cause hepatic burden, and interfere its application. Thus, using this agent for anti-cancer therapy should consider hepatic condition.
In summary, maslinic acid treatments at 4-16 M caused apoptosis and declined invasion ad migration in esophagus and stomach cancer cells. This triterpene increased DNA fragmentation, and caspase-3 and caspase-8 activities, as well as lowered ROS, VEGF and TGF-beta1 production in these cancer cell lines. This agent also effectively suppressed the expression of cleaved caspase-3, HIF-1alpha, MMP-2 and MMP-9. These findings indicated that maslinic acid was a potent agent against esophagus and stomach cancers.
Conflict of interest statement None 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
REFERENCES
Allouche, Y., Beltrán, G., Gaforio, J.J., Uceda, M., & Mesa, M.D. (2010). Antioxidant and antiatherogenic activities of pentacyclic triterpenic diols and acids. Food and Chemical Toxicology, 48, 2885-2890.
Augoff, K., Grabowski, K., Rabczynski, J., Kolondra, A., Tabola, R., & Sikorski, A.F. (2009). Expression of decorin in esophageal cancer in relation to the expression of three isoforms of transforming growth factor-beta ( TGF -beta1, -beta2, and -beta3) and matrix metalloproteinase-2 activity. Cancer Investigation, 27, 443-452.
Cui, T., Li, J.Z., Kayahara, H., Ma, L., Wu, L.X., & Nakamura, K. (2006). Quantification of the polyphenols and triterpene acids in Chinese hawthorn fruit by high-performance liquid chromatography. Journal of Agricultural and Food Chemistry, 54, 4574-4581.
Frejlich, E., Rudno-Rudzińska, J., Janiszewski, K., Salomon, L., Kotulski, K., Pelzer, O., Grzebieniak, Z., Tarnawa, R., & Kielan, W. (2013). Caspases and their role in gastric cancer . Advances in Clinical and Experimental Medicine, 22, 593-602.
Fuyuhiro, Y., Yashiro, M., Noda, S., Kashiwagi, S., Matsuoka, J., Doi, Y., Kato, Y., Hasegawa, T., Sawada, T., & Hirakawa, K. (2011). Upregulation of cancer -associated myofibroblasts by TGF -β from scirrhous gastric carcinoma cells. British Journal of Cancer, 105, 996-1001.
Griffiths, E.A., Pritchard, S.A., Valentine, H.R., Whitchelo, N., Bishop, P.W., Ebert, M.P., Price, P.M., Welch, I.M., & West, C.M. (2007). Hypoxia -inducible factor-1alpha expression in the gastric carcinogenesis sequence and its prognostic role in gastric and gastro-oesophageal adenocarcinomas. British Journal of Cancer, 96, 95-103.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
Groblewska, M., Siewko, M., Mroczko, B., & Szmitkowski, M. (2012). The role of matrix metalloproteinases (MMPs) and their inhibitors (TIMPs) in the development of esophageal cancer . Folia Histochemica Et Cytobiologica, 50, 12-9.
Hamano, Y., Zeisberg, M., Sugimoto, H., Lively, J.C., Maeshima, Y., Yang, C., Hynes, R.O., Werb, Z., Sudhakar, A., & Kalluri, R. (2003). Physiological levels of tumstatin, a fragment of collagen IV alpha3 chain, are generated by MMP-9 proteolysis and suppress angiogenesis via alphaV beta3 integrin. Cancer Cell, 3, 589-601.
Hong, J., Resnick, M., Behar, J., Wands, J., Delellis, R.A., & Cao, W. (2011). Role of RAC1 in regulation of NOX5-S function in Barrett's esophageal adenocarcinoma cells. American Journal of Physiology. Cell Physiology,301, C413–C420.
Hu, M., Xu, L., Yin, L., Qi, Y., Li, H., Xu, Y., Han, X., Peng, J., & Wan, X. (2013).
Cytotoxicity of dioscin in human gastric carcinoma cells through death receptor and mitochondrial pathways. Journal of Applied Toxicology, 33, 712-722.
Juan, M.E., Planas, J.M., Ruiz-Gutierrez, V., Daniel, H., & Wenzel, U. (2008).
Antiproliferative and apoptosis-inducing effects of maslinic and oleanolic acids, two pentacyclic triterpenes from olives, on HT-29 colon cancer cells. British Journal of Nutrition, 100, 36-43.
Khamly, K., Jefford, M., Michael, M., & Zalcberg, J. (2006). Recent developments in the systemic therapy of advanced gastroesophageal malignancies. Expert Opinion on Investigational Drugs. 15, 131-153.
Kajdaniuk, D., Marek, B., Foltyn, W., & Kos-Kudła, B. (2011). Vascular endothelial growth factor ( VEGF ) - part 2: in endocrinology and oncology. Endokrynologia Polska, 62, 456-464.
Li, L., Yue, G.G., Lau, C.B., Sun, H., Fung, K.P., Leung, P.C., Han, Q., & Leung, P.S. (2012). Eriocalyxin B induces apoptosis and cell cycle arrest in pancreatic
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
adenocarcinoma cells through caspase- and p53-dependent pathways. Toxicology and Applied Pharmacology, 262, 80-90.
Lin, C.C., Huang, C.Y., Mong, M.C., Chan, C.Y., & Yin, M.C. (2011). Antiangiogenic potential of three triterpenic acids in human liver cancer cells. Journal of Agricultural and Food Chemistry, 59, 755-762.
Manikandan, P., Murugan, R.S., Priyadarsini, R.V., Vinothini, G., & Nagini, S. (2010).
Eugenol induces apoptosis and inhibits invasion and angiogenesis in a rat model of gastric carcinogenesis induced by MNNG. Life Science, 86, 936-941.
Milano, F., Mari, L., van de Luijtgaarden, W., Parikh, K., Calpe, S., & Krishnadath, K.K. (2013). Nano-curcumin inhibits proliferation of esophageal adenocarcinoma cells and enhances the T cell mediated immune response. Frontiers in Oncology, 3, 137. Noma, K., Smalley, K.S., Lioni, M., Naomoto, Y., Tanaka, N., El-Deiry, W., King, A.J.,
Nakagawa, H., & Herlyn, M. (2008). The essential role of fibroblasts in esophageal squamous cell carcinoma-induced angiogenesis. Gastroenterology, 134, 1981-1993. Park, J.H., Kim, T.Y., Jong, H.S., Kim, T.Y., Chun, Y.S., Park, J.W., Lee, C.T., Jung,
H.C., Kim, N.K., & Bang, Y.J. (2003). Gastric epithelial reactive oxygen species prevent normoxic degradation of hypoxia-inducible factor-1alpha in gastric cancer cells. Clinical Cancer Research, 9, 433-440.
Prasad, R., Vaid, M., & Katiyar, S.K. (2012). Grape proanthocyanidin inhibit pancreatic cancer cell growth in vitro and in vivo through induction of apoptosis and by targeting the PI3K/Akt pathway. PLoS One, 7, e43064.
Pugh, C.W., & Ratcliffe, P.J. (2003). Regulation of angiogenesis by hypoxia: role of the HIF system. Nature Medicine, 9, 677-684.
Reyes, F.J., Centelles, J.J., Lupiáñez, J.A., & Cascante, M. (2006). (2Alpha,3beta)- 2,3- dihydroxyolean-12-en-28-oic acid, a new natural triterpene from Olea europea,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
induces caspase dependent apoptosis selectively in colon adenocarcinoma cells. FEBS Letter, 580, 6302-6310.
Shimojo, Y., Akimoto, M., Hisanaga, T., Tanaka, T., Tajima, Y., Honma, Y., & Takenaga, K. (2013). Attenuation of reactive oxygen species by antioxidants suppresses hypoxia-induced epithelial-mesenchymal transition and metastasis of pancreatic cancer cells. Clinical & Experimental Metastasis, 30, 143-154.
Song, I.S., Wang, A.G., Yoon, S.Y., Kim, J.M., Kim, J.H., Lee, D.S., & Kim, N.S. (2009).
Regulation of glucose metabolism-related genes and VEGF by 1alpha and HIF-1beta, but not HIF-2alpha, in gastric cancer . Experimental and Molecular Medicine, 41, 51-58.
Xue, G., Zou, X., Zhou, J.Y., Sun, W., Wu, J., Xu, J.L., & Wang, R.P. (2013). Raddeanin A induces human gastric cancer cells apoptosis and inhibits their invasion in vitro.
Biochemical and Biophysical Research Communication, 439, 196-202.
Yin, M.C., Lin, M.C., Mong, M.C., & Lin, C.Y. (2012). Bioavailability, distribution, and antioxidative effects of selected triterpenes in mice. Journal of Agricultural and Food Chemistry, 60, 7697-7701.
Zhang, Y.S., Xie, J.Z., Zhong, J.L., Li, Y.Y., Wang, R.Q., Qin, Y.Z., Lou, H.X., Gao, Z.H., & Qu, X.J. (2013). Acetyl-11-keto-β-boswellic acid (AKBA) inhibits human gastric carcinoma growth through modulation of the Wnt/β-catenin signaling pathway. Biochimica et Biophysica Acta, 1830, 3604-3615.
Zheng, H., Takahashi, H., Murai, Y., Cui, Z., Nomoto, K., Niwa, H., Tsuneyama, K., & Takano, Y. (2006). Expressions of MMP-2 , MMP-9 and VEGF are closely linked to growth, invasion, metastasis and angiogenesis of gastric carcinoma. Anticancer Research, 26, 3579-3583. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Zhu, H., Feng, Y., Zhang, J., Zhou, X., Hao, B., Zhang, G., & Shi, R. (2011). Inhibition of hypoxia inducible factor 1 α expression suppresses the progression of esophageal squamous cell carcinoma. Cancer Biology & Therapy, 11, 981-987.
Figure caption
Fig. 1-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon cell viability (% of control), determined by MTT assay, in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8). a-dMeans among bars without a common letter differ, P<0.05.
Fig. 2-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon cleaved caspase-3, Bcl-2 and Bax expression in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8). a-dMeans among bars without a common letter differ, P<0.05.
Fig. 3-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon caspase-3 and caspase 8 activities (% of control) in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8). a-dMeans among bars without a common letter differ, P<0.05.
Fig. 4-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon cell invasion (% of control) and migration (% of control) in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8). a-dMeans among bars without a common letter differ, P<0.05.
Fig. 5-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon HIF-1alpha, MMP-2 and MMP-9 expression in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8). a-dMeans among bars without a common letter differ, P<0.05. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Fig. 1 1 2 3 4 5
Fig. 2 OE33 SGC-7901 Panc-1 Cleaved caspase-3 Bax Bcl-2 GAPDH MA 0 4 8 16 0 4 8 16 0 4 8 16 1 2
Fig. 3 1 2 3 4 5 6
Fig. 4 1
2 3
Fig. 5 OE33 SGC-7901 Panc-1 HIF-1alpha MMP-2 MMP-9 GAPDH MA 0 4 8 16 0 4 8 16 0 4 8 16 1 2 3
Table 1-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon DNA fragmentation, determined as enrichment factor; and ROS level (nmol/mg protein) in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8). DNA fragmentation OE33 SGC-7901 Panc-1 Control 1.00.08a 1.00.1a 1.00.05a MA, 4 1.420.11b 1.370.07b 1.090.06a MA, 8 1.930.20c 1.860.14c 1.450.1b MA, 16 2.460.23d 2.290.19d 1.520.13b ROS OE33 SGC-7901 Panc-1 Control 1.830.16d 2.130.21d 2.290.18c MA, 4 1.390.09c 1.760.15c 1.800.13b MA, 8 0.950.05b 1.110.10b 1.220.11a MA, 16 0.600.04a 0.730.06a 1.070.15a
a-dMeans in a column without a common letter differ, P<0.05. 1 2 3 4 5 6
Table 2-Effect of maslinic acid (MA) at 0 (control), 4, 8 or 16 M upon VEGF level (pg/mg protein) and TGF-beta1 level (ng/mg protein) in human OE33, SGC-7901 and Panc-1 cells. Cells were exposed to MA for 48 hr at 37°C. Data are mean SD (n = 8).
VEGF OE33 SGC-7901 Panc-1 Control 16812d 17015d 15711b MA, 4 1378c 12910c 1489b MA, 8 10110b 954b 1145a MA, 16 747a 686a 1067a TGF-beta1 OE33 SGC-7901 Panc-1 Control 18914d 21116d 20211b MA, 4 1508c 18310c 1947b MA, 8 1064b 1317b 1529a MA, 16 816a 925a 14310a
a-dMeans in a column without a common letter differ, P<0.05. 1
2 3