Effects of culturing conditions on production of D-hydantoinase from recombinant Escherichia coli
9
0
0
全文
(2) Production of D-hydantoinase from recombinant Escherichia coli. A recombinant Escherichia coli strain producing a significant amount of Dht has been constructed [26]. This recombinant E. coli strain harbors the dht gene from Agrobacterium radiobacter under the control of the T5 promoter. In this study, we investigated the optimal culture conditions for Dht production in shaker flasks or a benchtop fermentor under both batch and fed-batch operations. Furthermore, we report a fed-batch procedure that allows high cell density culture and leads to high-level expression of Dht.. Materials and Methods Chemicals 5-(4-Hydroxyphenyl)-hydantoin (HPH) was obtained from Tokyo Chemical Industry (Tokyo, Japan). The Ncarbamoyl-D-p-hydroxyphenylglycine (NC-D-p-HPG) standard was synthesized and kindly provided by Dr. Wu-Yung Yang (Union Chemical Laboratories, Industrial Technology Research Institute, Hsinchu, Taiwan). Yeast extract, tryptone and agar were obtained from Difco Laboratories (Detroit, MI, USA). Hydantoin, Tris, ampicillin (sodium salt), tetracycline (free base), isopropyl β-D-thiogalactoside (IPTG), bromthymol blue (sodium salt; BTB) and Coomassie brilliant blue R-250 were purchased from Sigma Chemical Co. (St Louis, MO, USA). Acrylamide was obtained from Merck KGaA (Darmstadt, Germany). All other chemicals were of reagent grade and were purchased from commercial suppliers.. Bacterial strain A 1.4-kb Kpn 1-Pst 1 fragment containing the dht gene of Agrobacterium radiobacter DUL-DH 101 and the sequence encoding the N-terminal His-tag was cloned into expression vector pQE30 (Qiagen, Valencia, CA, USA) to generate pQE30/dht using standard procedures [27]. NovaBlue E. coli (endA1 hsdR17 (rk12–mk12–) supE44 thi-1 recA1 gyrA96 relA1 lac F’ [proA+B+ lacIq lacZ∆M15::Tn10(Tetr)]), Novagen Inc. (Madison, WI, USA) was transformed with plasmid pQE30/dht using a classical CaCl2 preparation [26,27]. The dht gene transcription is under the control of the phage T5 promoter and lac operator (Qiagen) and can be induced with IPTG. The transformant, referred to as E. coli NovaBlue(pQE30/dht) was used throughout this study. The strain was maintained as 40% (v/v) glycerol stock at –30°C in Luria-Bertani (LB) medium and was very stable throughout the whole experiment. 314. Media LB medium [27] was employed for the propagation of E. coli transformants under the uninduced state. When necessary, ampicillin and tetracycline were added to the medium at concentrations of 100 and 10 µg/mL, respectively. Modified LB (MLB) medium was LB medium supplemented with the following per liter: 10 g glucose, 0.1 g ampicillin and 0.01 g tetracycline. Semi-synthetic production (SSP) medium contained the following per liter: 10 g glucose, 10 g yeast extract, 7 g K2HPO4, 3 g KH2PO4, 0.1 g ampicillin and 0.01 g tetracycline. A synthetic medium modified from that of Horn et al [28] contained the following per liter: 5 g glucose, 0.5 g NaCl, 2 g (NH4)2SO4, 9 g Na2HPO4, 3 g K 2HPO4 and trace elements of 60 mg/L Fe(III) citrate, 3 mg/L H3BO3, 15 mg/L MnCl2· 4H2O, 8.4 mg/L EDTA·2Na, 2 mg/L CuSO 4 ·5H 2 O, 2.5 mg/L Na 2 MoO 4 ·2H 2 O, 2.5 mg/L CoCl 2 ·6H 2O, 8 mg/L ZnCl2. Sterilization was performed at 121°C for 20 min. Glucose was autoclaved separately from the rest of the medium and added into the medium prior to inoculation. Ampicillin and tetracycline were sterilized by filtration.. Culture conditions For cell activation, 0.2 mL frozen stock culture was added to 25 mL of MLB medium in a 50 mL Falcon tube (Falcon Plastics, Oxnard, CA, USA) and the tube shaken at 37°C and 125 rpm for 12 h. The inoculum culture was then prepared by adding 0.1% (v/v) of this reactivated culture to 50 mL MLB medium in a 250 mL Erlenmeyer flask, followed by culturing under the same conditions for 12 h. For shaker-flask culture, 2 mL of inoculum culture was added to 100 mL production medium in a 500-mL Hinton flask. The cultures were incubated at 37°C in a rotary shaker set at 125 rpm and 7 cm stroke (Model 707R, Hotech Instruments Corp., Taipei, Taiwan). After 6 h cultivation, 0.02 mM IPTG was added and the flasks were moved to another shaker at 27°C and 140 rpm (Model 706R, Hotech Instruments Corp., stroke 3.85 cm) for enzyme induction. For large-scale experiments, batch and fed-batch fermentations were performed in a classical 5 L jar fermentor (Model LS-205, Hotech Instruments Corp.) with a digital measurement and control unit from Suntex Instruments Co., Ltd. (Taipei, Taiwan). Antifoam 289 (Sigma A-5551) was automatically added as necessary to control foaming in the fermentor. Fifty mL of inoculum culture was added to 2 L of initial fermentation medium and the cultivation was carried.
(3) Huang et al. out under the conditions used for the shaker-flask culture. The aeration was kept constant at 2.0 L air/min, and agitation at 1000 rpm. The pH-stat method was employed for the control of substrate feeding. Medium pH was adjusted to 6.9-7.1 during the cultivation processes by the addition of either 9% (v/v) ammonia water or solutions of 600 g/L glucose and 300 g/L yeast extract with a peristaltic pump. The pH of the culture increased as glucose was depleted. When the pH exceeded the set high limit of 7.1, an equal volume of glucose solution and yeast extract solution was automatically added so that the glucose concentration in the culture broth was increased while the pH was decreased. In the fed-batch culture, the concentration of glucose in the medium was maintained below 3.3 g/L. When the pH was decreased below the lower limit of 6.9, ammonia water was automatically added to recover pH as well as supply a nitrogen source. No nutrient was fed when the pH was between 6.9 and 7.1. Antibiotics were not used in the fermentor study. The dissolved oxygen level was determined with a steamsterilizable polarographic electrode (InPro 6000 series O2 sensors; Mettler-Toledo GmbH, Switzerland), and the pH of culture broth was measured with a steamsterilizable pH probe (InPro 3030/325 combination pH electrode; Mettler-Toledo GmbH, Switzerland). The time points of feeding and harvesting are reported in the Results section. Expression of the dht gene was induced by adding IPTG to a final concentration of 0.02 mM or as indicated. After induction with IPTG, cells were cultured for an additional 30 or more hours and harvested by centrifugation. Throughout the cultivation, pH, temperature, and dissolved oxygen were continuously monitored and recorded. Samples were automatically taken at various intervals for analysis.. Preparation of crude extracts by ultrasonication The cells were obtained by centrifugation (2000 × g, 15 min), washed twice with 0.05 M Tris-HCl buffer (pH 8.0), then resuspended in the same buffer. Aliquots of 2 mL cell suspension corresponding to 30 absorbance [optical density (OD600)] units were then disrupted with a cell sonicator (Model XL2020, 419A microtip probe; Misonix Inc., NY, USA) for 2 sec every 3 sec for a total of 165 sec in an ice bath. After centrifugation at 12,000 × g for 5 min at 4°C, the supernatants (soluble protein fraction) were used as the crude enzyme preparations. The pellets (inclusion body fraction) were washed twice with buffer and. resuspended in an equal volume of the buffer. The supernatants and pellets were analyzed by sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) [29]. The enzyme with a 6-histidine-residue tag at the aminoterminal end was further purified from the supernatants on Ni-affinity columns as described by Hsu et al [30].. Assay of D-hydantoinase activity Fifty µL of diluted supernatant or partially purified enzyme solution was incubated with 1% HPH (molecular weight [MW] 192.17) in 0.05 M Tris-HCl buffer (pH 8.0) in a total volume of 1 mL. The reaction was allowed to proceed at 37°C for 10 min in a reciprocating shaker water bath at 100 rpm, 3.85 cm stroke, and was terminated and deproteinized by the addition of 200 µL of 12% trichloroacetic acid (TCA). After removing the precipitate by centrifugation (8500 × g, 5 min), the amount of NC-D-p-HPG (MW 210.16) in the supernatant was quantified. One unit of Dht activity was defined as the amount of enzyme required to produce 1 µmole of NC-D-p-HPG per min under the assay conditions. A blank with TCAdenaturated enzyme was assayed for all samples. Dht activity in colonies was detected on an LBHBTB agar plate, a blue plate containing LB supplemented with 10.0 g/L hydantoin, 0.1 g/L BTB, 1.0 mL 1 M MnCl 2·4H 2O, 0.25 mL 20 mM IPTG, 1.0 mL 1% tetracycline and 20.0 g/L Bacto-agar. Colonies that express Dht convert hydantoin to hydantoic acid, thus lowering the pH, and a yellowish zone was observed in the plate around the Dht-positive colonies against a blue background after 36 h incubation at 32°C.. Analytical methods Dry cell weight (DCW) was measured after the cells were washed twice with distilled water and dried to a constant weight at 105°C. Cell density was monitored by measuring the optical density of the culture broth at 600 nm (OD 600) using a spectrophotometer. The culture broth was generally diluted with distilled water to yield an OD600 value of 0.2-0.5. Biomass was calculated from the OD600 and converted to DCW per liter according to predetermined calibration data. One OD unit corresponds to 0.33 ± 0.05 gDCW/L. The titers of total viable and ampicillin-resistant cells were determined by plating serial 10-fold diluted samples on LB agar plates in the presence or absence of 100 µg/mL ampicillin in triplicate. The number of colony-forming units was determined after incubating the plates at 37°C for 18 h. Glucose concentrations were measured 315.
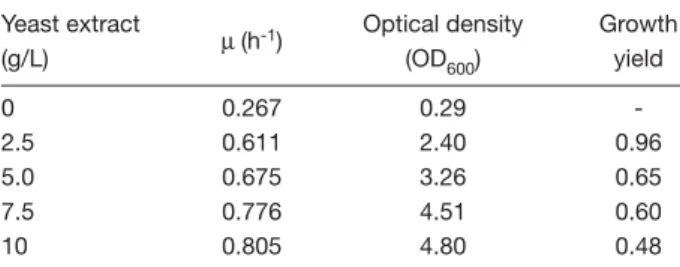
(4) Production of D-hydantoinase from recombinant Escherichia coli. colorimetrically using dinitrosalicylic acid reagent [31]. Protein concentration was measured by the method of Bradford [32], using bovine serum albumin as a standard. SDS-PAGE was performed according to the method of Laemmli [29], using 12.5% polyacrylamide gels. Proteins were visualized by staining with Coomassie brilliant blue R-250 (Sigma) and quantified by densitometric analysis of stained bands on the gels with a Kodak Digital Science Electrophoresis Documentation and Analysis System. NC-D-p-HPG was quantitated by high-performance liquid chromatography (HPLC) or estimated spectrophotometrically at 450 nm using Ehrlich’s reagent (10% p-dimethylaminobenzaldehyde in 6.0 N HCl) as described by Grifantini et al [13]. HPLC analysis was performed on a reverse phase C18 column (4.6 × 250 mm; Merck, USA) [33]. Samples were eluted isocratically with acetonitrile:water (95:5 v/v) at a flow rate of 0.8 mL/min and detected at 274 nm using an ABI 783A UV detector (Applied Biosystems, Inc., Foster City, CA, USA). The detector was coupled to a chromatocorder for quantification. The retention times for NC-D-p-HPG and HPH were 6.5 and 9.8 min, respectively. The correlation coefficient of concentration and peak area for NC-D-p-HPG was 0.9997. The results obtained by colorimetric method and HPLC were in good agreement, with the variation around 3%. Stability of the recombinant plasmid was determined by culturing plasmid-bearing E. coli in a 500 mL Hinton flask containing 100 mL SSP in the absence of ampicillin. Cells (0.1 mL) were inoculated into 100 mL of the next sterile medium. The proportion of Dht-producing cells in the population was determined by plating the cells on LBHBTB agar plates and incubating at 32°C for 36 h.. Results Plasmid stability The plasmid stability of E. coli NovaBlue(pQE30/dht) was investigated in SSP medium in the absence of ampicillin. At various intervals after inoculation, the proportions of Dht-producing cells in serially diluted samples were determined by plating the cells on LBHBTB agar plates. The development of yellowish color around each colony indicated that there was no loss in its capability to produce Dht. Plate assay indicated that the recombinant plasmid was stable over 200 generations in the ampicillin-free SSP medium. All colonies maintained the Dht plasmid after 200 316. generations. In addition, Dht levels were comparable among the different subcultures.. Effects of medium composition on cell growth LB medium, a synthetic medium and SSP medium were used to evaluate the influence of medium composition on cell growth in 500 mL Hinton flasks. Although there was little or no growth of E. coli NovaBlue(pQE30/dht) in the synthetic medium because the strain used requires thiamine, good growth rates and high cell density were achieved in both LB and SSP media. The SSP medium was selected because of its lower cost and simple composition. The presence of yeast extract significantly increased the growth rate of this culture. With increasing amounts of yeast extract, the cell concentration and the specific growth rate increased, but the growth yields decreased in terms of yeast extract concentration (Table 1). The cell mass increased with increasing concentrations of yeast extract up to 7.5 g/L. Other components were tested but no significant effects were observed.. IPTG induction Shake flask experiments were conducted to evaluate the influence of IPTG on the induction of Dht production in E. coli NovaBlue(pQE30/dht). The optimal temperature and IPTG concentration for Dht expression were found to be 27°C and 0.02 mM, respectively (Fig. 1). Low-level induction by IPTG allowed the growth of the recombinant strain to continue, thus prolonging the production phase of the induced cells. Lower temperature reduced the growth rate, therefore minimizing the formation of inhibitory byproducts such as acetate, and reducing the demand for oxygen. Even in the absence of IPTG, some amount of Dht was expressed at 27°C or 37°C. In addition, the formation of inclusion bodies was also significantly reduced (Fig. 2). Fig. 3 further shows that the dht gene could be efficiently induced with 0.02 mM IPTG in the mid-exponential phase of cell growth (at around Table 1. Effects of yeast extract concentration on the growth rate of recombinant Escherichia coli NovaBlue(pQE30/dht) Yeast extract (g/L). µ (h-1). Optical density (OD600). Growth yield. 0 2.5 5.0 7.5 10. 0.267 0.611 0.675 0.776 0.805. 0.29 2.40 3.26 4.51 4.80. 0.96 0.65 0.60 0.48.
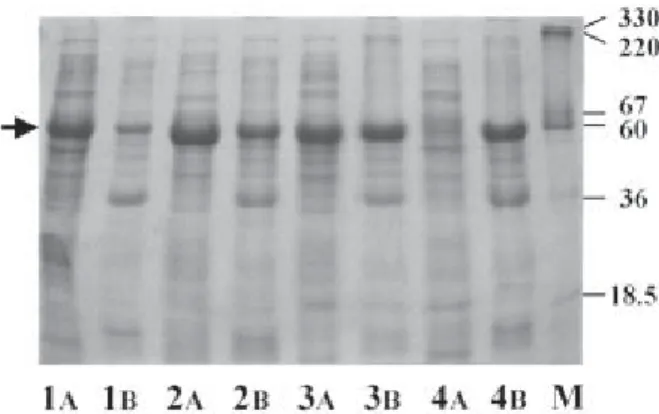
(5) Huang et al. Relative activity of Dht (%). 120 27°C 37°C. 100 80 60 40 20 0 0. 0.01 0.02 0.04 0.06 0.10 IPTG concentration (mM). 1.00. Fig. 1. Effects of isopropyl β-D-thiogalactoside (IPTG) concentration and induction temperature on D-hydantoinase (Dht) expression. E. coli NovaBlue(pQE30/dht) was grown in Luria-Bertani medium in shaker flasks at 37°C under 125 rpm agitation. Following 6-h cultivation, different amounts of IPTG were added. The flask, containing two-thirds of the original culture, was transferred to another shaker set at 27°C and 140 rpm and cultured for an additional 6 h. The Dht activity of the culture broth was determined as described in Materials and Methods. The highest activity was taken as 100%. The percent activities under various IPTG concentrations and temperature conditions are shown. The mean value of at least 2 experiments is indicated by column height. The variation was less than 10%.. 6 h). The maximal productivity, obtained when induced at 6 h following inoculation, was about 2.6-fold that obtained when IPTG induction was carried out 3 h earlier in the log phase of growth. No Dht activity was detected when cells were induced at the late log phase of growth (at 9 h).. D-Hydantoinase production Based on the results described above, optimal conditions for Dht production of E. coli NovaBlue(pQE30/dht) in a 5 L jar fermentor were as follows. The optimal composition of batch medium was glucose, 0.25%; yeast extract, 0.75%; (NH4)2SO4, 0.25%; KH2PO4, 0.2%, while that for the feeding solution was glucose, 60%; yeast extract, 30%; ammonia water, 9%. During cultivation, the agitation and aeration rates were kept at 1000 rpm and 1 vvm, respectively and the pH was controlled automatically at 7.0 ± 0.1, as described in Materials and Methods. Following 25-h cultivation, which was about the mid-log growth phase, 0.02 mM IPTG was added the temperature was shifted from 32 to 27°C. Under these conditions, a typical time course of the cultivation of E. coli NovaBlue(pQE30/dht) is shown. Fig. 2. Sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) analysis of the soluble (A) and insoluble (B) crude extracts of E. coli NovaBlue(pQE30/dht) induced at different temperatures. The positions of molecular size markers in kDa are indicated on the right. Cells were grown in semi-synthetic production medium in shake flasks at 37°C under 125 rpm agitation. Following 6-h cultivation, 0.02 mM isopropyl β-Dthiogalactoside was added, and the flask was transferred immediately to another shaker, set at the different temperatures shown below, for enzyme induction. The cells were then harvested and lysed as described in Materials and Methods. Ten µg of protein was loaded in each lane on a 12.5% SDSPAGE gel as described. Lane 1A and 1B, 27°C; lane 2A and 2B, 30°C; lane 3A and 3B, 32°C; lane 4A and 4B, 37°C; lane M, molecular mass markers in kDa. Bands corresponding to the 60 kDa protein are indicated by the arrow.. in Fig. 4. The specific growth rate was determined from the slope of the semi-logarithmic plot of cell density (OD600) versus cultivation time. During the first 10 h of culture, a high specific growth rate of 0.423/h was obtained. However, the specific growth rate decreased to 0.026/h due to a combination of factors including IPTG induction, down-shift of temperature, and presumably carbon, oxygen, and growth factor depletion. The beginning of this second phase coincided with the onset of initial glucose depletion. Glucose was fed at this stage to maintain the maximum cell concentration for intracellular protein synthesis. To avoid excessive feeding of glucose, which is known to cause acetate accumulation [34], the nutrient feeding solution was added only when the glucose concentration declined to a certain level as indicated by an increase in pH. The highest Dht activity of about 400,000 U/L or 5 g/L of Dht was attained within 48 h and the activity remained relatively high even at 54 h. The maximum cell concentration reached around OD600 75 (39 h) or 25 gDCW/L. A summary of Dht production under various culture conditions in E. coli NovaBlue(pQE30/ dht) is shown in Table 2. 317.
(6) Production of D-hydantoinase from recombinant Escherichia coli. Relative activity of D-hydantoinase (%). 120 100 80 60 40 20 0. 0. 10. 15. 20 25 Culture time (h). 30. 35. 40. Fig. 3. Effect of induction time on D-hydantoinase (Dht) expression. Cells were grown in semi-synthetic production medium at 37°C at an agitation rate of 125 rpm. After 3- ( ), 6- ( ), or 9-h ( ) cultivation, 0.02 mM isopropyl β-D-thiogalactoside was added, and the flask was transferred immediately to another shaker at conditions of 27°C and 140 rpm, for enzyme induction. At different time intervals, the Dht activity of the culture broth was determined as described in Materials and Methods. The highest activity was taken as 100%.. SDS-PAGE analysis of Dht In order to determine if all Dht was appropriately processed to an active state, aliquots of cultures under different induction temperatures were analyzed by SDSPAGE (Fig. 2). A prominent protein band with an apparent molecular mass of 60 kDa, corresponding to 2.5. the subunit MW of Dht was observed [35]. The 6-Histagged-Dht was purified to homogeneity by His-Bind resin affinity chromatography. Purified Dht had Dhydantoinase activity also showed a single band with an apparent molecular mass of approximately 60 kDa. After 6 h induction at 27°C, about 80% of the enzyme. 100 400 µ2 = 0.026 h-1. 2.0. 0.5. 60 OD600. log OD600. 1.0. µ1 = 0.423 h-1 200. 40. 0.0 0.5. 100. 20. 12 10 8 6 4. -1.0. 2 0. 0. -1.5 0. 10. 20 30 Culture time (h). 40. Glucose (g/L). 300. Dht activity (103 U/L). 80 1.5. 0. 50. Fig. 4. Profile of D-hydantoinase (Dht) production by a fed-batch high cell density culture of Escherichia coli NovaBlue(pQE30/ dht). The composition (per L) of the initial fermentation medium was: 2.5 g glucose, 7.5 g yeast extract, 2.5 g (NH4)2SO4, 2.0 g KH2PO4; and that of the feeding solution was: 600 g glucose, 300 g yeast extract, 333 mL of 28% ammonia water. The time courses of biomass [optical density (OD600, )], glucose (––), Dht activity ( ) as well as specific growth rates are shown. The closed box symbol ( ) represents log OD600, which was used to determine the specific growth rate. Agitation speed and aeration rate were kept constant at 1000 rpm and 1 vvm, respectively, throughout the cultivation. Glucose concentration and pH were controlled at below 3.3 g/L and 6.9-7.1, respectively. The time of isopropyl β-D-thiogalactoside addition, followed immediately by temperature shift from 32 to 27°C, is indicated by the arrow.. 318.
(7) Huang et al. Table 2. Summary of D-hydantoinase production under different culture conditions in Escherichia coli NovaBlue(pQE30/dht) Hinton flask. Isopropyl β-D-thiogalactoside (mM) Cultivation time (h) Final optical density (OD600) Yield (U/g glucose) Specific activity (U/g dry cell weight) Productivity (U/L/h) D-hydantoinase activity (U/L) Increase of Dht activity Yield of purified enzyme (g/L). 5 L fermentor. Luria-Bertani medium. Semi-synthetic production medium. High cell density culture. 0.02 32 7.3 6100 460 14,700 ND. 0.02 32 12 6500 16,400 2000 65,000 4.42-fold ND. 0.02 48 75 4950 16,200 8300 400,000 27.2-fold 5. Abbreviation: ND = not determined. was present as active and soluble protein (Fig. 2, lane 1A and 1B). The ratios of soluble protein to total Dht were 63% (Fig. 2, lane 2A and 2B) and 51% (Fig. 2, lane 3A and 3B) when induction temperatures were 30°C and 32°C, respectively. No soluble Dht protein was detectable when the induction temperature was 37°C (Fig. 2, lane 4A and 4B).. Discussion Hydantoinases belong to the family of cyclic amidases. They are closely related to dihydropyrimidinases and most likely take part in the catabolism of pyrimidines and thus might be harmful to cells. In this study, the optimal culture conditions to allow the production of a significant amount of the active and soluble form of Dht were investigated. The application of these modes of fed-batch cultivation resulted in a 6 to 7-fold increase in Dht activity compared to shake flask cultivation (Table 2). The enzyme production process can be divided into 2 phases (Fig. 4). A rapid enzyme production phase positively correlated to the late exponential phase of growth, which was followed by a slower phase of production resulting from a slower growth rate. No significant toxicity of the over-expressed foreign protein to the host was observed. Dht activity continued to increase even after the stationary phase was reached. After the maximal cell concentration was reached, the culture was maintained for an additional 20-25 h. During this period, no loss in enzyme activity or significant lysis was observed, as the cell concentration remained relatively constant at 25.0 ± 0.2 g/L and the activity of Dht in the culture broth was about 0.2% of the total. Under simple culture conditions, at a constant feed rate without oxygen enrichment, a satisfactory quantity of. total Dht (5 g/L) could be obtained. Dht activity as high as 1,900,000 U/L has been reported from a recombinant E. coli [25]. However, the enzymatic activity in that report was defined as the hydrolysis of hydantoin, instead of that of D,L-p-hydroxyphenylhydantoin which is utilized in industry and in our study. The advantageous features of E. coli NovaBlue(pQE30/dht) for industrial scale enzyme production include its ability to stably retain plasmid and to produce significant amounts of Dht from an antibiotic-free medium. Our results also showed that not only was the yield of the recombinant product high but also the fraction of insoluble, functionally inactive, protein was generally quite low (Fig. 2). The production of industrial enzymes as an insoluble form is obviously undesirable, as additional (and generally inefficient) measures are required to refold the protein. Furthermore, the enzyme could be easily purified by a 1-step affinity chromatography. The crude or purified overproduced Dht can be further utilized, either in solution or in immobilized form, for the production of D-amino acids.. Acknowledgment This study was supported by a grant NSC 87-2316B002-006-BI from the National Science Council, Taiwan, ROC.. References 1. Syldatk C, May O, Altenbuchner J, Mattes R, Siemann M. Microbial hydantoinases – industrial enzymes from the origin of life? Appl Microbiol Biotechnol 1999;51:293-309. 2. Takahashi S, Ohashi T, Kii Y, Kumagai H, Yamada H. Microbial transformation of hydantoins to N-carbamyl-D-amino acids. J Ferment Technol 1979;57:328-32. 319.
(8) Production of D-hydantoinase from recombinant Escherichia coli. 3. Olivieri R, Fascetti E, Angelini L, Degen L. Microbial transformation of racemic hydantoins to D-amino acids. Biotechnol Bioeng 1981;23:2173-83. 4. Ogawa J, Shimizu S. Stereoselective synthesis using hydantoinases and carbamoylases. In: Patel RN, ed. Stereoselective biocatalysis. New York: Marcel Dekker; 2000:1-21. 5. Soong CL, Ogawa J, Honda M, Shimizu S. Cyclic-imidehydrolyzing activity of D-hydantoinase from Blastobacter sp. Strain A17p-4. Appl Environ Microbiol 1999;65:1459-62. 6. Syldatk C, Laufer A, Muller R, Hoke H. Production of optically pure D- and L-α-amino acids by bioconversion of D,L-5monosubstituted hydantoin dervatives. Adv Biochem Eng Biotechnol 1990;41:29-75. 7. Yagasaki M, Ozaki A. Industrial biotransformations for the production of D-amino acids. J Mol Catal, B Enzym 1998; 4:1-11. 8. Sudge SS, Bastawde KB, Gokhale DV, Kalkote UR, Ravindranathan T. Production of D-hydantoinase by halophilic Pseudomonas sp. NCIM 5109. Appl Microbiol Biotechnol 1998;49:594-9. 9. Demain AL, Elander RP. The β-lactam antibiotics: past, present, and future. Antonie van Leeuwenhoek 1999;75:5-19. 10. Bradford PA. Extended-spectrum β-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat. Clin Microbiol Rev 2001;14: 933-51. 11. Cohen ML. Epidemiology of drug resistance: Implications for a post-antimicrobial era. Science 1992;257:1050-5. 12. Gold HS, Moellering RC Jr. Antimicrobial-drug resistance. N Engl J Med 1996;335:1445-53. 13. Grifantini R, Galli G, Carpani G, Pratesi C, Frascotti G, Grandi G. Efficient conversion of 5-substituted hydantoins to D-αamino acids using recombinant Escherichia coli strains. Microbiology 1998;144:947-54. 14. Ogawa J, Shimizu S. Diversity and versatility of microbial hydantoin-transforming enzymes. J Mol Catal, B Enzym 1997; 2:163-76. 15. Morin A, Hummel W, Kula MR. Production of hydantoinase from Pseudomonas fluorescens strain DSM 84. Appl Microbiol Biotechnol 1986;25:91-6. 16. Yamada H, Takahashi S, Kii Y, Kumagai H. Distribution of hydantoin hydrolyzing activity in microorganisms. J Ferment Technol 1978;56:484-91. 17. LaPointe G, Viau S, Leblanc D, Robert N, Morin A. Cloning, sequencing, and expression in Escherichia coli of the Dhydantoinase gene from Pseudomonas putida and distribution of homologous genes in other microorganisms. Appl Environ Microbiol 1994;60:888-95. 18. Morin A, Tran Trung NH, LaPointe G, Dubeau H. Conditions used with a continuous cultivation system to screen for D320. hydantoinase-producing microorganisms. Appl Microbiol Biotechnol 1995;43:259-66. 19. Matsuda K, Sakata S, Kaneko M, Hamajima N, Nonaka M, Sasaki M, et al. Molecular cloning and sequencing of a cDNA encoding dihydropyrimidinase from the rat liver. Biochim Biophys Acta 1996;1307:140-4. 20. May O, Nguyen PT, Arnold FH. Inverting enantioselectivity by directed evolution of hydantoinase for improved production of L-methionine. Nature Biotechnol 2000;18:317-20. 21. Morin A, Leblanc D, Paleczek A, Hummel W, Kula MR. Comparison of seven microbial D-hydantoinases. J Biotechnol 1990;16:37-48. 22. Takahashi S, Kii Y, Kumagai H, Yamada H. Purification, crystallization and properties of hydantoinase from Pseudomonas striata. J Ferment Technol 1978;56:492-8. 23. Hartley CJ, Manford F, Burton SG, Dorrington RA. Overproduction of hydantoinase and N-carbamoylamino acid amidohydrolase enzymes by regulatory mutants of Agrobacterium tumefaciens. Appl Microbiol Biotechnol 2001; 57:43-9. 24. Achary A, Hariharan KA, Bandhyopadhyaya S, Ramachandran R, Jayaraman K. Application of numerical modeling for the development of optimized complex medium for Dhydantoinase production from Agrobacterium radiobacter NRRL B 11291. Biotechnol Bioeng 1997;55:148-54. 25. Lee DC, Kim GJ, Cha YK, Lee CY, Kim HS. Mass production of thermostable D-hydantoinase by batch culture of recombinant Escherichia coli with a constitutive expression system. Biotechnol Bioeng 1997;56:449-55. 26. Perng RY. Characterization and protein engineering of Dhydantoinase from Agrobacterium radiobacter and mutational analysis of the catalytic sites of D-hydantoinase from Bacillus caldolyticus. M.S. thesis, National Chung-Hsing University, Taichung, Taiwan, ROC; 1998. 27. Sambrook J, Fritsch EF, Maniatis T, eds. Molecular cloning: A laboratory manual. 2nd ed. New York: Cold Spring Harbor Laboratory Press; 1989. 28. Horn U, Strittmatter W, Krebber A, Knupfer U, Kujau M, Wenderoth R, et al. High volumetric yields of functional dimeric miniantibodies in Escherichia coli, using an optimized expression vector and high-cell-density fermentation under nonlimited growth conditions. Appl Microbiol Biotechnol 1996; 46:524-32. 29. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227: 680-5. 30. Hsu WH, Chien FT, Hsu CL, Wang TC, Yuan HS, Wang WC. Expression, crystallization and preliminary X-ray diffraction studies of N-carbamyl-D-amino-acid amidohydrolase from Agrobacterium radiobacter. Acta Cryst D 1999;55:694-5..
(9) Huang et al. 31. Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 1959;31:426-8. 32. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248-54. 33. Runser S, Ohleyer E. Properties of the hydantoinase from Agrobacterium sp. IP I-671. Biotechnol Lett 1990;12:259-64. 34. Akesson M, Hagander P, Axelsson JP. Avoiding acetate. accumulation in Escherichia coli cultures using feedback control of glucose feeding. Biotechnol Bioeng 2001;73: 223-30. 35. Runser SM, Meyer PC. Purification and biochemical characterization of the hydantoin hydrolyzing enzyme from Agrobacterium species: A hydantoinase with no 5,6dihydropyrimidine amidohydrolase activity. Eur J Biochem 1993;213:1315-24.. 321.
(10)
數據


相關文件
You are given the wavelength and total energy of a light pulse and asked to find the number of photons it
好了既然 Z[x] 中的 ideal 不一定是 principle ideal 那麼我們就不能學 Proposition 7.2.11 的方法得到 Z[x] 中的 irreducible element 就是 prime element 了..
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
volume suppressed mass: (TeV) 2 /M P ∼ 10 −4 eV → mm range can be experimentally tested for any number of extra dimensions - Light U(1) gauge bosons: no derivative couplings. =>
For pedagogical purposes, let us start consideration from a simple one-dimensional (1D) system, where electrons are confined to a chain parallel to the x axis. As it is well known
The observed small neutrino masses strongly suggest the presence of super heavy Majorana neutrinos N. Out-of-thermal equilibrium processes may be easily realized around the
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
incapable to extract any quantities from QCD, nor to tackle the most interesting physics, namely, the spontaneously chiral symmetry breaking and the color confinement..
