ELSEVIER SCIENTIFIC
PUBLISHERS IRELAND Plant Science 96 (1994) 41-44
p an cience
Lipid peroxidation and antioxidative enzymes in senescing
tobacco leaves during post-flooding
W e e i P i r n g H u r n g , C h i n g H u e i K a o *Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (Received 4 August 1993; revision received 15 October 1993; accepted 18 October 1993)
Abstract
Role of lipid peroxidation and antioxidative enzymes (superoxide dismutase and catalase) in leaf senescence of tobacco plants during the recovery phase after 2 days of flooding was investigated. The senescence of leaves was follow- ed by measuring the decrease in level of chlorophyll. An enhancement in leaf senescence was observed in flooded plants during the recovery phase. Increase in lipid peroxidation is closely correlated with senescence in leaves of flooded plants during recovery phase. Catalase and superoxide dismutase activities decreased in leaves of flooded plants during early and later phase of post-flooding, respectively. It seems that post-flooding-induced enhancement of leaf senescence is associated with an increased lipid peroxidation coupled with reduced catalase and superoxide dismutase activities. Key words: Catalase; Flooding; Nicotiana tabacurn; Senescence; Superoxide dismutase
1. Introduction
Reactive oxygen species, such as superoxide anion and hydrogen peroxide, can initiate lipid peroxidation [1]. Measurements of malondial- dehyde (MDA) in tissues are frequently used to indicate lipid peroxidation and indirectly to in- dicate the function of oxygen species. Plants possess antioxidative enzymes, superoxide dis- mutase (SOD) and catalase, to remove superoxide anion and hydrogen peroxide, respectively [1-31.
* Corresponding author.
Abbreviations: EDTA, etbylenediamine tetraacetic acid; MDA, malondialdehyde; SOD, superoxide dismutase.
It has been shown that postanoxic injury of soy- bean roots and in rhizomes of the anoxia-sensitive species, Iris germanica, after anoxic stress were associated with increased superoxide radical production capacity and lipid peroxidation coupled with a reduced SOD activity [4-6]. We have found that leaf senescence of tobacco was enhanced during the recovery phase after a short duration (2 days) of flooding. Leaf senescence has been shown to be associated with increasing levels of lipid peroxidation [3,7,81. However, it is not known whether oxidative damage occurs in leaves during post-flooding. In the study now reported, activities of some enzymes associated with activa- ted oxygen metabolism, as well as the process of lipid peroxidation were monitored in order to 0168-9452/94/$06.00 © 1994 Elsevier Scientific Publishers Ireland Ltd. All rights reserved.
42 14~ P. Hurng, C.H. Kao / Plant Sci. 96 (1994) 41-44
follow the pattern of senescence in leaves of tobac- co plants which had been subjected to 2 previous days of flooding.
2. Materials and methods
Seedlings of Nicotiana tabacum (cv. Speight G- 70) were selected 30 days after sowing the seeds in plastic trays and grown in a greenhouse thereafter, each in a small plastic bag (25 cm2). After 30 days, the seedlings were again selected for unifor- mity and transplanted to pots (5 dm 2) containing sandy loam. Each pot contained a plant and received 30 g compound fertilizer (N-P2Os-K20, 7-21-21). Each treatment had 4 plants. Pots were then placed in the field.
Flooding treatment started at 40 days after transplanting when tobacco plants had 15-17 leaves. Tobacco plants were flooded for 2 days by maintaining the water level at 1 cm above the soil surface. Excess water was drained during a 6-day recovery period. Unflooded control plants were allowed to receive the optimum amount of water. At the times indicated, the lowest leaves were col- lected for chemical analysis and enzyme assays.
Chlorophyll levels were determined according to Wintermans and De Mots [9]. MDA was extracted with 5% (w/v) trichloroacetic acid and determined according to Heath and Packer [10]. For enzyme extraction, leaf tissues were homogenized in an ice bath with a mortar and pestle and 0.1 M sodium phosphate buffer (pH 6.8) containing 0.1% Triton X-100 and 0.05 g Polyclar and centrifuged at 12 000 × g, 4°C for 20 min. The supernant frac- tions were used for enzyme assays. Catalase activ- ity was assayed by measuring the initial rate of disappearance of H202 [11]. Three ml of catalase assay reaction mixture contained 0.1 M sodium phosphate buffer (pH 7.0), 2 mM H202 and 0.1 ml enzyme extract. The decrease in H202 was followed as the decline in optical density at 240 nm, and activity was calculated using the extinction coefficient (40.0 mM -= cm -~ at 240 nm) for H202 [11]. The activity was expressed in units, where one unit of catalase converts 1 nmol of H202 in 1 min. SOD activity was assayed by the method of Paoletti et al. [12]. The assay mixture contained 1.6 ml triethanolamine-diethanolamine
buffer (0.1 M, pH 7.4), 0.08 ml N A D H (reduced form nicotinamide adenine dinucleotide, 7.5 mM), 0.05 ml EDTA/MnCI2 (100 mM/50 mM, pH 7.0), 0.1 ml mercaptoethanol (10 mM) and 0.1 ml enzyme extract. The changes in optical density at 340 nm were recorded for 10 min. One unit of SOD was expressed as the amount of SOD capable of inhibiting by 50% the rate of N A D H oxidation observed in the control.
3. Results and discussion
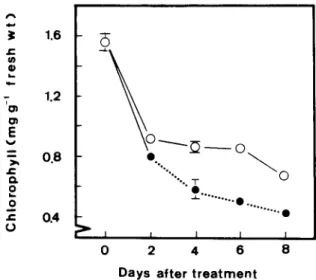
The senescence of tobacco leaves was followed by measuring the decrease in the level of chlorophyll. Fig. 1 shows the changes in the chlorophyll level in tobacco leaves during flooding and recovery compared with unstressed controls. In control leaves, the decrease of chlorophyll was evident at 2 days after the start of the experiment. The reported changes of the leaves by flooding and during recovery therefore may only apply to leaves which have already started the senescence process. As can been seen from Fig. 1, the chlorophyll level was lower in leaves of tobacco plant flooded for 2 days than the corresponding controls during
2
J= 1 . 6 o h , . - 1 . 2 E "- 0 . 8 • © ~ °. n "', %. T 0 0 " O . " ° ' " ' ° ' " • . . . o ° , . ° . . " 0 . 4 ' • J ~ I I I 0 2 4 6 8 D a y s a f t e r t r e a t m e n tFig. I. Changes in chlorophyll levels in tobacco leaves during flooding and recovery (.--) compared with unstressed controls. Vertical bars represent standard errors (n = 4). O, Control; 0, flooded. Only those standard errors larger than the symbol size
I¥.P. Hurng, C.H. Kao/Plant Sci. 96 (1994) 41-44 43 3 2 A 2 8 P • 2 4 '0) o E 2O C a • 1 6 1 2
q
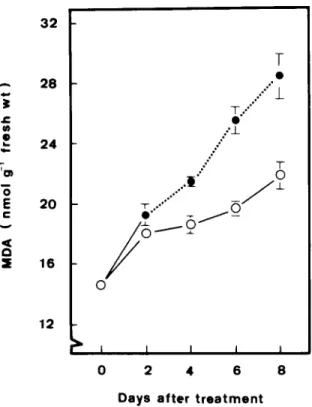
°°'°" _1_ T ."" • " .:J_ - T...
."" © I I I I I 0 2 4 6 8 D a y s a f t e r t r e a t m e n tFig. 2. Changes in malondialdehyde (MDA) levels in tobacco leaves during flooding and recovery (...) compared with unstressed controls. Vertical bars represent standard errors (n = 4). O, Control; e, flooded. Only those standard errors
larger than the symbol size are shown.
A ]¢ 4 >_ j= • ";ca 2 el (J ~ 0 T O O °'°°°°°° i --°"°"°'°.o° °°.°o°°°°~ • I I I I I 0 2 4 6 8 D a y s a f t e r t r e a t m e n t
Fig. 3. Changes in catalase activity in tobacco leaves during flooding and recovery (...} compared with unstressed controls. Vertical bars represent standard errors (n = 4). O, Control; e, flooded. Only those standard errors larger than the symbol size
are shown.
in leaves o f flooded plants during post-flooding m a y be a reflection o f the decline o f protective enzymes. Likely candidates for these enzymes are S O D and catalase since these dismutate super- oxide and destroy h y d r o g e n peroxide, respectively,
flooding period and recovery period. The results also show that e n h a n c e m e n t o f chlorophyll reduc- tion in leaves o f flooded plants is greater during post-flooding than during flooding.
Postanoxic injury o f soybean roots and in rhizomes o f anoxia-sensitive Iris germanica has been shown to be associated with lipid peroxida- tion [4-6]. Free radical-induced lipid peroxidation is considered to be an i m p o r t a n t m e c h a n i s m o f leaf senescence [3,7,8,13]. Thus, lipid peroxidation, as judged by the level o f M D A , in leaves o f flooded plants during post-flooding is expected to be higher than the c o r r e s p o n d i n g controls. As in- dicated in Fig. 2, this is indeed the case. The increase o f the M D A level in leaves o f flooded plants is closely associated with the decrease o f the chlorophyll level during post-flooding.
The striking increase in lipid peroxidation seen
~ 18 .m t- • 0) -~" 16 O (/) e- -, 14
i
°° ... °'°'°'°°°°'°'°°" • -°'°°"°'° • | I I I I 0 2 4 6 8 D a y s a f t e r t r e a t m e n tFig. 4. Changes in superoxide dismutase (SOD) activity in tobacco leaves during flooding and recovery (---) compared with unstressed controls. Vertical bars represent standard errors (n = 4). O, Control; e, flooded. Only those standard er-
44 w.P. Hurng, C.H. Kao / Plant Sci. 96 (1994) 41-44 w h i c h a r e k n o w n to b e i n v o l v e d in the e v o l u t i o n o f t h e h i g h l y reactive a n d toxic h y d r o x y l r a d i c a l s , a p o t e n t i n i t i a t o r o f l i p i d p e r o x i d a t i o n [14]. Figs. 3 a n d 4 s h o w t h e c h a n g e s o f c a t a l a s e a n d S O D activ- ities in leaves o f t o b a c c o p l a n t s d u r i n g f l o o d i n g a n d r e c o v e r y c o m p a r e d w i t h u n s t r e s s e d c o n t r o l s d u r i n g the first 4 d a y s o f p o s t - f l o o d i n g . S O D activ- ity in leaves o f f l o o d e d p l a n t s w a s s i m i l a r to t h a t in the u n s t r e s s e d c o n t r o l s d u r i n g t h e first 2 d a y s o f p o s t - f l o o d i n g , b u t was l o w e r t h e r e a f t e r . T h e rela- t i o n s h i p b e t w e e n S O D a n d c a t a l a s e activities a n d lipid p e r o x i d a t i o n h a s b e e n d e m o n s t r a t e d u n d e r d r o u g h t , p o s t - a n o x i a a n d s a l i n i t y c o n d i t i o n s [ 5 , 6 , 1 5 - 1 7 ] . I n c r e a s e d lipid p e r o x i d a t i o n c o u p l e d w i t h r e d u c e d S O D a n d c a t a l a s e activities in leaves d u r i n g p o s t - f l o o d i n g h a s n o t p r e v i o u s l y b e e n d e m o n s t r a t e d . I n t h e p r e s e n t i n v e s t i g a t i o n , we p r o v i d e t h e e v i d e n c e t h a t o x i d a t i v e d a m a g e o c c u r s d u r i n g p o s t - f l o o d i n g . I n c o n c l u s i o n , we f o u n d t h a t p o s t - f l o o d i n g in- d u c e d e n h a n c e m e n t o f l e a f s e n e s c e n c e c o i n c i d e d w i t h a n e l e v a t e d l i p i d p e r o x i d a t i o n a n d r e d u c e d S O D a n d c a t a l a s e activities i n t o b a c c o leaves. 4. References
1 W.W. Kellog and I. Fridovich, Superoxide, hydrogen per- oxide and singlet oxygen in lipid production by a xanthine oxidase system. J. Biol. Chem., 250 (1975) 8812-8817. 2 I. Fridovich, Superoxide dismutases. Annu. Rev. Bio-
chem., 44 (1975) 147-159.
3 R. Dhindsa, P. Plumb-Dhindsa and T. Thorpe, Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 126 (1981) 93-101.
4 T.T. VanToai and C.S. Bolles, Postanoxic injury in soy- bean (Glycine max) seedlings. Plant Physiol., 97 (1991) 588-592.
5 M.I.S. Hunter, A.M. Hetherington and R.M.M. Crawford, Lipid peroxidation - - a factor in anoxia intolerance in Iris species? Phytochemistry, 22 (1983)
1145-1147.
6 L.S. Monk, K.V. Fagerstedt and R.M.M. Crawford, Oxy- gen toxicity and superoxide dismutase as an antioxidant in physiological stress. Physiol. Plant., 76 (1989) 456-459. 7 R.S. Dhindsa, P.L. Plumb-Dhindsa and D.M. Reid, Leaf senescence and lipid peroxidation: Effect of some phytochromones, and scavengers of free radicals and singlet oxygen. Physiol. Plant., 56 (1982)453-457. 8 K.J. Kunert and M. Ederer, Leaf aging and lipid peroxi-
dation: The role of the antioxidants vitamin C and E. Physiol. Plant., 65 (1985) 85-88.
9 J.F.G.M. Wintermans and A. De Mots, Spectrophoto- metric characteristics of chlorophylls a and b and their pheophytins in ethanol. Biochim. Biophys. Acta, 109 (1965) 448-452.
10 R.L. Heath and L. Packer, Photooxidation in isolated chloroplasts. 1. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125 (1968) 189-198.
11 M. Kato and S. Shimizu, Chlorophyll metabolism in higher plants VII. Chlorophyll degradation in senescing tobacco leaves; phenolic-dependent peroxidative degrada- tion. Can. J. Bot., 65 (1987) 729-735.
12 F. Paoletti, D. Aldinucci, A. Mocali and A. Cparrini, A sensitive spectrophotometric method for the determina- tion of superoxide activity in tissue extracts, Anal. Biochem., 154 (1986) 536-541.
13 A. Upadhyaya, D. Sankhla, T.D. Davis, N. Sankhla and B.N. Smith, Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J. Plant Physiol., 121 (1985)453-461.
14 C. Bowler, M. Van Montagu and D. Inze, Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol., 43 (1992) 83-116.
15 S.R. Chowdhury and M.A. Choudhuri, Hydrogen perox- ide metabolism as an index of water tolerance in jute. Physiol. Plant., 65 (1985) 503-507.
16 J.J. lrigoyen, D.W. Emerich and M. Sanchez-Diaz, Alfal- fa leaf senescence induced by drought stress: Photosynthe- sis, hydrogen peroxide metabolism, lipid peroxidation and ethylene evolution. Physiol. Plant., 84 (1992) 67-72. 17 M. Gong, N.C. Ding, Z.Y. He and Y.L. Liu, Correlation
between lipid peroxidation damage and ultrastructural changes of mesophyll cells in barley and wheat seedlings during salt stress. Acta Bot. Sin., 31 (1989) 841-846.