Nitric oxide counteracts the senescence of rice leaves induced by
hydrogen peroxide
Kuo Tung HUNG and Ching Huei KAO*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (Received February 17, 2004; Accepted July 26, 2004)
Abstract. In the present study, we evaluate the protective effect of nitric oxide (NO) against the senescence of rice leaves promoted by hydrogen peroxide (H2O2). Senescence of rice leaves was determined by decreases in protein content. H2O2 treatment resulted in (1) increases in leaf H2O2 content, (2) induction of leaf senescence, (3) increases in lipid peroxidation, (4) decreases in ascorbic acid (AsA) and reduced form glutathione (GSH) contents, and (5) increases in antioxidative enzyme activities (ascorbate peroxidase, glutathione reductase, and peroxidase). NO donors [N-tert-butyl-α-phenylnitrone (PBN), sodium nitroprusside, 3-morpholinosydonimine, and AsA + NaNO2] were ef-fective in reducing H2O2-induced leaf senescence. PBN prevented H2O2-increased H2O2 content, H2O2-induced lipid peroxidation, H2O2-decreased AsA and GSH contents, and H2O2-increased antioxidative enzyme activities. The pro-tective effect of PBN on H2O2-promoted senescence, H2O2-increased H2O2 content and lipid peroxidation, H2O2 -decreased AsA and GSH contents, and H2O2-increased antioxidative enzyme activities was reversed by 2-(4-carboxy-2-phenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide, a NO-specific scavenger, suggesting that the protective effect of PBN is attributable to the NO released. Reduction of H2O2-induced senescence by NO in rice leaves is most likely mediated through its ability to scavenge H2O2.
Keywords: Hydrogen peroxide; Lipid peroxidation; Nitric oxide; Oryza sativa.
Introduction
Hydrogen peroxide (H2O2) is a constituent of oxidative plant metabolism and is itself an active oxygen species (AOS). H2O2 can also react with superoxide radicals to form more active hydroxyl radicals in the presence of trace amounts of Fe or Cu (Van Breusegem et al., 2001). The hydroxyl radicals initiate self-propagating reactions lead-ing to peroxidation of membrane lipids and destruction of proteins (Halliwell and Gutteridge, 1989). H2O2 has been shown to promote leaf senescence (Parida et al., 1978; Mondal and Choudhuri, 1981; Sarkar and Choudhuri, 1981; Begam and Choudhuri, 1992; Lin and Kao, 1998), and in-duction of senescence is accompanied by an increase in endogenous H2O2 content (Mondal and Choudhuri, 1981). Lipid peroxidation is considered an important mechanism of leaf senescence (Thompson et al., 1987; Strother, 1988). The peroxidation of lipids can be initiated by active oxy-gen species (Thompson et al., 1987; Halliwell and Gutteridge, 1989). Thus, H2O2-induced leaf senescence is mediated, at least in part, through lipid peroxidation.
In mammalian systems, nitric oxide (NO), a bioactive molecule, is produced mainly from L-arginine by NO syn-thase (NOS). However, in plants neither the protein nor the appropriate gene for NOS activity have been detected (Lamattina et al., 2003). Recent work demonstrates that
*Corresponding author. Fax: +886-2-23620879; E-mail: [email protected]
the mammalian NOS antibodies recognize many NOS-unre-lated plant proteins (Butt et al., 2003), suggesting that in-ferring the presence of plant NOS using the immunological technique may be inappropriate. These results led to speculation that the plant NOS-like enzyme could be struc-turally different from the mammalian NOS. Recently, a pathogen-inducible plant NOS has been identified as a vari-ant P protein of the mitochondrial glycine decarboxylase complex (Chandok et al., 2003; Wendehenne et al., 2003). Additionally, Guo et al. (2003) reported the presence of an NOS gene (AtNOS1) in Arabidopsis. AtNOS1 turned out to be a protein very similar to a group of bacterial pro-teins with putative GTP-binding or GTPase domains. Nei-ther the variant P protein nor the purified AtNOS1 protein had sequence similarities to any mammalian NOS. It is in-creasingly evident that plant nitrate reductase also cata-lyzes a NAD(P)H-dependent reduction of nitrite to NO (Yamasaki et al., 1999; Rockel et al., 2002, Sakihama et al., 2002; Lamattina et al., 2003). Evidence is mounting that NO acts as an important messenger in plant physiological processes, including growth, development, and defense responses (Noritake et al., 1996; Gouvea et al., 1997; Dangl, 1998; Beligni and Lamattina, 2000, 2001; Neill et al., 2002; Pagnussat et al., 2002; Jih et al., 2003; Zhao et al., 2004). NO has been shown capable of counteracting the toxicity of paraquat and diquat, which are known to generate su-peroxide radicals, in potato leaves (Beligni and Lamattina, 1999a,b; 2002) and block H2O2 production induced by jasmonic acid in tomato leaves (Orozco-Cárdenas and Ryan, 2002). NO seems to be a potent antioxidant, and its
ability to directly scavenge AOS may be behind its action (Beligni and Lamattina, 2002).
In rice leaves, we have shown that NO counteracts oxi-dative stress induced by paraquat, dehydration, and poly-ethylene glycol (Cheng et al., 2002; Hung et al., 2002). More recently, we have shown that the promotion of rice leaf senescence by abscisic acid and methyl jasmonate, both of which induce H2O2 production and lipid peroxidation, can be counteracted by NO donors (Hung and Kao, 2003, 2004). In the present investigation, we ex-amined the effect of NO on the H2O2-induced senescence of rice leaves.
Materials and Methods
Plant Material and Chemicals
Rice (Oryza sativa L., cv. Taichung Native 1) was ster-ilized with 2.5% sodium hypochlorite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dishes with wetted filter paper at 37°C under dark conditions. After a 48-h incubation, uni-formly germinated seeds were selected and cultivated in a 500 mL beaker containing half-strength Kimura B solution as described previously (Chu and Lee, 1989). The hydro-ponically cultivated seedlings were grown for 12 days in a Phytotron (Agriculture Experimental Station, National Taiwan University, Taipei, Taiwan) with natural light 30°C day (12 h)/25°C night (12 h) and 90% relative humidity. The apical 3 cm of the third leaf was used in all experiments. A group of ten segments was floated in a Petri dish containing 10 mL of test solution. Incubation was carried out at 27°C in the dark.
Test solutions included H2O2, NO donors, and a NO s c a v e n g e r [ 2 ( 4 c a r b o x y 2 p h e n y l ) 4 , 4 , 5 , 5 -tetramethylimidazoline-1-oxyl-3-oxide, c-PTIO]. N-tert-bu-tyl-α-phenylnitrone (PBN), 3-morpholino-sydonimine (SIN-1), and sodium nitroprusside (SNP) were used as NO donors. We also used a solution containing ascorbic acid (AsA) and NaNO2 as another NO donor. All chemicals were purchased from Sigma Co. (St. Louis, MO, USA).
D e t e r m i n a t i o n s o f P ro t e i n , H2O2, L ip i d peroxidation, GSH, and AsA
Chlorophyll was determined according to Wintermans and De Mots (1965) after extraction in 96% (v/v) ethanol. For protein extraction, leaf segments were homogenized in 50 mMsodium phosphate buffer (pH 6.8). The extracts were centrifuged at 17,600 g for 20 min, and the superna-tants were used for determination of protein by the method of Bradford (1976) and antioxidant enzyme activities. The H2O2 content was measured colorimetrically as described by Jana and Choudhuri (1981). H2O2 was extracted by ho-mogenizing leaf tissue with phosphate buffer (50 mM, pH 6.5) containing 1 mMhydroxylamine. The homogenate was centrifuged at 6,000 g for 25 min. To determine H2O2 content, extracted solution was mixed with 0.1% (v/v) ti-tanium chloride in 20% (v/v) H2SO4. The mixture was then
centrifuged at 6,000 g for 25 min. The absorbance was mea-sured at 410 nm. The H2O2 content was calculated using the extinction coefficient 0.28 µmol-1 cm-1. Lipid peroxidation in leaf tissue was determined by measuring thiobarbituric acid reactive substances (TBARS). The leaf tissue was extracted with 5% (w/v) trichloroacetic acid and determined according Heath and Packer (1968). GSH in 5% (w/v) sulfosalicylic acid extract and AsA in 5% (w/v) trichloroacetic acid extract were determined as described by Smith (1985) and Laws et al. (1983), respectively.
Enzyme Assays
Peroxidase (POD) activity was measured using a modi-fication of the procedure of MacAdam et al. (1992). Ac-tivity was calculated using the extinction coefficient (26.6 mM-1 cm-1 at 470 nm) for tetraguaiacol. Catalase (CAT) activity was assayed by measuring the initial rate of dis-appearance of H2O2 (Kato and Shimizu, 1987). The decrease in H2O2 was followed as the decline in absorbance at 240 nm, and activity was calculated using the extinction coef-ficient (40 mM-1 cm-1 at 240 nm) for H
2O2 (Kato and Shimizu, 1987). Superoxide dismutase (SOD) was determined ac-cording to Paoletti et al. (1986). Ascorbate peroxidase (APOD) was determined according to Nakano and Asada (1981). The decrease in AsA concentration was followed as the decline in optical density at 290 nm and activity was calculated using the extinction coefficient (2.8 mM-1 cm-1 at 290 nm) for AsA. Glutathione reductase (GR) was de-termined by the method of Foster and Hess (1980). One unit of activity for CAT, POD, SOD, APOD, and GR was defined as the amount of enzyme which degraded 1 µmol H2O2 per min, caused the formation of 1 µmol tetraguaiacol per min, inhibited 50% the rate of NADH oxidation ob-served in control, degraded 1 µmol of AsA per min, and decreased 1 A340 per min, respectively.
Statistical Analysis
Statistical differences between measurements (n = 4) on different treatments or on different times were analyzed following Duncan’s multiple range test.
Results and Discussion
The most obvious character of leaf senescence is yellowing. Chlorophyll loss has long been considered the principal criterion of senescence. The protein breakdown that occurs during leaf senescence has been recognized since the earliest studies performed. We have shown that protein breakdown precedes chlorophyll loss during rice leaf senescence (Kao, 1980). Thus, senescence of rice leaves in the present investigation was followed by mea-suring the decrease of protein. The changes in protein and TBARS content in detached rice leaves treated with 10 mM H2O2 in the dark are shown in Figures 1A and 1B. The decease in protein and increase in TBARS were evi-dent at 2 days after H2O2 treatment. Clearly, H2O2 is effec-tive in promoting the senescence of rice leaves. To be sure that the described effect of H2O2 on leaf senescence
was related to an increase in the leaf H2O2 content, H2O2 concentration was determined in detached rice leaves treated with either water or 10 mM H2O2 (Figure 1C). H2O2 content in control leaves remained unchanged during 3 days of incubation in the dark. The increase in H2O2 con-tent in H2O2-treated detached rice leaves was clearly evi-dent at 2 days after treatment.
H2O2 treatment resulted in a marked increase in TBARS content, indicating that H2O2 brings about lipid peroxidation (Figure 1B). In previous work, when free radi-cal scavengers such as AsA, GSH, sodium benzoate, and thiourea were used together with H2O2, it was found that
they prevented H2O2-promoted senescence and lipid peroxidation (Lin and Kao, 1998). All these results sug-gest that H2O2 treatment caused an oxidative stress.
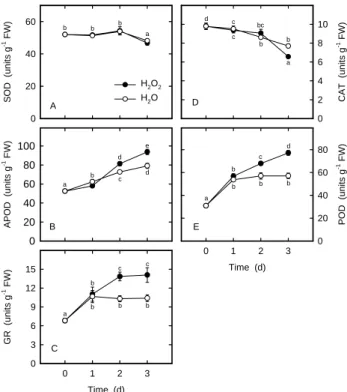
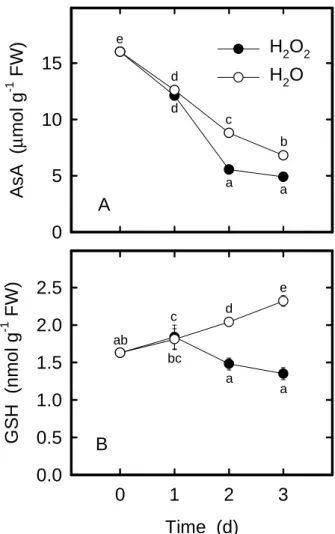
Plant cells are equipped with several AOS detoxifying enzymes and antioxidants to protect them against oxida-tive damage. Antioxidant enzymes include SOD, APOD, GR, CAT, and POD (Foyer et al., 1997). AsA and GSH are two main water-soluble antioxidants (Foyer et al., 1997). The striking increase in lipid peroxidation seen in rice leaves treated with H2O2 (Figure 1B) may reflect the change in the activities of antioxidative enzymes and in the con-tents of antioxidants. It was observed that, H2O2-treated rice leaves had higher activities of APOD (Figure 2B), GR (Figure 2C), and POD (Figure 2E) than the controls at 2 days after dark incubation. Increased CAT activity by H2O2 was observed at 3 days after treatment (Figure 2D). However, H2O2 had no effect on SOD activity in rice leaves (Figure 2A). Treatment with H2O2 significantly decreased AsA and GSH contents compared with the control leaves, with the increase occurring 2 day after treatment (Figures 3A and 3B). The increased activities of APOD, GR, and POD and the decreased contents of AsA and GSH in rice leaves in response to H2O2 further suggest a strong in-duction of oxidative stress.
Figure 1B and 1C show that no changes in TBARS or H2O2 contents were observed in the control leaves. Lipid peroxidation and dark-induced senescence of rice leaves appear to have no direct relationship under dark
Prot ein (m g g -1 FW) 0 10 20 30 40 50 T BAR S (nm o l g -1 FW) 0 10 20 30 40 50 H2O2 H2O Time (d) 0 1 2 3 H2 O2 (µ mo l g -1 FW ) 0 10 20 30 40 50 d cd cd c b b a a a a a a b c c a a a a a b A B C
Figure 1. Changes in the contents of protein (A), MDA (B), and H2O2 (C) in rice leaves treated with H2O2. Detached rice leaves were treated with either water or 10 mMH2O2 in the dark. Values with the same letter are not significantly different at P < 0.05, according to Duncan’s multiple range test.
S O D (u ni ts g -1 FW) 0 20 40 60 H2O2 H2O APO D ( uni ts g -1 FW ) 0 20 40 60 80 100 Time (d) 0 1 2 3 G R (uni ts g -1 FW) 0 3 6 9 12 15 CA T (u ni ts g -1 FW) 0 2 4 6 8 10 Time (d) 0 1 2 3 PO D ( uni ts g -1 FW ) 0 20 40 60 80 a b b b b a c d d e a b b bc c c d a b b b b c d a b b b b c c A B C E D
Figure 2. Changes in the activities of SOD (A), APOD (B), GR (C), CAT (D) and POD (E) in rice leaves treated with ei-ther water or 10 mMH2O2 in the dark. Values with the same letter are not significantly different at P < 0.05, according to Duncan’s multiple range test.
conditions. The changes in the activities of SOD (Figure 2A), APOD (Figure 2B), GR (Figure 2C), CAT (Figure 2D), and POD (Figure 2E) and the contents of AsA (Figure 3A) and GSH (Figure 3B) which occurred in detached rice leaves after incubation in water may be a manifestation of changes in metabolism associated with excision and con-sequent senescence.
In the present investigation, we show that the promo-tion of senescence in detached rice leaves by H2O2 is as-sociated with lipid peroxidation or oxidative stress. NO is known to counteract oxidative stress in plants (Beligni and Lamattina, 1999a,b; 2002; Beligni et al., 2002; Cheng et al., 2002; Hung et al., 2002; Hung and Kao, 2003, 2004). Thus, it is of great interest to know whether the protective power of NO is also active in the H2O2-promoted senescence of rice leaves.
To study the role of NO, various donors were employed, with the assumption that they release NO. Typically, NO is applied to plant tissues via a NO-donor, that is, a mol-ecule that will generate NO, sometimes after passage into
AsA
(µ
mol
g
-1FW
)
0
5
10
15
Time (d)
0
1
2
3
GS
H
(nmol g
-1FW
)
0.0
0.5
1.0
1.5
2.0
2.5
H
2O
2H
2O
e d d c b a a a a ab bc c d eA
B
Table 1. Effect of NO donors on protein content in rice leaves treated with H2O2. The concentrations of H2O2, PBN, SIN-1, SNP, AsA, and NaNO2 were 10 mM, 100 µM, 100 µM, 100 µM, 100 µM, and 200 µM, respectively. Protein content was determined 2 days after treatment in the dark. Values with the same letter are not significantly different at P < 0.05, according to Duncan’s multiple range test.
Treatment Protein (mg g-1 FW) H2O 40.9 ± 0.37 (c) H2O2 29.8 ± 0.29 (a) H2O2 + PBN 36.6 ± 0.54 (b) H2O2 + SIN-1 34.5 ± 1.1 (b) H2O2 + SNP 36.8 ± 0.67 (b) H2O2 + AsA+ NaNO2 35.8 ± 0.78 (b) Figure 3. Changes in the contents of AsA (A) and GSH (B) in
rice leaves treated with either water or 10 mMH2O2 in the dark. Values with the same letter are not significantly different at P < 0.05, according to Duncan’s multiple range test.
cells. Chamulitrat et al. (1993) postulated that PBN can undergo oxidative decomposition to release NO. SIN-1 is known to generate both superoxide anion and nitric oxide which spontaneously form peroxynitrite (Spiecker et al., 1993). Peroxynitrite has been shown to react with H2O2 to yield nitrite ion and oxygen (Martinez et al., 2000). This reaction has been suggested to be the mechanism of NO cytoprotective actions in animals (Wink et al., 1993). NO is released from SNP mainly by decomposition of pure so-lutions due to photochemical reactions or by the various reducing metabolites, including thiols, in biological or-ganelles (Rochelle et al., 1994; Rao and Cederbaum, 1995; Ioannidis et al., 1996). As shown in Table 1, all NO donors, such as PBN, SIN-1, SNP, and a mixture of AsA and NaNO2, are effective at inhibiting H2O2-promoted senescence of rice leaves.
To investigate whether the protective effect induced by PBN treatment was the result of the production of NO, 100 µM c-PTIO, a NO-specific scavenger, was applied along with 100 µM PBN. The effect of PBN on H2O2-promoted protein loss and H2O2-induced increase in lipid peroxidation and H2O2 content could be reversed by c-PTIO (Table 2). We also observed that PBN counteracted H2O2-induced increases in antioxidant enzyme activities, and c-PTIO re-versed the effect of PBN-decreased enzyme activities (Table 3). Furthermore, the effect of PBN on H2O2-decreased AsA and GSH contents could be reversed by c-PTIO (Table 4). Clearly, the effect of NO donor PBN would then be attrib-utable to NO released.
It has been shown that NO is a potent antioxidant in plants and that its action may, at least in part, be explained by its ability to directly scavenge AOS, H2O2 and O2 •-(Beligni and Lamattina, 2002). If NO acts as an antioxidant, it may reduce H2O2 content in H2O2-treated rice leaves. Since NO reduces H2O2-increased H2O2 content (Table 2), it appears that it indeed has the ability to scavenge AOS. Orozco-Cárdenas and Ryan (2002) also reported that NO blocked the H2O2 production that was induced by jasmonic acid. We also observed that NO donors blocked abscisic acid- and methyl jasmonate-induced H2O2 produc-tion in rice leaves (Hung and Kao, 2003, 2004).
In the present investigation, we found that NO reduced H2O2-increased lipid peroxidation (Table 2) and antioxidant enzyme activities (Table 3) in rice leaves. These results are in agreement with our previous work, in which we dem-onstrated that NO counteracted paraquat-increased lipid peroxidation and antioxidant enzyme activities (Hung et al., 2002). Because lipid peroxidation and the increase in anti-oxidant enzyme activities are the consequence of AOS (Thompson et al., 1987) and NO acts as an AOS scavenger, the reduction of lipid peroxidation and antioxidant enzyme activities could be a result of low levels of H2O2 in rice leaves treated with NO and H2O2 (Table 1). The fact that NO counteracts H2O2-decreased AsA and GSH (Figures 3A and 3B) should result in an increased capacity to scav-enge H2O2 in rice leaves treated with NO and H2O2 com-pared to rice leaves treated with H2O2 alone and might account in part for the lower contents of H2O2 observed in rice leaves treated with both NO and H2O2 (Table 2).
Table 2. Effect of PBN on the contents of protein, MDA, and H2O2 in H2O2-treated rice leaves in the presence or absence of c-PTIO. The concentrations of H2O2, PBN, and c-PTIO were 10 mM, 100 µM, and 100 µM, respectively. Protein, MDA, and H2O2 contents were determined 2 days after treatment in the dark. Values with the same letter are not significantly different at P < 0.05, according to Duncan’s multiple range test.
Treatment Protein (mg g-1 FW) TBARS (nmol g-1 FW) H
2O2 (µmol g -1 FW) H2O 41.9 ± 0.68 (c) 26.0 ± 0.73 (a) 35.1 ± 0.48 (b) H2O2 31.3 ± 0.41 (a) 36.0 ± 0.44 (b) 42.1 ± 0.44 (d) H2O2 + PBN 36.8 ± 0.41 (b) 27.5 ± 0.35 (a) 32.4 ± 0.30 (a) H2O2 + PBN + c-PTIO 31.9 ± 1.0 (a) 38.4 ± 0.97 (b) 39.3 ± 0.79 (c)
Table 3. Effect of PBN on the activities of antioxidant enzymes in H2O2-treated rice leaves in the presence or absence of c-PTIO. The concentrations of H2O2, PBN, and c-PTIO were 10 mM, 100 µM, and 100 µM, respectively. Enzyme activities were deter-mined 2 days after treatment in the dark. Values with the same letter are not significantly different at P < 0.05, according to Duncan’s multiple range test.
Treatment APOD (units g-1 FW) POD (units g-1 FW) GR (units g-1 FW)
H2O 72.7 ± 0.92 (a) 55.5 ± 0.97 (a) 10.6 ± 0.23 (a)
H2O2 81.9 ± 0.98 (c) 74.2 ± 0.86 (c) 13.9 ± 0.14 (c)
H2O2 + PBN 75.3 ± 0.55 (b) 60.9 ± 1.6 (b) 11.4 ± 0.15 (b)
H2O2 + PBN + c-PTIO 82.0 ± 1.2 (c) 74.7 ± 1.7 (c) 14.0 ± 0.17 (c)
N O h as b e e n s h o w n t o ac t as i r o n l i g an d i n haemoproteins (Stamler et al., 1992). Ferrer and Barceló (1999) found that the NO donor SNP (5 mM) and NO (55 µM) itself were able to inhibit POD activity in the xylem of Zinnia elegans. Clark et al. (2000) also demonstrated that NO donor (0.8 mM) inhibited the activities of tobacco CAT and APOD, heme-containing enzymes. However, our re-sults show that PBN alone had no effect on the lipid peroxidation and activities of POD and APOD in rice leaves (data not shown). Thus, the reduction of H2O2-induced increase in antioxidant enzyme activities by NO is not likely due to a direct NO-mediated inhibition of the enzymes. Why is PBN able to counteract H2O2-induced senescence, but not able to inhibit the activities of POD, CAT, and APOD in rice leaves? In the present investigation, we used a µM concentration range of PBN and did not measure the concentration of NO released by PBN in rice leaves. It is possible that the concentration of NO released by PBN was high enough to scavenge H2O2, but not sufficient to in-hibit the activities of heme-containing enzymes.
In conclusion, the results presented in this paper pro-vide epro-vidence that NO acts as an antioxidant in inhibiting H2O2-promoted rice leaf senescence.
Acknowledgements. This work was supported by Grant NSC 91-2313-B-002-364 from the National Science Council of the Republic of China.
Literature Cited
Begam, H.H. and M.A. Choudhuri. 1992. H2O2 metabolism dur-ing senescence of two submerged angiosperms Hydrilla and Ottelia: Changes in enzyme activities in light and darkness. Biochem. Physiol. Pflanzen 188: 105-115.
Table 4. Effect of PBN on the contents of AsA and GSH in H2O2-treated rice leaves in the presence or absence of c-PTIO. The concentrations of H2O2, PBN, and c-PTIO were 10 mM, 100 µM, and 100 µM, respectively. AsA and GSH contents were determined 2 days after treatment in the dark. Values with the same letter are not significantly different at P < 0.05, ac-cording to Duncan’s multiple range test.
Treatment AsA (µmol g-1 FW) GSH (nmol g-1 FW)
H2O 8.53 ± 0.24 (c) 1.93 ± 0.01 (d)
H2O2 5.86 ± 0.10 (a) 1.40 ± 0.02 (a) H2O2 + PBN 7.65 ± 0.19 (b) 1.70 ± 0.02 (c) H2O2 + PBN + c-PTIO 5.91 ± 0.15 (a) 1.54 ± 0.01 (b)
Beligni, M.V. and L. Lamattina. 1999a. Nitric oxide counteracts cytotoxic processes mediated by reactive oxygen species in plant tissues. Planta 208: 337-344.
Beligni, M.V. and L. Lamattina. 1999b. Nitric oxide protects against cellular damage produced by methyl viologen her-bicides in potato plants. Nitric Oxide Biol. Chem. 3: 199-208.
Beligni, M.V. and L. Lamattina. 2000. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocot-yls elongation, three light-inducible responses in plants. Planta 210: 215-221.
Beligni, M.V. and L. Lamattina. 2001. Nitric oxide in plants: the history is just beginning. Plant Cell Environ. 24: 267-278. Beligni, M.V. and L. Lamattina. 2002. Nitric oxide interferes with
plant photo-oxidative stress by detoxifying reactive oxy-gen species. Plant Cell Environ. 25: 737-748.
Beligni, M.V., A. Fath, P.C. Bethake, L. Lamattina, and R.L. Jones. 2002. Nitric oxide acts as an antioxidant and delays programmed cell death in barley aleurone layers. Plant Physiol. 129: 1462-1650.
Bradford, M.M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248-254.
Butt, Y.K.-C., J.H.-K. Lum, and S.C.-L. Lo. 2003. Proteomic identification of plant proteins probed by mammalian ni-tric oxide synthase antibodies. Planta 216: 762-771. Chamulitrat, W., S.J. Jordan, R.P. Mason, K. Saito, and R.
Gutler. 1993. Nitric oxide formation during light-induced decomposition phenyl N-tert-butylnitrone. J. Biol. Chem. 268: 11520-11527.
Chandok, M.R., A.J. Ytterberg, K.J. van Wijk, and D.F. Klessig. 2003. The pathogen-inducible nitric oxide synthase (iNOS) in plants is a variant of the P protein of the glycine decar-boxylase complex. Cell 113: 469-482.
Cheng, F.-Y., S.-Y. Hsu, and C.H. Kao. 2002. Nitric oxide coun-teracts the senescence of detached rice leaves induced by dehydration and polyethylene glycol but not by sorbitol. Plant Growth Regul. 38: 265-272.
Chu, C. and T.M. Lee. 1989. The relationship between ethyl-ene biosynthesis and chilling tolerance in seedlings of rice (Oryza sativa). Bot. Bull. Acad. Sin. 30: 263-273. Clark, D., J. Dunar, D.A. Navarre, and D.F. Klessig. 2000.
Ni-tric oxide inhibition of tobacco catalase and ascorbate peroxidase. Mol. Plant-Microbe Interact. 13: 1380-1384. Dangl, J. 1988. Innate immunity. Plants just say NO to
pathogens. Nature 394: 525-527.
Ferrer, M.A. and A.R. Barceló. 1999. Differential effects of ni-tric oxide on peroxidase and H2O2 production by the xy-lem of Zinnia elegans. Plant Cell Environ. 22: 891-897. Foster, J.G. and J.L. Hess. 1980. Responses of superoxide
dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol. 66: 482-487.
Foyer, C.H., H. Lopez-Delgado, J.F. Dat, and I.M. Scott. 1997. Hydrogen peroxide and glutathione-associated mechanism of acclimatory stress tolerance and signaling. Physiol. Plant. 100: 241-254.
Gouvea, C.M.C.P., J.F. Souza, A.C.N. Magalhaes, and I.S. Martins. 1997. NO-releasing substances that induce growth elongation in maize root segments. Plant Growth Regul.
21: 183-187.
Guo, F.-Q., M. Okamoto, and N.M. Crawford. 2003. Identifi-cation of a plant nitric oxide synthase gene involved in hor-monal signaling. Science 302: 100-103.
Halliwell, B. and J.M.C. Gutteridge. 1989. Free Radicals in Bi-ology and Medicine. Oxford: Clarendon Press.
Heath, R.L. and L. Packer. 1968. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125: 189-198. Hung, K.T. and C.H. Kao. 2003. Nitric oxide counteracts the
senescence of rice leaves induced by abscisic acid. J. Plant Physiol. 160: 871-879.
Hung, K.T. and C.H. Kao. 2004. Nitric oxide acts as an antioxi-dant and delays methyl jasmonate-induced senescence of rice leaves. J. Plant Physiol. 161: 43-52.
Hung, K.T., C.J. Chang, and C.H. Kao. 2002. Paraquat toxicity is reduced by nitric oxide in rice leaves. J. Plant Physiol. 159: 159-166.
Ioannidis, I., M. Bätz, T. Paul, H.-G. Korth, R. Sustamann, and H. de Groot. 1996. Enhanced release of nitric oxide causes increased cytotoxicity of S-nitroso-N-acetyl-DL-penicil-lamine and sodium nitroprusside under hypoxic conditions. Biochem. J. 318: 789-795.
Jana, S. and M.A. Choudhuri. 1981. Glycolate metabolism of three submerged aquatic angiosperm during aging. Aquat. Bot. 12: 345-354.
Jih, P.-J., Y.-C. Chen, and S.T. Jeng. 2003. Involvement of hy-drogen peroxide and nitric oxide in expression of the ipomoelin gene from sweet potato. Plant Physiol. 132: 381-389.
Kao, C.H. 1980. Senescence of rice leaves IV. Influence of benzyladenine on chlorophyll degradation. Plant Cell Physiol. 21: 1255-1262.
Kato, M. and S. Shimizu. 1987. Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing tobacco leaves: Phenolic-dependent peroxidative degradation. Can. J. Bot. 65: 729-735.
Lamattina L., C. García-Mata, M. Graziano, and G. Pagnussat. 2003. Nitric oxide: the versatility of an extensive signal molecule. Annu. Rev. Plant Biol. 54: 109-136.
Laws, M.Y., S.A. Charles, and B. Halliwell. 1983. Glutathione and ascorbic acid in spinach chloroplasts: the effect of hy-drogen peroxide and of paraquat. Biochem. J. 210: 899-903.
Lin, J.N. and C.H. Kao. 1998. Effect of oxidative stress by hy-drogen peroxide on senescence of rice leaves. Bot. Bull. Acad. Sin. 39: 161-165.
MacAdam, J.W., C.J. Nelson, and R.E. Sharp. 1992. Peroxidase activity in the leaf elongation zone of tall fescue. Plant Physiol. 99: 872-878.
Martinez, G.R., P. DiMascio, M.G. Bonini, O. Augusto, K. Briviba, H. Sies, P. Maurer, U. Röthlisberger, S. Herold, and W.H. Koppenol. 2000. Peroxynitrite does not decom-pose to singlet oxygen (1∆gO
2) and nitroxyl (NO -). Proc.
Natl. Acad. Sci. USA 97: 10307-10312.
Mondal, R. and M.A. Choudhuri. 1981. Role of hydrogen per-oxide in senescence of excised leaves of rice and maize. Biochem. Physiol. Pflanzen 176: 700-709.
Nakano, Y. and K. Asada. 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts.
Plant Cell Physiol. 22: 867-880.
Neill, S.J., R. Desikan, A. Clarke, and J.T. Hancock. 2002. Ni-tric oxide is a novel component of abscisic acid signaling in stomatal guard cells. Plant Physiol. 128: 13-16.
Noritake, T., K. Kawakita, and N. Doke. 1996. Nitric oxide in-duces phytoalexin accumulation in potato tuber tissues. Plant Cell Physiol. 37: 113-116.
Orozco-Cárdenas, M. and C.A. Ryan. 2002. Nitric oxide nega-tively modulates wound signaling in tomato plants. Plant Physiol. 130: 487-493.
Pagnussat, G.C., M. Simontacchi, S. Puntarulo, and L. Lamattina. 2002. Nitric oxide is required for root organogenesis. Plant Physiol. 129: 954-956.
Paoletti, F., D. Aldinucci, A. Mocali, and A. Capparini. 1986. A sensitive spectrophotometric method for the determination of superoxide dismutase activity in tissue extracts. Anal. Biochem. 154: 536-541.
Parida, R.K., M. Kar, and D. Mishra. 1978. Enhancement of se-nescence in excised rice leaves by hydrogen peroxide. Can. J. Bot. 56: 2937-2941.
Rao, D.N.R. and A.I. Cederbaum. 1995. Production of nitric ox-ide and other iron-containing metabolites during the reduc-tive metabolism of nitroprusside by microsomes and by thiols. Arch. Biochem. Biophys. 321: 363-371.
Rochelle, L.G., H. Kruszyna, R. Kruszyna, A. Barchowsky, D. E. Wilcox, and R.P. Smith. 1994. Bioactivation of nitroprus-side by porcine endothelial cells. Toxicol. Appl. Pharmcol. 128: 123-128.
Rockel, P., F. Strube, A. Rockel, J. Wildt, and W.M. Kaiser. 2002. Regulation of nitric oxide (NO) production by plant nitrate reductase in vivo and in vitro. J. Exp. Bot. 53: 103-110.
Sakihama, Y., S. Nakamura, and H. Yamasaki. 2002. Nitric oxide production mediated by nitrate reductase in the green alga Chlamydomonas reinharatii: an alternative NO production pathway in photosynthetic organisms. Plant Cell Physiol. 43: 290-297.
Sarkar, U. and M.A. Choudhuri. 1981. Effect of some oxidants
and antioxidants on senescence of isolated leaves of sun-flower with special reference to glycolate content, glyco-late oxidase, and catalase activities. Can. J. Bot. 59: 392-396. Smith, I.K. 1985. Stimulation of glutathione synthesis in photorespiring plants by catalase inhibitors. Plant Physiol. 79: 1044-1047.
Spiecker, M., H. Darius, and J. Meyer. 1993. Synergistic plate-let antiaggregatory effects of the adenylate cyclase activa-tor iloprost and the guanylate cyclase activating agent SIN-1 in vivo. Thromb. Res. 70: 405-415.
Stamler, J.S., D.J. Singel, and J. Loscalzo. 1992. Biochemistry of nitric oxide and its redox-activated forms. Science 258: 1898-1902.
Strother, S. 1988. The role of free radicals in leaf senescence. Gerontology 34: 151-156.
Thompson, J.E., R.L. Legge, and R.F. Barber. 1987. The role of free radical in senescence and wounding. New Phytol. 105: 317-344.
Van Breusegem, F., E. Vranová, J.F. Dat, and D. Inzé. 2001. The role of active oxygen species in plant transduction. Plant Sci. 161: 405-414.
Wendehenne, D., O. Lamotte, and A. Pugin. 2003. Plant iNOS: conquest of the holy grail. Trends Plant Sci. 8: 465-468. Wink, D.A., I. Hanbauer, M.C. Krishna, W. DeGraff, J. Gamson,
and J.B. Mitchell. 1993. Nitric oxide protects against cellu-lar damage and cytotoxicity from reactive oxygen species. Proc. Natl. Acad. Sci. USA 90: 9813-9817.
Wintermans, J.F.G.M. and A. De Mots. 1965. Spectrophoto-metric characteristics of chlorophyll a and b and their pheophytins in ethanol. Biochim. Biophys. Acta 109: 448-453.
Yamasaki, H., Y. Sakihama, and S. Takahashi. 1999. An alterna-tive pathway for nitric oxide production in plants: new fea-tures and an old enzyme. Trends Plant Sci. 4: 128-129. Zhao, L., F. Zhang, J. Guo, Y. Yang, B. Li, and L. Zhang. 2004.
Nitric oxide functions as a signal in salt resistance in the callus from two ecotypes of reed. Plant Physiol. 134: 849-857.
glutathione GSH ascorbic acid AsA ascorbate
peroxidase, glutathione reductase peroxidase [N-tert-butyl-α-phenylnitrone
PBN sodium nitroprusside 3-morpholinosydonimine AsA NaNO2
MDA GSH
AsA