Nitric oxide counteracts the senescence of detached rice leaves induced by
dehydration and polyethylene glycol but not by sorbitol
Fang-yi Cheng, Shih-Yung Hsu and Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China; *Author for correspondence (e-mail: [email protected]; phone: +866-2-23698159; fax: +886-2-23620879)
Received 12 February 2002; accepted in revised form 23 May 2002
Key words: Dehydration, Lipid peroxidation, Oryza sativa, Polyethylene glycol, Sorbitol
Abstract
In the present study, we evaluated the protective effect of nitric oxide (NO) against senescence of rice leaves enhanced by water deficit. Dehydration (DH), polyethylene glycol (PEG) and sorbitol (ST) treatments were used to induce water deficit. Senescence of rice leaves was determined by the decrease of protein content. NO donors [N-tert-butyl-␣-phenylnitrone (PBN), sodium nitroprusside (SNP), 3-morpholinosydnonimine (SIN-1), and ascor-bic acid + NaNO2] were effective in inhibiting senescence of dehydrated and PEG-treated rice leaves, but had no
effect on senescence of ST-treated rice leaves. PEG or DH resulted in a marked increase in malondialdehyde (MDA) content and decrease in superoxide dismutase (SOD) activity, but ST had no effect on MDA content and SOD activity. Treatment with NO donors caused a reduction of PEG- and DH-induced increase in MDA content and decrease in SOD activity. The protective effect of NO donors on promotion of senescence, increase in lipid peroxidation and decrease in SOD activity induced by PEG and DH was reversed by 2-(4-carboxy-2-phenyl)-4,4,5,5- tetra-methylinmidazoline-1-oxyl-3-oxide, a NO specific scavenger, suggesting that the protective effect of NO donors is attributed to NO released. The inhibition of PEG- and DH- enhanced senescence of rice leaves by NO is most likely mediated through increasing SOD activity and decrease in lipid peroxidation.
Abbreviations: Asc – ascorbate, c-PTIO – 2-(4-carboxy-2-phenyl)-4,4,5,5-tetra-
methylinmidazoline-1-oxyl-3-oxide, DH – dehydration, MDA – malondialdehyde, NO – nitric methylinmidazoline-1-oxyl-3-oxide, PBN – N-tert-butyl-␣-phenylnitrone, PEG – polyethylene glycol, SIN-1 – 3-morpholinosydnonimine, SNP – sodium nitroprusside, SOD – superoxide dismutase, ST – sorbitol
Introduction
Water deficit is known to promote leaf senescence (Smirnoff 1993; Lin and Kao 1998; Lin et al. 1999; Lin and Kao 2000). Lipid peroxidation is considered to be an important mechanism of leaf senescence (Th-ompson et al. 1987; Strother 1988). Active oxygen species can initiate lipid peroxidation (Kellogg and Fridovich 1975). Drought-induced inhibition of pho-tosynthesis leads to increased active oxygen species in the chloroplasts (Smirnoff (1993, 1998); Dat et al. 2000). Numerous studies report increased oxidative stress and lipid peroxidation during water deficit (Sgherri and Navari-Izzo 1995; Wellburn et al. 1996;
Aziz and Larher 1998; Boo and Jung 1999; Loggini et al. 1999; Lin and Kao 2000). Thus, it is likely that drought-enhanced senescence of leaves is linked to active oxygen-mediated lipid peroxidation (Smirnoff 1993; Lin and Kao 2000).
Nitric oxide (NO) is a labile free radical and is produced from L-arginine by NO synthase in various mammalian cells (Sakuma et al. 1988; Rees et al. 1989). NO is used by mammals to regulate various biological processes of the immune, nervous and vas-cular systems (Schmidt and Walter 1994). It is now becoming evident that NO is also a ubiquitous signal in plants. NO promotes leaf expansion, seed germi-nation and de-etiolation, but it also inhibits hypocotyl
and internode elongation, induces defense genes and phytoalexin production, potentiates the induction of hypersensitive cell death, and extends post-harvest life (Leshem 2000; Beligni and Lamattina 2001). Re-cently, several reports demonstrate convincingly that NO is able to counteract the toxicity of paraquat and diaquat, which are known to generate free radicals, in potato and rice leaves (Beligni and Lamattina (1999a, 1999b); Hung et al. 2002). Thus, a possible participa-tion of NO in antioxidant system in plants, as occurs in animals (O’Donnell et al. 1997; d’Ischia et al. 2000), is suggested.
García Mata and Lamatina (2001) reported that NO is able to confer an increased tolerance to severe drought stress through induction of stomatal closure, decrease in transpiration rate and lowering of ion leakage. We have recently demonstrated that NO do-nors reduced oxidative stress of rice leave through increases in antixoidative enzyme activities and de-creases in lipid peroxidation (Hung et al. 2002). In the present investigation, we examined the effect of NO donors on water deficit-enhanced senescence of rice leaves.
Materials and methods
Rice (Oryza sativa L., cv. Taichung Native1) was cul-tured as described previously (Lin et al. 1999). The apical 3-cm segments excised from the third leaves of 12-day-old seedlings were used. Induction of wa-ter deficit was achieved by dehydration (DH), and treating with osmotica [polyethylene glycol (PEG) and sorbitol (ST)]. For leaf DH, the method of Stew-art (1971) was adopted. Groups of ten leaf segments were exposed to the vapour above a 0.5 M NaCl so-lution, by spreading the leaf segments on water-proof paper boats, which were in turn kept over 20 ml NaCl solution in Petri dishes. Comparable leaf samples were kept turgid by floating them in 20 ml of distilled water to serve as the control. For osmotically-induced water deficit, leaf segments were exposed to PEG-6000 or ST solution of osmotic potential −1.5 MPa. All NO donors and scavenger used were obtained from Sigma (St. Louis, USA) and dissolved in dis-tilled water.
RWC, defined as water content of leaf tissue as a percentage of that of the fully turgid tissue, was de-termined by the method of Weatherley (1950). Pro-line was extracted and its concentration determined following the method of Bates et al. (1973). Leaf
segments were homogenised with 3% (w/v) sulphol-salicylic acid and centrifuged. The supernatant was treated with acetic acid and acid-ninhydrin, boiled for 1 h, and extracted with toluene. Then, its absorbance at 520 nm was read. For protein extraction, leaf seg-ments were homogenised in 50 mM sodium phos-phate buffer (pH 6.8). The extracts were centrifuged at 17,000 g for 20 min, and the supernatants were used for determinations of both protein by the method of Bradford (1976) and superoxide dismutase (SOD) activity.
Malondialdehyde (MDA), routinely used as an in-dicator of lipid peroxidation, was extracted with 5% (w/v) trichloroacetic acid and determined according to Heath and Packer (1968). SOD was determined according to Paoletti et al. (1986). One unit activity for SOD was defined as the amount of enzyme which inhibited 50% the rate of NADH oxidation observed in control.
Proline, protein and MDA contents were expressed on the basis of initial fresh weight. SOD activity was expressed as units per mg protein. Absolute contents of each measurement varied among experiments be-cause of seasonal effects. However, the patterns of responses to water deficit or water deficit plus NO donors were reproducible. For biochemical analysis, four independent extractions were performed for each treatment. All experiments described here were re-peated at least three times. Similar results and identi-cal trends were obtained each time. The data reported here are from a single experiment.
Results and discussion
In the present study, leaf segments exposed to the va-pour above a 0.5 M solution of NaCl were consid-ered as DH treatment. PEG and ST solutions of os-motic potential −1.5 MPa were also used to induce water deficit in the leaves. RWC of detached rice leaves subjected to DH, and exposed to PEG or ST solution of −1.5 MPa decreased during 12-h of incu-bation (Figure 1), suggesting that DH, PEG, and ST treatments in our study did indeed cause water deficit in the leaves. The decreases in RWC in dehydrated and PEG-treated rice leaves were greater than that in ST-treated rice leaves (Figure 1). Proline is known to accumulate in leaves under water deficit (Hanson and Hitz 1982). Under our experimental condition, we also observed that proline accumulation in dehydrated and PEG-treated rice leaves was more pronounced
than that in ST-treated rice leaves (Figure 1). Based on the results of RWC and proline content we con-clude that water deficit in rice leaves induced by DH and PEG treatments was more severe than that in-duced by ST treatment. In previous study, O’Toole and Cruz (1980) reported a linear relationship be-tween leaf rolling and water potential and concluded that leaf rolling in rice could be used as an estimate of water deficit. Apparently, leaf rolling in PEG-treated leaves was greater than in ST-PEG-treated leaves (Figure 2). Since ST molecules can be absorbed by plant cells (Flores and Galston 1984), a lower degree of water deficit induced by ST appears to result from a certain amount of osmotic adjustment, due to the accumulation of ST.
Beligni and Lamattina (1999a, 1999b) reported that NO counteracts cellular damage caused by diquat and paraquat in potato leaves. Based on their evi-dence, they proposed that NO acts as an antioxidant in reducing cellular damage in potato leaves. Our re-cent work also demonstrated that the reduction of paraquat toxicity by NO in rice leaves is mediated through an increase in antioxidative enzyme activities and a decrease in lipid oxidation (Hung et al. 2002). More recently, García Mata and Lamatina (2001) re-ported that exogenous NO donors are able to with-stand water deficit, probably through reducing ion leakage, a consequence of oxidative stress. We have previously reported that the promotion of senescence (the decrease of protein content) of detached rice leaves in the light by DH was linked to lipid peroxi-dation (Lin and Kao 2000). If NO also plays a pro-tective role in water deficit-enhanced senescence of rice leaves, it is expected that exogenous NO donors would be effective in inhibiting senescence of rice leaves enhanced by water deficit.
In the present study, PBN, SIN-1, SNP, and a mix-ture of Asc and NaNO2were used as NO donors. For
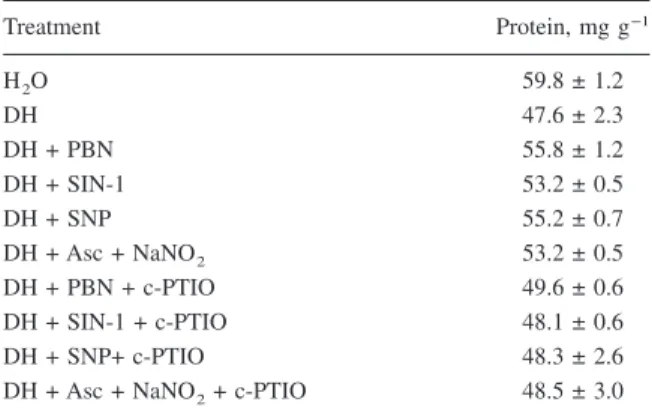
DH experiments, detached rice leaves were pretreated with distilled water or NO donors for 6 h in the light and then treated with distilled water or subjected to DH for 12 h. As indicated in Table 1, DH clearly pro-moted senescence of rice leaves and all the NO do-nors used were effective in inhibiting DH-promoted senescence in rice leaves. For PEG and ST experi-ments, NO donors were applied together with PEG or ST solution to detached rice leaves for 12 h. Both PEG and ST promoted senescence of rice leaves (Ta-bles 2 and 3). NO donors were also observed to be effective in inhibiting PEG-enhanced senescence of rice leaves (Table 2). However, NO donors had no
effect on ST-enhanced senescence of rice leaves (Ta-ble 3).
Figure 1 shows that increasing concentrations of PBN from 50 to 200 M slightly decreased protein content. When applied together with PEG, PBN con-centrations between 50 and 200M produced a clear protection against protein loss (Figure 3). The opti-mal concentration of PBN in inhibiting
PEG-en-Figure 1. Changes in RWC and proline content in rice leaves
treated with sorbitol (ST), polyethylene glycol (PEG), and dehy-dration (DH). Vertical bars represent standard errors (n = 4). Only those standard errors larger than the symbol are shown.
hanced senescence of rice leaves was observed to be 100M (Figure 3). It was also observed that the op-timal concentration of PBN pretreatment in protect-ing against protein loss induced by DH was 100M (Figure 3). The protective effect of PBN on senes-cence of rice leaves induced by DH and PEG was evident at 4 and 8 h, respectively, after treatment (Fig-ure 4).
To investigate whether the protective effect in-duced by treatment with NO donors was the result of the production of NO, 100 M c-PTIO, a NO-spe-cific scavenger, was applied along with NO donors.
The protective effect of NO donors on PEG- and DH-induced senescence of rice leaves could be reversed by c-PTIO (Tables 1 and 2). Clearly, the protective effect on PEG- and DH-induced rice leaf senescence by NO donors is due to NO.
Table 4 shows that PEG or DH treatment resulted in a marked increase in MDA content, but ST treat-ment had no effect on MDA content. Treattreat-ment with PBN caused a reduction of the PEG- and DH-induced increase in MDA content (Table 4). The effect of PBN on PEG- and DH-induced MDA content could also be reversed by c-PTIO (Table 4). MDA is routinely used as an indicator of lipid peroxidation. The protective
Figure 2. Effect of sorbitol (ST) and polyethylene glycol (PEG) on leaf rolling of rice. Picture was taken after 8 h of treatment. Table 1. Effect of NO donors on protein content in dehydrated rice
leaves in the presence or absence of c-PTIO
Treatment Protein, mg g−1 H2O 59.8 ± 1.2 DH 47.6 ± 2.3 DH + PBN 55.8 ± 1.2 DH + SIN-1 53.2 ± 0.5 DH + SNP 55.2 ± 0.7 DH + Asc + NaNO2 53.2 ± 0.5 DH + PBN + c-PTIO 49.6 ± 0.6 DH + SIN-1 + c-PTIO 48.1 ± 0.6 DH + SNP+ c-PTIO 48.3 ± 2.6
DH + Asc + NaNO2+ c-PTIO 48.5 ± 3.0 Detached rice leaves were pretreated with distilled water or NO donors in the presence or absence of c-PTIO for 6 h in the light and then treated with distilled water and subjected to dehydration (DH) for 12 h. The concentrations of PBN, SIN-1, SNP, Asc, NaNO2, and c-PTIO were 100, 100, 100, 100, 200, and 100M, respectively. Data are the means ± standard errors (n = 4).
Table 2. Effect of NO donors on protein content in PEG-treated
rice leaves in the presence or absence of c-PTIO
Treatment Protein, mg g−1 H2O 56.8 ± 0.5 PEG 47.2 ± 1.7 PEG + PBN 51.1 ± 1.6 PEG + SIN-1 51.7 ± 1.4 PEG + SNP 52.0 ± 1.6
PEG + Asc + NaNO2 51.9 ± 1.5
PEG + PBN + c-PTIO 46.4 ± 0.7
PEG + SIN-1 + c-PTIO 46.2 ± 1.7
PEG + SNP+ c-PTIO 46.0 ± 0.4
PEG + Asc + NaNO2+ c-PTIO 47.7 ± 1.0 The concentrations of PBN, SIN-1, SNP, Asc, NaNO2, and c-PTIO were 100, 100, 100, 100, 200, and 100M, respectively. Protein content was determined 12 h after treatment in the light, data are the means ± standard errors (n = 4).
effect of PBN on senescence of rice leaves induced by PEG and DH is clearly associated with the de-crease in lipid peroxidation (Table 4). Plant cells are equipped with several free radical detoxifying en-zymes to protect them against oxidative damage. These enzymes include SOD, ascorbate peroxidase, glutathione reductase, catalase, and peroxidase (Foyer et al. 1997; Nakano and Asada 1981). The striking changes in lipid peroxidation seen in leaves treated with PEG or DH alone may be a reflection of the in-ability of the scavenging enzymes to cope with the increased levels of oxidant species and/or their
deg-radation by these species. Previously, we found that DH-induced enhancement of rice leaf senescence co-incided with an elevated lipid peroxidation and re-duced SOD activity in detached rice leaves (Lin and Kao 2000). In the present study, the PEG- and DH-induced loss of SOD activity was found to be pro-tected by PBN (Table 4). Table 4 also showed that c-PTIO arrested PBN-mediated SOD protection (Ta-ble 4). It should be noted that ST treatment had no effect on SOD activity and MDA content (Table 4).
Oxidative stress or lipid peroxidation was gener-ally considered to be generated as a consequence of water deficit in plants. Baisak et al. (1994) reported that enhancement of lipid peroxidation was observed in leaves subjected to a higher degree of water deficit but not in those subjected to mild stress. Parallel to these observations, we also noticed an increase in lipid peroxidation in detached rice leaves treated with PEG and DH. On the other hand, detached rice leaves did not exhibit an increase in lipid peroxidation when exposed to ST (Table 4). Our results show that the promotion of rice leaf senescence caused by PEG and DH, which induced lipid peroxidation, can be coun-teracted by NO donors, but the senescence caused by ST, which had no effect on lipid peroxidation, cannot be counteracted by NO donors.
Table 3. Effect of NO donors on protein content in sorbitol
(ST)-treated rice leaves in the presence or absence of c-PTIO
Treatment Protein, mg g−1 H2O 55.2 ± 0.6 ST 49.8 ± 0.9 ST + PBN 50.0 ± 1.6 ST + SIN-1 49.4 ± 3.3 ST + SNP 47.1 ± 1.7 ST + Asc + NaNO2 48.8 ± 1.4
The concentrations of PBN, SIN-1, SNP, Asc, and NaNO2were 100, 100, 100, 100, and 200M, respectively. Protein content was determined 12 h after treatment in the light, data are the means ± standard errors (n = 4).
Figure 3. Effect of PBN concentrations on protein content in rice leaves treated with either distilled water, dehydration (DH) or polyethylene
glycol (PEG). For the PEG experiment, protein was determined 12 h after treatment in the light. For the DH experiment, detached rice leaves were pretreated with distilled water or various concentrations of PBN for 6 h in the light and then were treated with distilled water or sub-jected to dehydration for 12 h. Vertical bars represent standard errors (n = 4). Only those standard errors larger than the symbol are shown.
The present study extends our previous results concerning the putative physiological roles of NO in rice leaves (Hung et al. 2002). We now demonstrate the potential ability of NO to withstand water deficit. The inhibition of PEG- and DH-enhanced senescence of rice leaves by NO is most likely mediated through
an increase in SOD activity and a decrease in lipid peroxidation.
Figure 4. Changes in protein content in rice leaves treated with distilled water, polyethylene glycol (PEG) or dehydration (DH) in the
pres-ence or abspres-ence of PBN in the light. PBN concentration was 100M. Vertical bars represent standard errors (n = 4). Only those standard errors larger than the symbol are shown
Table 4. Effect of PBN on MDA content and SOD activity in PEG-, ST-treated and dehydrated rice leaves in the presence or absence of
c-PTIO
Treatment MDA, nmol g−1 SOD, units mg−1protein
H2O 40.3 ± 0.9 1.70 ± 0.03 ST 43.4 ± 2.1 1.51 ± 0.09 PEG 67.3 ± 1.6 1.04 ± 0.14 PEG + PBN 53.1 ± 1.4 1.59 ± 0.10 PEG + PBN + c-PTIO 68.2 ± 2.1 0.99 ± 0.14 H2O 40.3 ± 0.4 1.70 ± 0.04 DH 93.4 ± 1.2 0.73 ± 0.02 DH + PBN 76.5 ± 2.3 1.17 ± 0.03 DH + PBN + c-PTIO 91.1 ± 1.5 0.71 ± 0.01
The concentrations of PBN and c-PTIO were 100 and 100M, respectively. For sorbitol (ST) and polyethylene glycol (PEG) experiments, MDA and SOD were determined at 12 h after treatment in the light. For dehydration (DH) experiment, detached rice leaves were pretreated with distilled water or PBN in the presence or absence of c-PTIO for 6 h in the light and then treated with distilled water or subjected to dehydration for 12 h. Data are the means ± standard errors (n = 4).
Acknowledgements
This work was supported by grant NSC 90-2313-B-002-267 from the National Science Council of the Republic of China.
References
Aziz A. and Larher F. 1998. Osmotic stress induced changes in lipid composition and peroxidation in leaf discs of Brassica
nupus. L. J. Plant Physiol. 153: 754–762.
Baisak R., Rana D., Acharya P.B.B. and Kar M. 1994. Alterations in the activities of active oxygen scavenging enzymes of wheat leaves subjected to water stress. Plant Cell Physiol. 35: 489– 495.
Bates L.S., Waldrer R.P. and Teare I.D. 1973. Rapid determination of free proline for water stress studies. Plant Soil 39: 205–207. Beligni M.V. and Lamattina L. 1999a. Nitric oxide counteracts cy-totoxic processes mediated by reactive oxygen species in plant tissues. Planta 208: 337–344.
Beligni M.V. and Lamattina L. 1999b. Nitric oxide protects against cellular damage produced by methylviologen herbicides in po-tato plants. Nitric Oxide Biol. Chem. 3: 199–208.
Beligni M.V. and Lamattina L. 2001. Nitric oxide in plants: the history is just beginning. Plant Cell Environ. 24: 267–278. Boo Y.C. and Jung J. 1999. Water deficit-induced oxidation stress
and antioxidative defenses in rice plants. J. Plant Physiol. 155: 255–261.
Bradford M.M. 1976. A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the princi-ple of protein-dye binding. Anal. Biochem. 72: 248–254. Dat J., Vandenabeele S., Vranová E., Van Montagu M., Inzé D. and
Van Breusegem F. 2000. Dual action of the active oxygen spe-cies during plant stress responses. Cell. Mol. Life Sci. 57: 779– 795.
d’Ischia M., Palumbo A. and Buzzo F. 2000. Interaction of nitric oxide with lipid peroxidation products under aerobic condi-tions: Inhibitory effects on the formation of malondialdehyde and related thiobarbituric acid-reactive substances. Nitric Ox-ide Biol. Chem. 4: 4–14.
Flores H.E. and Galston A.W. 1984. Osmotic stress-induced polyamine accumulation in cereal leaves I. Physiological pa-rameters of the response. Plant Physiol. 75: 102–109. Foyer C.H., Lopez-Delgado H., Dat J.F. and Scott I.M. 1997.
Hy-drogen peroxide and glutathione associated mechanism of ac-climatory stress tolerance and signaling. Physiol. Plant. 100: 241–254.
García Mata C. and Lamatina L. 2001. Nitric oxide induces sto-matal closure and enhances the adaptive plant responses against drought stress. Plant Physiol. 126: 1196–1204.
Hanson A.D. and Hitz W.D. 1982. Metabolic responses of meso-phytes to plant water deficits. Annu. Rev. Plant Physiol. 33: 163–203.
Heath R.L. and Packer L. 1968. Photoperoxidation in isolated chl-roplasts I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125: 189–198.
Hung K.T., Chang C.J. and Kao C.H. 2002. Paraquat toxicity is reduced by nitric oxide in rice leaves. J. Plant Physiol. 159: 159–166.
Kellogg E.W. and Fridovich I. 1975. Superoxide, hydrogen, perox-ide, and singlet oxygen in lipid peroxidation by xanthine oxi-dase system. J. Biol. Chem. 250: 8812–8817.
Leshem Y.Y. 2000. Nitric Oxide in Plants. Function, Occurrence and Use. Kluwer Academic Publishers, Dordrecht.
Lin J.-N. and Kao C.H. 1998. Water, stress, ammonium, and leaf senescence in detached rice leaves. Plant Growth Regul. 25: 165–169.
Lin J.-N. and Kao C.H. 2000. Involvement of lipid peroxidation in water stress-promoted senescence of detached rice leaves. Biol. Plant. 43: 141–145.
Lin J.-N., Wang J.-W. and Kao C.H. 1999. Effect of abscisic acid and water stress-promoted senescence of detached rice leaves. Biol. Plant. 42: 313–316.
Loggini B., Scartazza A., Brugnoli E. and Navari-Izzo F. 1999. Antioxidative defense system, pigment composition, and pho-tosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiol. 119: 1091–1099.
Nakano Y. and Asada K. 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 22: 867–880.
O’Donnell V.B., Chumley P.H., Hogg N., Bloodsworth A., Darley-Usmar V.M. and Freeman B.A. 1997. Nitric oxide inhibition of lipid peroxidation: Kinetics of reaction with lipid peroxyl radi-cals and comparison with tocopherol. Biochemistry 36: 15216– 15223.
O’Toole J.C. and Cruz R.T. 1980. Response of leaf water poten-tial, stomatal resistance, and leaf rolling to water stress. Plant Physiol. 65: 428–432.
Paoletti F., Aldinueci D., Mocali A. and Capparini A. 1986. A sen-sitive spectro- photometric method for the determination of su-peroxide dismutase activity in tissue extracts. Anal. Biochem. 154: 536–541.
Rees D.D., Palmer R.M., Hodson H.F. and Moncada S. 1989. A specific inhibitor of nitric oxide formation from L-arginine at-tenuates endothelium-dependent relaxation. Br. J. Pharmacol. 96: 418–424.
Sakuma I., Stuehr D.J., Gross S.S., Nathan C. and Levi R. 1988. Identification of arginine as a precursor of endothelinm-derived relaxing factor. Proc. Natl. Acad. Sci. USA 85: 8664–8667. Schmidt H.H.W.H. and Walter U. 1994. NO at work. Cell 78: 919–
925.
Sgherri C.L.M. and Navari-Izzo F. 1995. Sunflower seedlings sub-jected to increasing water deficit stress: oxidative stress and defence mechanisms. Physiol. Plant. 93: 25–30.
Smirnoff N. 1993. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 125: 27– 58.
Smirnoff N. 1998. Plant resistance to environmental stress. Curr. Opin. Biotechnol. 9: 214–219.
Stewart C.R. 1971. Effect of wilting on carbohydrates during in-cubation of excised bean leaves in the dark. Plant Physiol. 48: 792–794.
Strother S. 1988. The role of free radicals in leaf senescence. Ger-ontology 34: 151–156.
Thompson J.E., Legge R.L. and Barber R.F. 1987. The role of free radicals in senescence and wounding. New Phytol. 105: 317– 344.
Weatherley P.Z. 1950. Studies in the water relation of cotton plant. I. The field measurement of water deficits in leaves. New Phy-tol. 49: 81–97.
Wellburn F.A.M., Lau K.-K., Milling P.M.K. and Wellburn A.R. 1996. Drought and air pollution affect nitrogen cycling and free
radical scavenging in Pinus halepensis (mill). J. Exp. Bot. 47: 1361–1367.