11, 4,
MaryAnnLiebert, Inc., Publishers
Pp.315-320
Structure and
Expression
of
Mouse
cxi-Acid
Glycoprotein
Gene-3
(AGP-3)
CHING-JIN
CHANG,
MING-YANGLAI,
DING-SHINNCHEN,
and SHENG-CHUNG LEEABSTRACT
The genome ofMus
domesticus
hasmultiple
genes of thea,-acid
glycoprotein
(AGP).
TwocDNA cloneswereidentified
corresponding
to AGP-1 and AGP-2.Moreover,
two alíelesof AGP-1 exist in inbred mice.The
genomic
DNA ofthe AGP-2gene has been clonedand
studied. Herewereport
thegenomic
organiza-tion ofthree M.
domesticus AGP
genes, the sequenceanalysis
oftheAGP-3genomic
DNA,
andthe expres-sion of the AGP-3 gene. Themajor
structural differences between AGP-2 and AGP-3 genesare located in introns 1 and 5. The low level ofAGP-3
mRNA can be detectedby
thepolymerase
chain reaction(PCR).
The molecular basis of the low level
expression
ofAGP-3
and thepossible
classification of AGP-3 as apseudogene
arediscussed.INTRODUCTION
Cci'AciD
glycoprotein(AGP)
is asingle-chain
poly-peptide
withamolecularweight
ofapproximately
44,000,
containing
about45%carbohydrate.
Thisplasma
protein
is anacute-phase
reactantsynthesized
in the liver ofMus domesticus. Plasma concentrations of AGP in the mouseincrease several fold
during infection, inflammation,
or areinduced
following
subcutaneousinjection
ofturpentine
orlipopolysaccharide. Expression
of the AGP gene is regu-latedby
interleukin-1(IL-1),
tumor necrosis factor-a(TNF-a),
interleukin-6(IL-6),
andglucocorticoids
(Bau-mann and
Maquat,
1986;
Klein etai, 1987;
Prowse andBaumann,
1988;
WonandBaumann, 1990).
Thebiological
function of AGP isunknown,
but there are indicationsthat it may suppress the immune response
(Bennett
andSchmid, 1980).
Someexperiments
suggest that AGP is anonspecific
antiinfection agent(Friedmann, 1983),
andthat it possesses nerve
growth-promoting activity
(Liu
etai, 1988).
Multiple
forms ofmouse AGP canbedemonstratedby
two-dimensional
gel
electrophoresis
(Baumann
etai,
1984),
andahigh degree
ofheterogeneity
intheaminoacidcomposition
of humanAGP has been described(Schmid,
1975).
Denteet al.(1987)
cloned andsequenced genomic
DNA segments
coding
for three humanAGPgenes. Theone
designated
AGP-A,
is transcribed to agreater extentthan AGP-B orAGP-C
(Dente
etai,
1987). Previously,
we
reported
the existence of at least two AGP genes, AGP-1 andAGP-2,
in M. domesticus(Lee
etai,
1989).
Both c/s-elements and
rra/is-acting
factorsreportedly
in-volved in the
regulation
of AGPexpression
havebeenana-lyzed
extensively
(Lee
etai,
unpublished observations).
One of
these,
AGP/EBP(a
liver-enrichedtranscription
factor),
has beenanalyzed
and iskey
toregulating
AGPexpression (Chang
etai, 1990).
Tostudy
thegenomic
orga-nization,
the molecular basis ofthedifferentialexpression,
and the
regulation
ofAGPexpression,
weanalyzed
theM. domesticus AGP genes. We nowshow thattherearethreeAGP genes in theM. domesticus genome; we determined
the
complete
sequence ofthenewly
identifiedgene,AGP-3,
andshowthat AGP-3mRNA isexpressed
atverylowlevels.MATERIALS
AND METHODS Isolationof
AGPgenomic
clones
A
genomic library
fromTlymphocytes
ofC57BL/6 M. domesticus andE. coliK803(r",
m",gal",
met")
weregifts
of Dr. L. Mori
(University
ofMilan,
Italy).
Thelibrary
was screened with an AGP-1 cDNA
probe (Lee
etai,
1989).
Of theapproximately
4 x 105plaques
screened,
four
positive plaques
were recovered andpurified by
sec-ondary screening. Sequencing
wascarriedoutby
thedide-oxynucleotide
termination method(Sanger
etai, 1977).
Institute ofBiological Chemistry, AcademiaSinicaand Institute of ClinicalMedicine,National TaiwanUniversity, Taipei,Taiwan.
316 CHANG ET AL. RNase
protection
assayThe RNase
protection
assaywasperformed
as describedby
Meltonetai(1984).
Total RNA(20 ^g)
was dissolved in 20fd
ofhybridization
buffer(60% formamide,
40 mMPIPES
pH 6.7,
400 mMNaCl,
1 mMEDTA)
containing
theriboprobe
(2-5
x 105cpm).
Thehybridization
mixturewasheated at 85°C for 5
min,
followedby
incubation at45°C foraminimumof 4 hr. Then300
/tl
of RNasediges-tion buffer
(10
mMTris«HClpH
7.5, 5mMEDTA,
300 mMNaCl) containing
40fig/ml
ofRNase Aand50 U/mlof RNaseTl. The reaction wascontinued at 30°C for 30
min,
thenstopped by
additionof20fd
of10%NaDodS04
and50figof
proteinase
K. Afterincubating
at37°C for15min,
the mixture was extracted withphenol/chloroform
andprecipitated
withethanol(with
20 fig carrier tRNAin-cluded).
The sequences oftheRNAsamples
werethende-termined and
analyzed.
Polymerase
chain reactionEach
polymerase
chainreaction(PCR)
included10 ng of cloned DNAfragments
or 100 ng ofsingle-stranded
cDNA,
1.25 units of ofTaq polymerase,
100fiM
deoxy-ribonucleotides,
2.5 nMeacholigonucleotide primer,
1.5 mMMgCl2,
50 mMKC1,
and 10 mMTris-HClpH
8.3 inafinal volume of 100
¡d.
The reaction mixture was overlaidwith mineral
oil,
andrunthrough
30cycles
of 1 mineachat
94°C,
1 minat57°C,
and30sec at72°C. Theproduct
was
analyzed
by
native20%polyacrylamide gel.
proximately
16kb,
while theXAgp-25
plaques
containedan insert of
approximately
18 kb. Three distinct AGPgeneswereidentified
by
restrictionmapping
andpartial
se-quencing
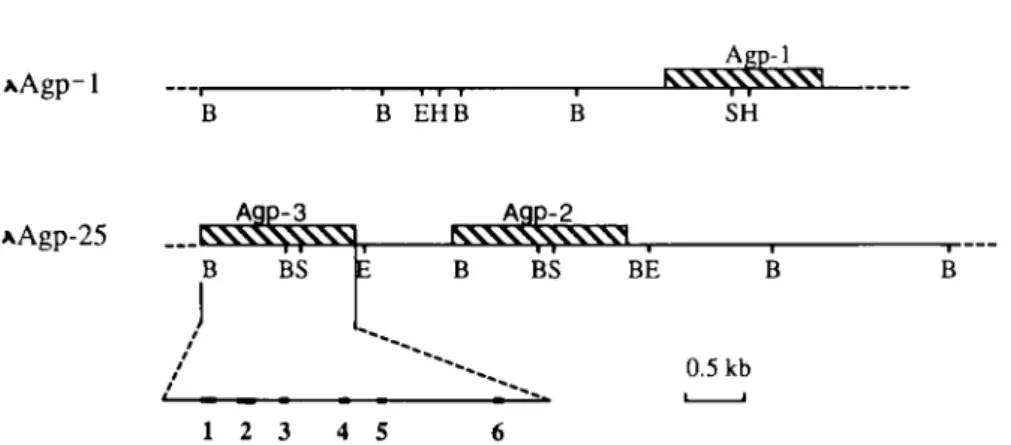
ofthese five clones(Fig.
1).
cDNAscorrespond-ing
to twoofthem,
AGP-1 andAGP-2,
hadbeencharac-terized
previously
(Cooper
andPapaconstantinou,
1986;
Leeet
ai,
1989),
andathirdgenedesignated
AGP-3wasidentified. AGP-1 is contained in X clone
Agp-1
whileAGP-2and AGP-3 are in
XAgp-25
clones.Only
two types of cDNAs and thecorresponding
pro-teins of AGP have been identified in the M. domesticus
(Baumann, 1984;
Leeetai,
1989).
To address the issue of whether or not AGP-3 isexpressed,
thegenomic
DNAcontaining
thecoding
andnoncoding
sequencesofAGP-3was
sequenced.
This gene spans 4 kb from 548bp
up-streamfrom the
putative transcription
initiation siteto 130bp
downstream ofthepolyadenylation signal,
AATAAA(Fig.
2).
This gene has 6 exonsand 5introns,
a structureanalogous
tothe rat(Liao
etai, 1985;
ReinkeandFeigel-son,
1985),
human(Dente
etai,
1985; Dente etai, 1987),
and mouse AGP-2 genes
(Cooper
etai, 1987;
thePapa-constantinougroup
designates
itAGP-1,
while our desig-nation isAGP-2).
Theconsensus sequence forsplice
junc-tions is
preserved
in both AGP-2 and AGP-3 genes.The mouse AGP-3 and AGP-2 gene
(Cooper
etai,
1987)
sequences are verysimilar; however,
there are twomajor
differences between them: AGP-3 lacks 86nucleo-tidesin intron 1
(between
+331 and+332),
whileAGP-2has an additional
(GT)28
tract in intron 5(+2815),
notfound in AGP-3.
RESULTS
Isolation and
characterization
of
M.domesticus
AGP
genomic
clonesSeveral
genomic
clones were obtainedby
screening
theEMBL-3
library (C57BL/6)
with an AGP cDNAprobe.
Fourpositive plaques
ofXAgp-1
containedaninsertofap-Differential
expression of
AGPgenesIt would be
interesting
to know the relative level of mRNAexpression
amongAGP-1, -2,
and -3 genes in M domesticus liver under normal conditions andduring
theacute-phase
reaction. Becauseofthehigh
degree
ofsimi-larity
amongthesethreegenes, anRNaseprotection
analy-sis was
performed
to discriminate among these mRNAs..lkb.
xAgp-1
B EHBkX-xV^v\
X^ SHAAgp-25
^\S\S\Vsl
35
BS A9P-2^5
NNNXNXNXWI B BS BE 0.5kb 12 3 4 5 6FIG. 1. Partial restrictionmap of«,-acid
glycoprotein
genomic clones,
XAgpl
andXAgp25.
Shaded boxes representAGP-1,
-2,and -3 genes.Exonsarerepresented
by
solid boxes. Abbreviations: B,BamHI; E,
EcoRI;H,
Hind III;S,
Sail.-548 GGATCCTTTCCTGCTGTAGATACTGGGAGCTTTGCTGAACTAGATGTTCAAGTCA -4 93 GAATCAACCCTTTTTGGGCATTTGGATGCCTCTAGGCTGGGAAGGGGTCCTCAGG -438 AACATCACACTCCTTTGGAAACTAATCCATCTTTGTCCTTGGGCCTTAACTTGAG -383 CCCCTAAGTGTCTTCTATGTTCACTATGAACTTGACCTGGGACCCCTTCTTATCA -328 TGCTTGGGGGCGGGTTGATGTATGTGTAGGTTTCACTCCTGCTAGGCAGCTTCAT -273 GGGATAAGAGAGGGTGGGGACCACTGTCTGGGACCTAAGTATCATCAGGCTACCC -218 TGTACCCACCTTGACCATGAATCAGCCACTCTGGTGTAGGGCAGGAGCCTGTGTC -163 ACAGCCAGCTGGCTGAGAGAGCTGCACAAAGCTGGCTTGAGGGAACATTTTGCAC -108 AAGACATTTATCAAGTGCTGGTGAGTTTGTGGCACTGCTCTACAGCCCCTGGCTG +1 -53
CAGTCCCATGCCCTCCCCACATCCTGTtTATAÄJAAGCCACTGCACCCTCCAGCCAC
Exon 1 3 CAGTTATCTCTTCCAAGCTCTGGTGCCTCTGAGTATCCTCAGCATGGAACTACAC MetGluLeuHis 58 ACGGTTCTCATCATGTTGAGCCTCCTGCCACTGTTGGAAGCTCAGAACCCAGAGC ThrValLeuIleMetLeuSerLeuLeuProLeuLeuGluAlaGlnAsnProGluH 113 ATGCCATCAACATAGGCGACCCTATCACCAATGAGACCCTGAGCTGGGTAAGTGT isAlalleAsnlleGlyAspProIleThrAsnGluThrLeuSerTrp 168 CTGCCCGGGGCCTGGACCTGTTCACTGTAGGATTCTACTCTTTCCTCTGGGCTTT 2 2 3 CCCTTCCCTGGTGTCTGTGTTCAGCTCTGGGCTCCTGGTACTGCCCTTCCCACTT 2 78 GTGTATACCCTGGTGGCATCCCCCTGTCTCCAAAGGCAGAATCATCACTCTGAGC 333 CATAGCTTGCCTGCCCCTCATCGTGGATGAATGCCAAGGTCCTCACTACAAGGCC Exon 2 388 TGCTCATCTGTGTGCCTGCTTCTCCCCAGCTATCTGGCAAATGGTTTCTCATTGC LeuLeuGlyLysTrpPheLeuIleAl 44 3 TGTGGCTGACTCAGACCCTGATTATAGGCAGGAAATTCAAAAGGTACAGACTATA aValAlaAspSerAspProAspTyrArgGlnGluIleGlnLysValGlnThrlle 4 98 TTTTTTTACCTTACCCTAAACAAGATAAATGACACGATGGAGCTTCGAGAGTATC PhePheTyrLeuThrLeuAsnLeulleAsnAspThrMetGluLeuArgGluTyrH 553 ACACCAAGTGAGTCCTTGTAACAGCCAGCCCACCCTGGCCCTGGCTTCCACTCCC isThrLy 608 AGATTCCTAGAGACCTGAGCAAACTGGCTCTGCCTGGCCTCCCCACCCACTTTCA 663 GAAATGGGGACAGCTGTCTTGCCTCTTGCCCCCTTCTACCCTGGGCTAGTCAGAT Exon3 718 CACCTCTCCATCAGTTGTCCTCTCTCTTTGCTTTTTAGAGATGACCACTGTGTCT sAspAspHisCysValT 77 3 ATAACTCCAACCTTCTGGGATTCCAGAGAGAGAATGGGACCCTCTTCAAGTATGG yrAsnSerAsnLruLeuGlyPheLeuArgGluAsnGlyThrLeuPheLysTyrG 82 8 TGAAGGTGTGAACTCACTCCTTTCTGGGAGGGTATTGCCAGTTCTGAGGGGACAG 88 3 CAGAACAGGGCAGTTTGGTCTGTCAAGTCACTCTCTGGGGCTTGTAAGTGGACGT 93 8 AGATTTCAAACTGGAGTCCAGCTGCCAGGCCTGACTTGTTTTGATCAGGCTTTAT 993 GACCCTTCGTGACCTCAGAGTGGAAAGCCATGGGTTGGGAAACCAGTGACTTTAG 104 8 CCACACCCCCAGGTCACCAGGGACAGCATGGAGGAGAACAACCCAATTGCTGGTG 110 3 GGCCAGATCAGACACTGGTTAGCTTTTAATTGTCCTAAGCAGATGTTTTGATGTT 1158 TAAGGGGAAGTGTTAACATTATTATTGCCCCACCACCACACCACCAAGGGCCCTT 1213 TCCAAGGCCCCAGCTCTTCCATACCTATGAAGAATGAGAAGTGAGGCTTGCATCC 12 68 AGCATGAGGCTGAGCACATGGCAGCCTCAGGGAGCCCCAGGCACTTGCCATAGCT 1323 ATGTGCTTCCTTCCCTTGGGGATGTGAACCACATCATCATTCTAGTGACTCACAA Exon* 1378 AGACCTTCCTCCAACAGAAGGAGAAGTAGAAAACCCTTCTCACCTGAGAGTGCTA luGlyGluValGluAsnProSerHisLeuArgValLeu 14 3 3 GAGAAACATGGGGCCATCATGCTTTTCTTTGACCTGAAGGATGAGAAGAAACGGG ArgLysHisGlyAlalleMetLeuPhePheAspLeuLysAspGluLysLysArgG 1488 GACTGTCCCTGAGCGGTAGGGTCCTCTCATCCCTGGTGCGCCCAGCTCAACTGGC lyLeuSerLeuSerA 154 3 CTTACTCTTGGTCACCCACCCACATCCCACATCCCTACCTGGCCTCCCATCTACT Exon 5 15 98 GGACCCATAGCCAGCATAACTTTGGATCCCTTCTCCCATGCAGCTAGAAGGCCAG laArgArgProA 1653 ATATCCCCCCGGAGCTGCGGGAAGTATTTCAGAAGGCTGTCACACACGTGGGCAT spIleProProGluLeuArgGluValPheGlnLysAlaValThrHisValGlyMe 17 08 GGATGAATCAGAAATCATATTTGTCGACTGGAAAAAGGTAAATGAAGGAGGCTGT tAspGluSerGluIlellePheValAspTrpLysLys 17 63 ACGATACCACCCCAGCAGTGCGCCCAGTGTCAGTGACCCTAGAGGCTCAGAGAGG 1818 GCAAGCTTCTGGTTAAGGCAGCTCAGCGAGGCAGGTATCTTGTTAACTCTCCTGC 187 3 CTCCTCCTCATCAGGAGATCACAGAGACCCTAGATGGGCAGTGAGCCTCAGGGAG 192 8 GTGAAGTTAAGTAGGAGGTCCTGGAAAGCTTGTGGAGGATAAGAGGAAGATCAGG 198 3 AGGGTCACTTAGGGAACAGCCAGTGCCAGGGTGCCAGGTTTCTTCCTGTCCTTCA 2 0 3 8 TATTACTACCTTTTCAAGCAGGAGTTTTGATTGACATCTTCCATGTCACCCCAAC 2 0 93 TCCAGCAAGCCCGATGGCTTTGATAGGCAGGGTTGACCACACTGAGACTCTTGAT 214 8 GTCCGGTCTACACATTGTGCAGAGGGAGAGGCAGCATCAGTTTTGTTTTTCACCC 22 03 ATGGCGAATGCATGGGATCAAACAGTCACCTTGCATGTAGTTTAAGATACTCAAT 22 58 AGCTTTTGTAACTTCATTCTCTGGTCACCTGAGCCTTTCCTGGCATCATCACCAG 2313 CCCCAGGATTCCCGGGAGAGGTGCCTGCACACAGACACTGCCATTCACAGCATGA 2 368 CTTCCACCCACACCAGTGGGCCAGTAGACTCATCCTGCACCTGTGGACAGAAGTG 2423 TTAGATAATGCCTGCCCTTTGGGGATTCTGCTCACAATCAATGGGTGAATAAGCC 2 478 GGAGCTCAGAGATGAGGGACAACTTACCCTAGACTAGTGGTTCTCTAGGATGGGT 2 5 33 CTCAACCTGTATGTCAGATGTCCTGCACGTCAGGTATTTATTGATTTATAATAGT 2588 AGCATATAATTACAGTTATTGAAGTAGCAATGAAATCATGGTAGGTGATCACCAC 2 64 3 AACACAAGAAACTGTGTTAAAGTTTTGCATTATTAGGAAGGCTGAGAACCACTGT 2 6 98 CCTGCCACTGCAGGGAGCCATGGCAGATCTAAGACACATCTGGTTGACACTACCG 2753 GGCCATTTTGACCAACAACAGTACTCCCCCCAACCCACCTCACAATAGGTGTATT 2808 CATAGCTAG_TGTG_T_G;rGC_A_TGT_GTGA_C_TGTG_TGGGTACACAAGCATGCTATAAGA 2 863 CATGTGTGGATGTCAGAGGACACCTGTGGGCTATGTCCTCTTCTACCATTCTCTC 2918 CTGGGCTCTGGTTAAGGCTGGGTTGGCTTCAAGCTGCCCCTCAGGCTTACCTACC 2 973 TTGCCATTTTTTTTTGTTGTTCTGTCCTGTTTTTTCTGTTTTGTTTTGTTTTTGT 3028 ATTTAATCTTGCAGCCCAGGCTACTCTACTGCAACTCATAGCAATCCTCTTGCCT 30 83 CAGTATTCATCAACCCTGGTGTGTGCCACCAGCCCTGGCTTACTCACTCTGCTCT Exon6 3138 CCTCCCTGATATCTTCCAGGACAGGTGCAGTGAACAGGAAAAGAAGCATCTTGAG AspArgCysSerGluGlnGluLysLysHisLeuGlu 3193 TTGGAGAAGGAGACCAAGAAAGATCCTGAGGAAAGCCAGGCATGAACTCAGCTCT LeuGluLysGluThrLysLysAspProGluGluSerGlnAla 3243 CTGGTCTCCTTGGGCTGTCCCCATGTGTACCACACCCTACCCCATCCTGGTCACT 3303 TTGATTCTGTCTCTGTAAdAATAÄÄfeGTTTGCTGACACTGTCAATATCATTTCTT 3358 TGCTCCCTTCCTTTTCCTCCCTCCCTCCCTCCCTTCGTGGAGAGTCTTGAGTGGA 3413 GCTAGCTAAGTCAATAACCCTGCCAGGAATTCGAAAGGCTCTFIG. 2. Nucleotide sequence ofmouse AGP3 gene. The
putative
TATA box andpoly(A)
additionsignal
sequence(AATAAA)
areboxed. Sixexonsandencoded amino acidsequencesareshown. The siteof initiationof
transcription
is
depicted by
+1.The
specific riboprobes
of AGP-1 and AGP-2(Fig.
3a,
upper
panel)
wasdesigned
from their cDNA sequences(Lee
etai,
1989).
Theriboprobe
of AGP-3 was derivedfrom the
genomic
segments that contains theputative
exon4
(Fig.
3a).
The result of RNaseprotection analysis
is showninFig.
3a. Whenusing
RNAfrom normalliver andfrom
lipopolysaccharide-stimulated
liver,
protected
bandsweredetected
corresponding
to 247bp
ofAGP-1 and 318bp
ofAGP-2(Fig.
3a, lowerpanel,
lanes 1, 2orN-LandLpS-L).
However,
there were nosignals
for theprobe
de-rived from AGP-3. Theexpression
level of AGP-1 isabout fivefoldhigher
than that of AGP-2 and both genesre-spond
totheacute-phase reaction,
thusAGP appearsas aliver-specific
gene, because nosignal
wasdetectedin RNAfrom the
spleen.
Because the level of
expression
of AGP-3 is much less than AGP-1 andAGP-2,
a more sensitive method(e.g.,
PCR)
wasusedtodetectit. PrimersforAGP-2 andAGP-3were
synthesized
for PCRexperiments (Fig.
3b,
upperpanel).
Using
genomic
DNAofAGP-2orAGP-3genesastemplates,
we showed that theprimers
derived fromAGP-2 and AGP-3 were
specific
for thecorresponding
templates
(Fig.
3b,
lowerpanel,
lanes1, 2, 4,
5). However,
when theseprimers
were used for reversetranscriptase
(RT)-PCR
using
RNAderived from M. domesticusliver,
an
AGP-2,
but not anAGP-3fragment
canbeseeninthe agarosegel by
ethidium bromide. This does not excludethe
possibility
thatthelevel ofexpression
ofAGP-3might
318 CHANG ET AL. riboprobe protectedlength RI Anp-2 437
|
^-(390ni) (318ni) riboprobe protected length^g
WiW (570ol) riboprobe(108) protectedlength Agp2 primer R: 5'CCATGACAAGAATCATGTGC3'
+67 +52 N-L LPSL LPSS Probe 123123123123 L: 5'ATCTCTTCCAAGCCCTG3' +8 +24
Agp3 primer6F H R: 5' GAACCGTGTGTAGTT 3'
+64 +50
L: 5'ATCTCTTCCCAGCTCT 3'
+8 +24
1 2 3 4 5 6 7 8
FIG.3. a.
Upper panel.
RNaseprotection
assay.Specific riboprobes
andtheirprotected length
forAGP-1, -2,
and-3 arerepresented.
Theriboprobes
of AGP-1 and -2are derived from their cDNAs inpGEM3
vector.Thefragments
arenumberedrelative to the mRNA
transcription
initiation site indicatedby
+1. Theriboprobe
of AGP3 containsexon4andintronsegmentsin
pGEM
4vector. Lowerpanel.
RNaseprotection experiment
onRNAprepared
frommouseliver andspleen.
1, 2,
and 3 indicate theriboprobes
derived fromAGP1, 2,
and 3.N-L,
LPS-L, and LPS-S indicate thesources of RNA from normal
liver,
LPS-stimulatedliver,
and LPS-stimulatedspleen, respectively. Specific protection
bands are
represented by
arrows, b. PCRamplification
of AGP-2 andAGP-3. Theright
(R)
and left(L) primers
ofAGP-2 and AGP-3 are shown above, and the PCR
products analyzed
on 20%polyacrylamide
gel
are shown below. Lanes 1-6, Ethidium bromidestaining
patterns; lanes7 and8,
autoradiographic
patterns. Lanes1-3,
7, AGP2primer;
lanes4-6, 8, AGP3primer. Templates
used: lanes 1 and4,AGP2genomic
DNA; lanes 2 and5, AGP3genomic
DNA; lanes 3, 6, 7, 8,single-stranded
cDNAderived from mouseliver.To overcome
this,
5'-end labeledprimers
wereemployed
for RT-PCR. As demonstratedby
thisanalysis,
thesignal
for AGP-3 isatleasttwoorders of
magnitude
weakerthan AGP2(Fig.
3b,
compare lowerpanel
lanes 7 and8).
DISCUSSION
We have isolated the
genomic
clones for the entirese-quenceof three ofthe M. domesticus AGP genes, AGP-1,
-2,
and -3. Thenewly
identified gene,AGP-3,
is locatedapproximately
6 kbupstreamfrom AGP-2.By
the restric-tionmapof the X clones ofAgp-1
andAgp-25
and theevi-dence that AGP-1is
proximal
tothecentromere(Baumann
et
ai,
1984),
wepredict
thegenomic organization
of these three genes is:AGP-1, AGP-3,
andAGP-2, arrayed
intandem. AGP-3 and AGP-2 are
closely
linked while thereis some distance
(not determined)
between AGP-2 and AGP-3.AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 AGP-1 AGP-2 AGP-3 CGGCAGGAGTCTGTGTCAGGACCAGT GGGCAGGAGTCTGTGTCAGGGCC-GG GGGCAGGAGCCTGTGTCACAGCCAGC -ieo -170 _16° AGGTTGAGGGAGCTGCATAAAGCTGG -CTGCGAGGGAGCTGCACAAAGCTGG TGGCTGAGAGAGCTGCACAAAGCTGG -ISO -140 C T T G A G G S A A íi A T T ! T G C G C A A G A C A CTTGAGAGAACATTTTGCGCAAGACA CTTGAGGGAACAT TTTG CACAAGA C A -130 -120 -110 TTTCCCAAGTGCTGGTGAGATTGTG- TTTCCCAAGTGCTGGGGAGAATGTG-TTTATCAAG TGCTGGTGAG T T T G T G G -100 -90 --C-CACAGCTCTACTGTCCCTGGCT --C-CAGGGCTCTAGAGGCCCTGGCT C A C TCACTGCTCTACAGCCCCTGGCT TCAGTCCCATGCCCTCCCCACATCG-GCAGTCCCATGCCCTCGCCACATC-T GCAGTCCCATGCCCTCCCCACATCCT -40 -30 A T A A A T A A A T A A ATGTTGCTGCACCATCC AAGCCACTGTACCCTC-AAGCCACTGCACCCTCC -20 -ÏÔ AACACCAATGATCTTTTCCCAGGCCT --CACCAGTTATCTCTTCCAAGCCCT GCCACCAGTTATCTCTTCCAAGCTCT +1 10 +20
FIG.4.
Comparison
ofsequencesin thepromoterregion
ofmouseAGP genes. The TATAboxisboxed. The
num-bers are indicated
according
to the AGP3 sequence. TheAGP/EBP
binding
motifsareindicatedby
solidunderlin-ing,
GREs are indicatedby overlining,
and theconsensusacute-phase
sequence is indicatedby
dashedunderlining.
AGP-3 mRNA
by
RT-PCR. AGP-3 mRNAwas detectedin this
experiment,
but thesignal
was weaker than thatofAGP-2.
Therefore,
the AGP-3geneis transcribedatavery lowlevel. Theexpression
of AGP-3protein
isyettobede-termined.
We do not know
why
AGP-3 is sopoorly expressed.
Onepossibility
isthat structuralaberrationsarepresent in the introns.Comparing
AGP-3andAGP-2,
themajor
dif-ferences are the lack of 86bp
of intron 1 and no(GT)28
tract in intron 5. The 86
bp
of intron 1 may contain thebranch site of
splicing;
the deletion of this sequence maycause AGP-3 RNA to be
spliced inefficiently.
Anotherpossible
explanation
is thatalternating purine-pyrimidine
sequencescanformZ-DNA,whichmayintroducea poten-tial siteforgene
regulation.
Alternatively,
the low-levelex-pression
may be due to thepositional
effect. Proudfoot(1986)
has shown thattranscription
ofthe first gene in a genecluster interfereswith thetranscription
ofthefollow-ing
gene. It ispossible
that thetranscription
of AGP-1 gene interferes with thetranscription
of AGP-3 gene.Two AGP
proteins
have been identifiedcorresponding
to the
products
of AGP-1 andAGP-2;
aprotein
product
ofAGP-3 has not been identified.
By
comparing
the de-duced amino acid sequenceofAGP-3and those of AGP-1 and AGP-2(Fig.
5),
AGP-3 would contain 206 aminoAGP-1 MALHTVLIILSLLPMLEAQNPEHAN 25
AGP-2 MALHMILVMVSLLPLLEAQNPEHVÑ
AGP-3
MELHTVLIMLSLLPLLEAQNPEHA-upstream from the
putative
transcription
initiation siteto130
bp
3' ofthepolyadenylation
signal.
Thisgenecontains6 exons and 56 introns. Its
coding
sequences are normaland contain no frameshift or nonsense mutations. The
exon/intron
splicing
sites,
andthe5'-flanking
and3'-flank-ing
sequences areconserved,
ascompared
to the AGP-2gene. The
5'-fianking
regions
of three AGPgenesaresimi-lar inthefirst 180
bp
upstream from thecapsite,
asshown inFig.
4. AGP-3aswellasAGP-1 andAGP-2contain the.potential glucocorticoid-responsive
element(-125
to-113)
and threeAGP/EBPbinding
sites locatedat -119to
-110,
-107 to-98,
and -87 to -78(Chang
etai,
1990).
A sequence of 38bp
(-15
to+23)
issimilarto se-quences observed in the three humanacute-phase proteins
(Dente
etai, 1985)
located inAGP-3gene.Part ofthereg-ulatory
region (-180
to+60)
of these three genes wereseparately
fusedto thechloramphenicol
acetyl
transferase(CAT) (Gorman
etai,
1982)
reportergeneand thentrans-fected intoaBHKcell line. The
expression
was monitoredby assaying
theactivity
ofthe CAT enzyme. Theactivity
of AGP-3 promoter is similar to those of AGP-1 andAGP-2
(data
notshown).
Using
RNaseprotection
toassay theendogenous
expres-sion ofthethree AGPgenes, weshowedthat theamount
ofAGP-1 wasabout fivefold
higher
thanAGP-2 inanor-malM. domesticus liver and
during
acute-phase
reaction.However,
theexpression
ofAGP-3 could not bedetectedby
the RNaseprotection
assay. Todetermineif the AGP-3 gene is a bonafide pseudogene
orjust
expressed
at very lowlevel,
wesynthesized specific
primers
toamplify
AGP-1 FTIGEPITNETLSWLSDKWFFMGAA 50 AGP-2 ITIGDPITÑETLSWLSDKWFFIGAA AGP-3 INIGDPITNETLSWLLGKWFLIAVA AGP-1 FRKLETRQAIQTMQSEFFYLTTNLI 75 AGP-2 VLNPDYRQEIQKTQMVFFNLTPNLI AGP-3 DSDPDYRQEIQKVQTIFFYLTLNLI AGP-1 NDTIELRESQTIGDQCVYNSTHLGF 100 AGP-2 NDTMELREYHTIDDHCVYNSTHLGI AGP-3 NDTMELREYHTKDDHCVYNSNLLGF AGP-1 QRENGTFSKYEGGVETFAHLIVLRK 125 AGP-2 QREÑGTLSKYVGGVKIFADLIVLKM AGP-3 LPEÑGTLFKYEGEVENPSHLRVLRK AGP-1 HGAFMLAFDLKDEKKRGLSLYAKRP 150 AGP-2 HGAFMLAFDLKDEKKRGLSLNAKRP AGP-3 HGAIMLFFDLKDEKKRGLSLSARRP AGP-1 DITPDLRDVFQKAVTHVGMDESEII 175 AGP-2 DITPDLRDVFQKAVTHVGMDESEII AGP-3 DIPPDLRDVFQKAVTHVGMDESEII AGP-1 FV3WKKDRCGQQEKKQLELGKETKK 200 AGP-2 FVDWKKDRCSQQEKQQLELEKETKK AGP-3 FVÛWKKDRCSEQEKKHLELEKETKK AGP-1 D P L £ G Q A AGP-2 D P E E G Q A AGP-3 D P E E S Q A
FIG. 5. The amino acid sequence of mouse
AGP-1,
-2,
and the
putative
AGP-3. Dots above thesingle-letter
amino acid mark theposition
ofputative glycosylation
sites(sequence
Asn-X-Ser/Thr).
320 CHANG ET AL.
acids,
including
the 18-residueputative signal peptide.
Thereare45amino acid substitutionsbetween the encoded AGP-3protein
and that ofAGP-1 or AGP-2. AGP is ahighly
glycosylated protein; therefore,
itwasofinteresttolocalize
potential
carbohydrate
attachment sites indicatedby
thesequenceof Asn-X-Thr/Ser. Fivepotential
sitescanbe foundin the AGP-1 and six
potential
sitesinAGP-2;
however,
only
threepotential glycosylation
sites existedinthe
putative
AGP-3polypeptide.
If AGP-3 expresses afunctional
protein,
theaminoacidsubstitutions would af-fectitsfunction. The existenceand thepotential
functionalimplications
of AGP-3protein
remained to beinvesti-gated.
ACKNOWLEDGMENTS
We thank Dr.
George
Bolton forediting
and Ms. Joanne Kahrmannand Ru-Ju Chen fortyping
thismanu-script.
This researchwassupported by
Grant NSC80-0412-B002-09 from the NationalScience Council.REFERENCES
BAUMANN, H., and MAQUAT, L.E. (1986). Localization of DNA sequence involved in dexamethasone-dependent expres-sion of the ratal-acid glycoprotein gene. Mol. Cell. Biol. 6, 2551-2561.
BAUMANN, H., HELD, W.A.,andBERGER,F.G.(1984).The
acute phase response of mouse liver. Genetic analysis of the majoracute phasereactants. J. Biol. Chem. 259,566-573.
BENNETT, M., and SCHMID, K. (1980). Immunosuppression byhumanplasmaod-acidglycoprotein:Importanceof the
car-bohydrate moiety. Proc. Nati. Acad. Sei. USA77,6109-6113.
CHANG, C.J., CHEN, T.T., LEI, H.Y., CHEN, D.S., and
LEE, S.C. (1990). Molecularcloningofatranscriptionfactor,
AGP/EBP, that belongs to members of the C/EBP family. Mol. Cell. Biol. 10, 6642-6653.
COOPER, R., and PAPACONSTANTINOU, J. (1986). Evi-dence for the existence ofmultipleai-acidglycoproteingenesin the mouse. J. Biol. Chem.261, 1849-1853.
COOPER, R., ECKLEY, D.M.,and PAPACONSTANTINOU, J. (1987). Nucleotidesequenceof themousea,-acid
glycopro-teingene 1. Biochemistry26, 5244-5250.
DENTE, L., GILBERTO, G., andCÓRTESE, R. (1985).
Struc-tureof the humanai-acidglycoproteingene:Sequence homol-ogywith other humanacutephaseproteingenes. Nucleic Acids
Res. 13, 3941-3952.
DENTE, L., PIZZA, M.G., METSPALU, A., and CÓRTESE,
R.(1987).Structure andexpressionof thegenescodingfor
hu-mana.-acid glycoprotein.EMBO J. 6, 2289-2296.
FRIEDMAN, M.L. (1983). Control of malaria virulenceby
a¡-acid glycoprotein (orosomucoid), an acute phase (inflamma-tory)reactant. Proc. Nati. Acad. Sei. USA80, 5421-5424.
GORMAN, CM., MOFFAT, L.F.,andHOWARD,B.H.(1982).
Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2, 1044-1051.
KLEIN, E.S., REINKE, R., FEIGELSON, P., and RINGOLD,
G.M. (1987). Glucocorticoid-regulated expressionfrom the 5'-flanking region of therat aracid glycoproteingene. J. Biol. Chem. 262, 520-523.
LEE, S.C, CHANG, C.J., LEE, Y.M., LEI, H.Y., LAI, M.Y.,
and CHEN, D.S. (1989). Molecular cloning of cDNAs
corre-spondingto twogenes ofaracidglycoproteinand characteriza-tion oftwo alíeles of AGP-1 in themouse. DNA8, 245-251.
LIAO, Y.C, TAYLOR, J.M., VANNICE, J.L., CLAWSON, G.A., and SMUCKLER, E.A. (1985). Structure of therat
a,-acid glycoproteingene. Mol. Cell. Biol. 5, 3634-3639.
LIU, H.M., TAKAGAKI, K.,andSCHMID, K. (1988).In vitro nerve-growth-promoting activityofhumanplasmaa,-acid gly-coprotein. J. Neurosci. Res. 20,64-72.
MELTON, D.A., KRIEG, P.A., REBAGLIATI, M.R.,
MANI-ATIS, T., ZINN, K., and GREEN, M.R. (1984). Efficient in vitrosynthesisofbiologicallyactive RNA and RNA
hybridiza-tionprobesfromplasmids containingabacteriophageSP6
pro-moter. Nucleic AcidsRes. 12, 7035-7056.
PROUDFOOT, N.J.(1986).Transcriptionalinterference and
ter-mination betweenduplicated a-globin geneconstructs suggests
anovel mechanism forgeneregulation. Nature322, 562-566.
PROWSE, K.R.,andBAUMANN,H.(1988). Hepatocyte-stimu-lating factor, a¡-interferon, and interleukin-1 enhance expres-sion of therat <x,-acidglycoproteingeneviaadistal upstream
regulatory region. Mol. Cell. Biol. 8,42-51.
REINKE, R., and FEIGELSON, P. (1985). Rat a,-acid glyco-protein. Gene sequence and regulation by glucocorticoids in transfected L-cell. J. Biol. Chem. 260,4397-4403.
SANGER, F., NICKLEN, S., and COULSON, A.R. (1977).
DNAsequencingwithchain-terminatinginhibitors.Proc.Nati. Acad. Sei. USA74,5463-5467.
SCHMID,K.(1975). InThe PlasmaProteins,vol. I. F.Putnam,
ed. (Academic Press,New York) pp. 184-228.
WON, K.-A.,andBAUMANN,H.(1990).Thecytokineresponse element of the rat a,-acid glycoprotein gene is a complex of
several interacting regulatory sequence. Mol. Cell. Biol. 10,
3965-3978.
Address
reprint
requests to:Dr.
Sheng-Chung
Lee Instituteof
Biological
Chemistry
Academia SínicaTaipei,
TaiwanReceived for publicationOctober 21, 1991, and in revised form December 3, 1991.