中 國 醫 藥 大 學
臨床醫學研究所
碩士學位論文
CCL5 增加肺癌細胞轉移經由 PI3K, Akt 和 NF-B 路徑
CCL5 increases lung cancer migration via PI3K, Akt and
NF-B pathways
指 導 教 授 :許弘昌
教
授
共同指導教授:湯智昕
助理教授
研 究 生: 黃俊寅
Abstract
Lung cancer is the leading cause of cancer-related mortality worldwide. It frequently results in distal metastases, including, brain, liver and bone marrow etc while it is diagnosed. Investigation of the factors and mechanism affecting tumor metastases is important in treatment of the disease. Chemokine plays a crucial role in the inflammatory response and the migration and metastasis of human cancer cells.
CCL5 (previously called RANTES) is in the CC-chemokine family. Previous studies show it related with activation of T-cell and metastases of breast tumor cells. Besides, integrins are the major adhesive molecules in mammalian cells and important in the migration of cells. In this study, we examine the effect of CCL5 on integrin expression and migration activity in human non-small cell lung cancer cells.
Here we found CCL5 increased the migration and cell surface expression of vintegrin in human lung cancer cells (A549 cells, H929 and H1299). The CCR5 of lung cancer cells have high expression than in lung epithelium cells (HBE-E6/E7 and BEAS-2B). Furthermore, we found CCL5 stimulation increased phosphorylation of the p85α subunit of phosphatidylinositol 3-kinase (PI3K) and serine 473 of Akt. Also, PI3K inhibitor (Ly294002) or Akt inhibitor suppressed CCL5-induced migration activities and integrin expression of A549 cells. Transfection of cells with p85 or Akt mutant also reduced CCL5-mediated cancer migration. In addition, treatment of A549 cells
phosphorylation, p65 Ser536 phosphorylation, and B-luciferase activity. Furthermore, the CCL5-mediated increases in p65 Ser536 phosphorylation were inhibited by Ly294002 and Akt inhibitor.
Taken together, our results suggest that CCL5 acts through PI3K/Akt, which in turn activates IKKand NF-B, resulting in the activation of v3 integrin and contributing to the migration of human lung cancer cells.
Chinese Abstract
肺癌是全世界造成癌症死亡的主因,它時常在診斷時就已經發 現有遠處的轉移。研究轉移的因素與機轉對疾病的治療是很重要的。 趨化素(chemokines)在人類的發炎反應及癌細胞的遷移和轉移 中扮演一個重要的角色。CCL5 (早期稱作 RANTES)為 C-C 趨化素 的一員。先前的研究顯示,它與 T 細胞的活化和乳癌細胞的轉移有 關。此外,integrins 為細胞的主要黏著分子,與細胞的遷移有密不可 分的關係。在本篇研究當中,我們探討 CCL5 對人類非小細胞肺癌 細胞(A549 細胞)中 integrins 的表現及其與遷移活性的影響。 我們發現,CCL5 可增加三株人類肺癌細胞(A549 細胞、H929 細胞和 H1299 細胞)的遷移,以及增加細胞表面vintegrin 的表 現。另外,肺癌細胞中 CCR5 的表現量也高於其他肺部表皮細胞 (HBE-E6/E7 和 BEAS-2B)。 更進一步,我們發現 CCL5 的刺激會增加 phosphatidylinositol 3-kinase(PI3K)中的 p85亞單位和 Akt serine 473 的磷酸化。而且, 我們加 PI3K 抑制劑 (Ly294002) 或者是 Akt 的抑制劑,可抑制因 CCL5 而產生的 A549 細胞遷移活動,及 A549 細胞 integrin 的表現。 用以 p85 顯性突變抑制物(dominant negative mutant)或者 Akt 顯性突 變抑制物,也降低由於 CCL5 所引起的癌細胞轉移。另外,經 CCL5 處理過之A549 細胞,會誘發 IB kinase (IKK)、IB 和 p65 Ser536磷酸化及B-luciferase 的活性。最後,Ly294002 和 Akt 抑制劑也抑制了CCL5 調節的 p65 Ser 536 磷酸化的增加。
綜合上述,我們的結果顯示 CCL5 會經由 PI3K/Akt 的路徑,來活 化IKK和NF-B 以致vintegrin 的表現,促使人類肺癌細胞的 轉移。
Acknowledgement
碩士論文總算進入撰寫謝辭的最後階段,研究所兩年多的時間終於 接近尾聲,想到當初懷著忐忑的心情進入臨床醫學研究所就讀,終 於可以對自己與身邊關心愛護我的人有所交代。 在整篇論文撰寫過程中,首先要感謝指導教授許弘昌老師,許主任 不僅是自己臨床工作的指導老師,更感謝老師在學生這兩年來的學 生生涯和論文寫作期間給予的悉心指導,並在學問和處世方面都帶 給我許多成長,給我充分的包容與支持。同時也要感謝口試委員湯 智昕老師和曾文培老師,在口試期間給予相當寶貴的建議與指正, 使這份論文能夠更臻完善,在此致上我最誠摯的謝意。 在中國醫藥大學臨床醫學研究所的這兩年研究生生涯中,感謝有碩 士班同學相互鼓勵和陪伴下,讓我在學業學習上不會感到孤單。特 別感謝湯智昕老師,在基礎醫學實驗上的指導與支持,引領我走進 實驗室這有趣的小小天地,讓我從中學得許多寶貴的經驗,對學生 影響至為深遠,在此對恩師至上最深之感謝。 最後,謹以此篇論文獻給我最愛的家人,感謝你們長久以來對我的支持與包容,讓我能無慮的大步的向前走,繼續實現自己夢想,在 此將所有的喜悅與成果與你們共同分享,將本論文獻給我最親愛的 家人。 更期許自己,未來能有更精進的表現。 黃俊寅 謹識於 中國醫藥大學臨床醫學研究所 中華民國九十七年十一月
Contents
Abstract ···2
Chinese Abstract ··· 4
Acknowledgement··· 5
Contents ··· 7
Figure contents··· 9
Abbreviation··· 10
Introduction··· 11
Materials and methods··· 16
Materials
··· 16
Cell culture
··· 18
Migration assay
··· 18
Flow Cytometric Analysis
··· 19
Western blot analysis
··· 20
Transfection and reporter gene assay
··· 20
Quantitative real time PCR and ELISA
··· 21
Statistics
··· 22
Results··· 23
CCL5-directed lung cancer cells migration involves avβ3 integrin up-regulation
··· 23
PI3K and Akt signaling pathways are involved in CCL5-mediated integrin upregulation and migration of lung cancer cells
··· 24
NF-κB signaling pathways are involved in CCL5-mediated integrin upregulation and migration activity
··· 25
Discussion··· 28
References··· 32
Figures··· 39
Fig 1. CCL5 induced migration activity of human lung cancer cells
··· 39
Fig 2. CCL5-directed migration activity of human lung cancer cells involves upregulation of αvβ3 integrins.
··· 41
Fig 3. PI3K is involved in CCL5-mediated migration and integrinupregulation in human lung cancer cells.
··· 43
Fig 4. Akt is involved in CCL5-mediated migration and integrinupregulation in human lung cancer cells.
··· 45
Fig 5. CCL5 induces cells migration and integrin upregulationthrough NF-κB.
··· 47
Fig 6. PI3K and Akt pathways are involved in CCL5-induced p65 phosphorylation and NF-κB activity.··· 49
Figures contents
Fig 1.
CCL5 induced migration activity of human lung cancer cellsFig 2.
CCL5-directed migration activity of human lung cancer cells involves upregulation of vβ3 integrins.Fig 3.
PI3K is involved in CCL5-mediated migration and integrin upregulation in human lung cancer cells.Fig 4.
Akt is involved in CCL5-mediated migration and integrin upregulation in human lung cancer cells.Fig 5.
CCL5 induces cells migration and integrin upregulation through NF-κB.Fig 6.
PI3K and Akt pathways are involved in CCL5-induced p65 phosphorylation and NF-B activity.Abbreviation
Akt Serine/threonine kinase or protein kinase B
CCL5 Chemokine (C-C motif) ligand 5
CCR5 Chemokine (C-C motif) ligand receptor 5
ECM Extracellular matrix
IKK α/β IκB kinaseα/β
NSCLC Non-small cell lung cancer
NF-κB nuclear factor-kappa B
PI3K phosphatidylinositol 3-kinase
PDTC Pyrrolidine dithiocarbamate
Introduction
Lung cancer is the leading cause of cancer-related mortality in both men and women (Jemal A et al., 2005), with 1.2 million new cases diagnosed every year and 1 million deaths recorded worldwide (Parkin DM et al., 2001). Non-small cell lung cancer (NSCLC) affects approximately 80% of all lung cancer patients. Most patients present with locally advanced (37%) or metastatic (38%) disease at time of diagnosis (Jemal A et al., 2005), and a large percentage of those diagnosed with early-stage disease eventually experience recurrence of metastatic disease. Thus, the high invasiveness of NSCLC to regional lymph nodes, liver, adrenal glands, contralateral lung, brain, and bone marrow, etc. may play a key role in its biological virulence (Jemal A et
al., 2005).
In the 1970s it was recognised that tumour-derived proteases, e.g. plasminogen activators and neutral matrix metalloproteinases, play an important role in the cleavage of basement membranes and extracellular matrix molecules and thus assist in the processes of invasion and metastasis of cancer cells (Granelli and Reich, 1978). The first chemokine identified as a chemotactic factor responsible for macrophage infiltration into tumours is MCP-1 (Mantovani A et al., 1992). It took several years to purify this monocyte chemotactic protein from tumour cells and to determine its primary structure (Yoshimura T et al., 1989 ; Mantovani A et al., 1992). Other tumour-derived monocyte chemotactic proteins, such as MCP-2/CCL8 and MCP-3/CCL7 have been discovered
considered to exert anti-tumour activity. Since CCR5, a chemokine receptor common for MIP-1α/CCL3,MIP-1β/CCL4 and RANTES/CCL5, is a marker for Th1 lymphocytes, these CCR5 ligands could be responsible for lymphocyte infiltration into tumours to help establish specific anti-tumour immunity. On the other hand, in a murine lymphoma model, autocrine production of MIP-1α and RANTES by activated lymphoma cells might be directly implicated in the reduced metastatic behaviour of these tumour cells by distracting lymphoma cells from being attracted by CCR1,3,5 agonists (and even other chemokines) produced in specific organs (Wang JM et al., 1998 ; Menten P et al., 2002)
Decades of scrutiny into the molecular bases of cancer have largely focused on what causes oncogenic transformation and the incipient emergence of tumors (Gupta and Massague , 2006). The invasion of tumor cells is a complex, multistage process. To facilitate cell motility, invading cells need to change cell-cell adhesion properties, rearrange the extracellular matrix (ECM) environment, suppress anoikis and reorganize their cytoskeletons (Woodhouse EC et al., 1997). Integrins are a family of transmembrane adhesion receptors comprising of 19 α and 8 β subunits that interact noncovalently to form up to 24 different heterodimeric receptors. The combination of different integrin subunits on the cell surface allows cells to recognize and respond to a variety of different ECM proteins including fibronectin, laminin, collagen and vitronectin (Humphries MJ, 2000). Because integrins are the primary receptors for cellular adhesion to ECM molecules, they act as crucial
transducers of bidirectional cell signaling, regulating cell survival, differentiation, proliferation, migration and tissue remodeling (Giancotti and Ruoslahti , 1999). Integrin has been heavily implicated in tumor development (Brooks PC et al., 1994 ; Brooks PC et al., 1995), and has been correlated to reduced patient survival in colon carcinoma and melanoma (Albelda SM et al., 1990 ; Hieken TJ et al., 1996), and associated with breast cancer cell metastasis to bone (Pecheur I et al., 2002). In addition, in vitro studies have found that integrins facilitated prostate cancer cell adhesion and migration through several ECM substrates (Cooper CR et al., 2002 ; Kumar CC, 2003), and transendothelial migration (Wang X et al., 2005).
Regulated upon Activation Normal T cell Expressed and Secreted (RANTES, CCL5), was originally recognized as a product of activated T cells (Schall TJ et al., 1988). A causal role was recently attributed to inflammation in many malignant diseases, including breast cancer and lung cancer (Gali and Adit, 2008). The different inflammatory mediators that are involved in this disease include cells, cytokines and chemokines. Of these, many studies have addressed the involvement and roles of the inflammatory chemokines CCL2 (MCP-1) and CCL5 (RANTES) in breast malignancy (Gali and Adit, 2008). While minimally expressed by normal breast epithelial duct cells, both chemokines are highly expressed by breast tumor cells at primary tumor sites, indicating that CCL2 and CCL5 expression is acquired in the course of malignant transformation, and suggesting that the two chemokines play a role in breast cancer development and/or progression (Gali and Adit, 2008).
Now widely established as an inflammatory chemokine, CCL5 is known to mediate chemotactic activity in T cells, monocytes, dendritic cells, natural killer cells, eosinophils and basophils (Schall TJ et al., 1990 ; Roth SJ et al., 1995 ; de la Rosa G et al., 2003). CCL5 is associated with chronic inflammatory diseases such as rheumatoid arthritis, inflammatory bowel disease and cancer (McCormack G et al., 2001 ; Ben-Baruch A, 2006). An association between CCL5 expression and cancer has been reported in melanoma, lung, prostate and pancreatic cancer (Luboshits G et al., 1999 ; Niwa Y et al., 2001 ; Yaal-Hahoshen N
et al., 2006). The most striking findings thus far have been with breast
cancer. Several investigations have reported that CCL5 was detected in samples from patients with breast cancer and that expression levels correlated with disease progression (Luboshits G et al., 1999 ; Niwa Y et
al., 2001 ; Yaal-Hahoshen N et al., 2006). The chemokine RANTES, its
role in the pathophysiology of disease and potential as a target for therapy.
We present cell types which are the source of RANTES, specific chemokine receptors and the role of chemokine in regulating the movement of cells into the tissues during inflammation. Also opinions on mechanisms regulating the chemokine secretion by inflammatory cytokines were discussed (Basiewicz-Worsztynowicz and Karnas-Kalemba, 2004). Chemokine RANTES is strongly induced by viral and bacterial infections and plays a role in allergic diseases, in asthma exacerbation, interstitial pneumonia, in allograft rejection and in cancers. This chemokine may represent target for diagnostic procedures
and therapeutic intervention, and may be useful as a prognostic factor in cancer (Basiewicz-Worsztynowicz and Karnas-Kalemba, 2004)..Previous
studies have shown that CCL5 modulates cell migration and invasion in several cancer cells (Ben-Baruch A, 2006 ; Karnoub AE et al., 2007).
However, the effect of CCL5 on integrin expression and migration activity in human non-small cell lung cancer cells is mostly unknown. Here we found a phenomenon whereby CCL5 increased the migration and expression ofαvβ3 integrin in human lung cancercells.In addition, phosphatidylinositol 3-kinase (PI3K), Akt, IKKand NF-κB signaling pathways were involved.
Materials and methods
Materials
Protein A/G beads Santa Cruz Biotechnology (Santa Cruz, CA, USA) Anti-mouse IgG-conjugated
horseradish peroxidase
Santa Cruz Biotechnology (Santa Cruz, CA, USA) Anti-rabbit IgG-conjugated
horseradish peroxidase
Santa Cruz Biotechnology (Santa Cruz, CA, USA) Rabbit polyclonal antibodies
specific for Akt
Santa Cruz Biotechnology (Santa Cruz, CA, USA) Rabbit polyclonal antibodies
specific for p85α Santa Cruz Biotechnology (Santa Cruz, CA, USA) Rabbit polyclonal antibodies
specific for IκB Santa Cruz Biotechnology (Santa Cruz, CA, USA) Rabbit polyclonal antibodies
specific for β-actin Santa Cruz Biotechnology (Santa Cruz, CA, USA) Rabbit polyclonal antibodies
specific for IKK Santa Cruz Biotechnology (Santa Cruz, CA, USA)
Ly294002 Calbiochem (San Diego, CA,USA) Akt inhibitor Calbiochem (San Diego, CA,USA) TPCK Calbiochem (San Diego, CA,USA) Pyrrolidine dithiocarbamate
(PDTC)
Calbiochem (San Diego, CA,USA)
Rabbit polyclonal antibody
specific for phosphor-p85 (Tyr458)
Cell Signaling
(Danvers, MA, USA) Rabbit polyclonal antibody
specific for phosphor-IKKα/β (Ser180/181)
Cell Signaling
Rabbit polyclonal antibody
specific for phosphor-p65 (Ser536) Cell Signaling
(Danvers, MA, USA)
Recombinant human CCL5 PeproTech (Rocky Hill, NJ, USA) Selectivev3 integrin antagonist Peptides International
(Louisville, KY, USA) cyclic RGD (cyclo-RGDfV)
peptide
Peptides International (Louisville, KY, USA) cyclic RAD (cyclo-RADfV)
peptide
Peptides International (Louisville, KY, USA) Mouse monoclonal antibody
specific for2 integrin Chemicon (Temecula, CA, USA) Mouse monoclonal antibody
specific for5 integrin Chemicon (Temecula, CA, USA) Mouse monoclonal antibody
specific for β1 integrin Chemicon (Temecula, CA, USA) Mouse monoclonal antibody
specific for21 integrin Chemicon (Temecula, CA, USA) Mouse monoclonal antibody
specific forv3 integrin Chemicon (Temecula, CA, USA) p85α(Δp85;deletion of35 amino
acids from residues 479-513 of p85)
Gifts from Dr. W. M. Fu (National Taiwan University, Taipei,
Taiwan) Akt (Akt K179A)
dominant-negative mutants
Gifts from Dr. W. M. Fu (National Taiwan University, Taipei,
Taiwan) IKKα (KM)and IKKβ (KM)
mutants
Gifts from Dr. H. Nakano (Juntendo University, Tokyo, Japan)
pSV-β-galactosidase vector Promega (Madison, MA, USA) Luciferase assay kit Promega (Madison, MA, USA)
Human lung adenocarcinoma cell lines (A549, H928 and H1299)
From the American Type Culture Collection (Manassas, VA, USA) Transwell, 8-μm Costar ( NY, USA )
Other chemicals Sigma-Aldrich (St. Louis, MO, USA)
Cell culture
The human lung adenocarcinoma cell lines (A549, H928 and H1299) were obtained from the American Type Culture Collection (Manassas, VA, USA). The cells were maintained in Dulbecco's Modified Eagle's Medium/Nutrient Mixture Ham's F12 (DMEM/F12) medium which was supplemented with 10% heat-inactivated Fetal Calf Serum (FCS), 2 mM-glutamine, penicillin (100 U/ml) and streptomycin (100 ng/ml) at 37°C with 5 % CO2. The human lung epithelium cell lines (HBE-E6/E7
and BEAS-2B) were obtained from the American Type Culture Collection (Manassas, VA, USA). The cells were cultured in DMEM/-MEM supplemented with 10% FCS and maintained at 37°C in a humidified atmosphere of 5 % CO2.
Migration assay
The migration assay was performed using Transwell (Costar, NY, USA; pore size, 8-μm) in 24-well dishes. Before performing the migration assay, cells were pretreated for 30 min with different concentrations of inhibitors, including the Ly294002 (10 μM), Akt inhibitor (10 μM), PDTC (10 μM), TPCK (3 μM) or vehicle control
(0.1% DMSO). The concentrations of inhibitors didn’taffect cell death of A549 cells shown by a cell viability assay (data not shown). Approximately 1×104 cells in 200 μlofserum-free medium were placed in the upper chamber, and 300 μlofthesame medium containing 3 ng/ml CCL5 was placed in the lower chamber. The plates were incubated for 24 h at 37°C in 5 % CO2, then cells were fixed in methanol for 15 min and
stained with 0.05 % crystal violet in phosphate buffer saline (PBS) buffer
for 15 min. Cells on the upper side of the filters were removed with cotton-tipped swabs, and the filters were washed with PBS. Cells on the underside of the filters were examined and counted under a microscope. Each clone was plated in triplicate in each experiment, and each experiment was repeated at least three times. The number of invading cells in each experiment was adjusted by the cell viability assay to correct for proliferation effects of CCL5 treatment (corrected invading cell number = counted invading cell number/percentage of viable cells) (Tang CH et al., 2008).
Flow Cytometric Analysis
Human lung cancer cells were plated in six-well dishes. The cells were then washed with PBS and detached with trypsin at 37°C. Cells were fixed for 10 min in PBS containing 1% paraformaldehyde. After rinsing in PBS, the cells were incubated with mouse anti-human antibody against integrins (1:100) for 1 h at 4°C. Cells were then washed again and incubated with fluorescein isothiocyanate-conjugated goat anti-rabbit secondary IgG (1:150; Leinco Tec. Inc., St. Louis, MO, USA) for 45 min
and analyzed by flow cytometry using FACS Calibur and CellQuest software (BD Biosciences, Palo Alto, CA, USA).
Western blot analysis
The cellular lysates were prepared as described previously (Lai TH
et al., 2008). Proteins were resolved on sodium dodecyl sulfate
-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to Immobilon polyvinyldifluoride (PVDF) membranes. The blots were blocked with 4% Bovine serum albumin (BSA) for 1 hr at room temperature and then probed with rabbit anti-human antibodies against IκBα,p-IκB,IKKα/β orp-Akt (1:1000) for 1 hr at room temperature. After three washes, the blots were subsequently incubated with a donkey anti-rabbit peroxidase-conjugated secondary antibody (1:1000) for 1 hr at room temperature. The blots were visualized by enhanced chemiluminescence using Kodak X-OMAT LS film (Eastman Kodak, Rochester, NY, USA).
Transfection and reporter gene assay
Human lung caner cells were co-transfected with 0.8 g B-luciferase plasmid, 0.4 g -galactosidase expression vector. A549 cells were grown to 80% confluence in 12 well plates and were transfected on the following day with Lipofectamine 2000 (LF2000; Invitrogen, Carlsbad, CA, USA). DNA and LF2000 were premixed for 20 min and then applied to cells. After 24 hr transfection, the cells were
then incubated with the indicated agents. After a further 24 hr incubation, the media were removed, and cells were washed once with cold PBS. To prepare lysates, 100 l reporter lysis buffer (Promega, Madison, WI, USA) was added to each well, and cells were scraped from dishes. The supernatant was collected after centrifugation at 13,000 rpm for 2 min. Aliquots of cell lysates (20 l) containing equal amounts of protein (20–30g) were placed into wells of an opaque black 96-well microplate. An equal volume of luciferase substrate was added to all samples, and luminescence was measured in a microplate luminometer. The value of luciferase activity was normalized to transfection efficiency monitored by the co-transfected-galactosidase expression vector.
Quantitative real time PCR and ELISA
The quantitative real time PCR (qPCR) analysis was carried out using Taqman® one-step PCR Master Mix (Applied Biosystems, CA, USA). 100 ng of total cDNA were added per 25-μl reaction with sequence-specific primers and Taqman® probes. Sequences for all target gene primers and probes were purchased commercially [glyceraldehyde-3- phosphate dehydrogenase (GAPDH) was used as internal control (Applied Biosystems, CA, USA)]. qPCR assays were carried out in triplicate with an ABI Prism 7900 sequence detection system. The cycling conditions were 10-min polymerase activation at 95 °C followed by 40 cycles at 95 °C for 15 s and 60 °C for 60 s. The threshold was set above the non-template control background and within the linear phase of target gene amplification in order to calculate the
cycle number at which the transcript was detected (denoted CT). CCL5
enzyme-linked immunosorbent assay (ELISA) was performed according to the manufacturer’s protocol(R&D Systems, Minneapolis, Minn, USA).
Statistics
The values given are means ± S.E.M. The significance of difference between theexperimentalgroupsand controlswasassessed by Student’s t test. The difference was significant if the p value was <0.05.
Results
CCL5-directed lung cancer cells migration involves avβ3 integrin up-regulation
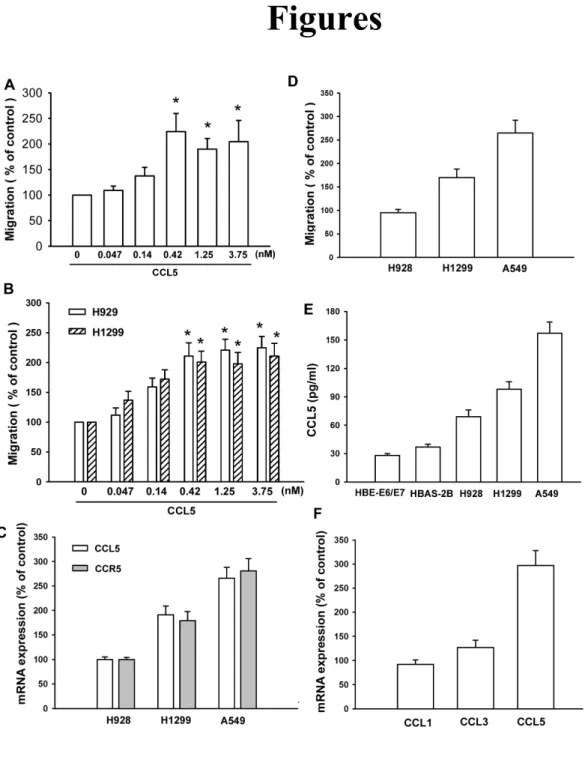
CCL5 has been reported to stimulate directional migration and invasion of human cancer cells (Ben-Baruch A, 2006 ; Karnoub AE et al., 2007). CCL5-trigered migration in lung cancer cells was examined using the Transwell assay with correction of CCL5-induced proliferation effects on human lung cancer cells (Tang CH et al., 2008 ; Lai TH et al., 2008). CCL5 directed human lung cancer cell (A549 cell) migration (Fig. 1A). On the other hand, CCL5 also increased the migration activity of the other lung cancer cell lines (H929 and H1299) (Fig. 1B). We then examined human lung cancer cell lines for expression of the CCL5 and CCL5 receptor (CCR5) by qPCR. qPCR revealed a higher level expression of CCL5 and CCR5 on A549 and a lower level in H928 cells (Fig. 1C). In addition, A549 cells were more invasive than H928 and H1299 (Fig. 1D). Expression of CCL5 in human lung cancer cell lines (H928, H1299 and A549) was significantly higher than in lung epithelium cells (HBE-E6/E7 and BEAS-2B) (Fig. 1E). Interaction of CCL5 with its specific receptor CCR on the surface of cancer cells has been reported to induce cancer invasion (Ben-Baruch A, 2006 ; Karnoub AE et al., 2007). However, A549 cells expressed a high level of CCR5 mRNA than CCR1 and CCR3 (Fig. 1F). Therefore, CCR5 is more important than CCR1 and CCR3 in lung cancer migration activity. The results indicated that the expression of CCL5/CCR5 axis was associated with an invasive and/or metastatic phenotype of lung cancer cell lines. Previous studies have shown significant expression of integrins in human
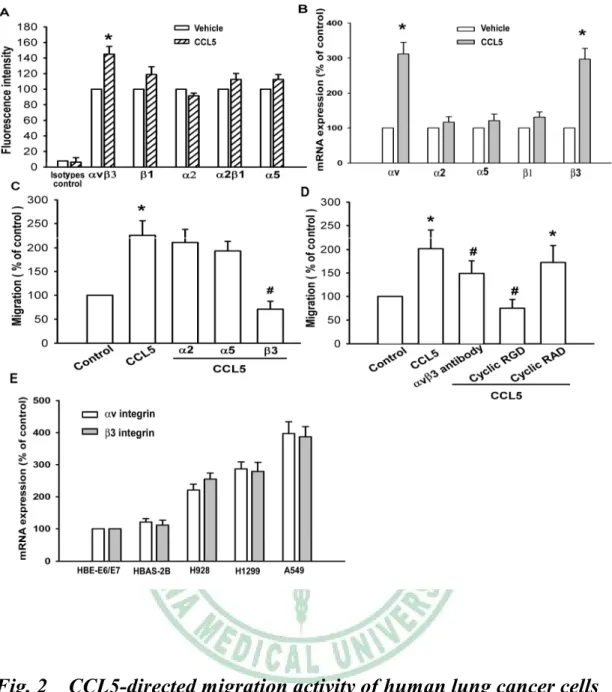
lung cancer cells (Huang YC et al., 2007 ; Fong YC et al., 2008). We hypothesized that integrins may be involved in CCL5-directed lung cancer cell migration. Flow cytometry analysis showed that CCL5-induced cell surface expression of vβ3 but not 1, 2, 5 or 21 integrin (Fig. 2A). In addition, CCL5 also increased the mRNA expression of v and β3 but not 1, 2 or 5 integrin (Fig. 2B). Pretreatment of cells with anti-but not or monoclonal antibody (mAb) (10 μg/ml)for 30 min markedly inhibited the CCL5-induced migration of lung cancer cells (Fig. 2C). In addition, vmAb also reduced CCL5-increased migration activity (Fig. 2D). The cyclic RGD peptide (cyclo-RGDfV) has been reported to bind v3 with high affinity and block its function effectively at low concentrations (Tang CH
et al., 2007). Treatment of cells with cyclic RGD but not cyclic RAD
inhibited CCL5-induced migration of lung cancer cells (Fig. 2D). In addition, expression of v and 3 integrin in human lung cancer cell lines (H928, H1299 and A549) was significantly higher than in lung epithelium cells (HBE-E6/E7 and BEAS-2B) (Fig. 2E). These data suggest that CCL5-induced cancer migration may occur via activation of vβ3 integrin receptor.
PI3K and Akt signaling pathways are involved in CCL5-mediated integrin upregulation and migration of lung cancer cells
PI3K/Akt can be activated by a variety of growth factors, such as insulin, nerve growth factors, and TGF-β1(Downard J, 1998 ; Horowitz JC et al., 2004). We examined whether CCL5 stimulation also enhanced
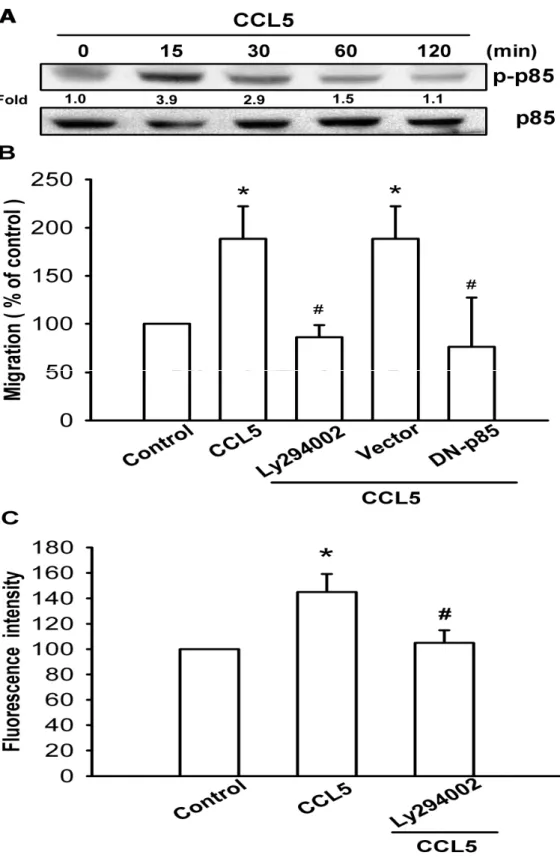
PI3K activation. Stimulation of A549 cells led to a significant increase in phosphorylation of p85 (Fig. 3A). CCL5-induced migration and vβ3 integrin expression of A549 cells were greatly reduced by treatment with Ly294002 (10 μM),aspecificPI3K inhibitor(Fig.3B&C). In addition, transfection of cells with p85αmutant also inhibited CCL5-induced migration of lung cancer cells (Fig. 3B). Ser473 residue phosphorylation of Akt by a PI3K-dependent signaling pathway causes enzymatic activation. To examine the crucial role of PI3K/Akt in cancer migration and integrin up-regulation, we next determined Akt Ser473 phosphorylation in response to CCL5 treatment. As shown in Fig. 4A, treatment of A549 cells with CCL5 resulted in time-dependent phosphorylation of Akt Ser473. Pretreatment of cells with Akt inhibitor (10 μM) antagonized CCL5-induced migration and vβ3 integrin expression of A549 cells (Fig. 4B&C). In addition, the Akt mutant also reduced CCL5-mediated cell migration (Fig. 4B).
NF-κB signaling pathways are involved in CCL5-mediated integrin
upregulation and migration activity
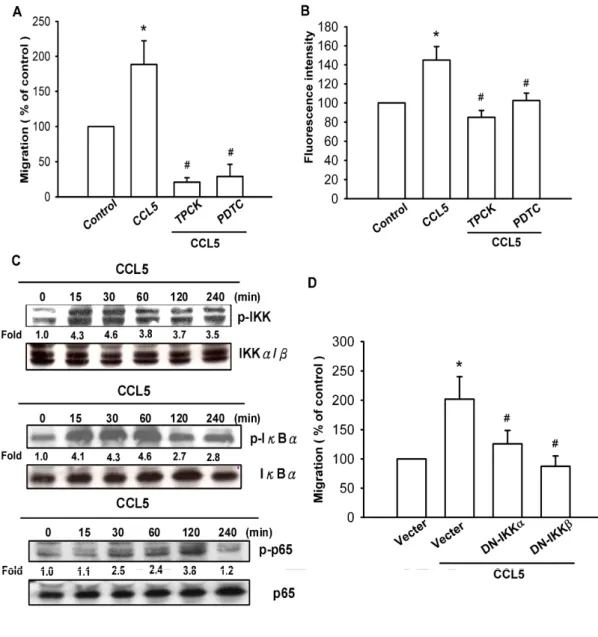
As previously mentioned, NF-κB activation is necessary for the migration and invasion of human cancer cells (Huang YC et al., 2007 ; Lai TH et al., 2008). To examine whether NF-κB activation isinvolved in CCL5-induced cancer migration, an NF-κB inhibitor,PDTC,wasused. Fig. 5A shows that A549 cells pretreated with PDTC (10 μM) and inhibited CCL5-induced lung cancer cell migration. Furthermore, A549 cells pretreated with TPCK (3μM), an IκB protease inhibitor,also
reduced CCL5-induced cancer cell migration (Fig. 5A). In addition, treatment of cells with PDTC or TPCK also antagonized CCL5-induced expression of vβ3 integrins (Fig. 5B). We further examined the upstream molecules involved in CCL5-induced NF-κB activation. Stimulation of cells with CCL5 induced IKKα/β phosphorylation in a time-dependent manner (Fig. 5C). Furthermore, transfection with IKKα orIKKβ mutantmarkedly inhibited CCL5-induced cancer cell migration (Fig. 5D). These data suggest that IKKα/β activation is involved in CCL5-induced migration activity of human lung cancer cells. Treatment of lung cancer cells with CCL5 also caused IκBαphosphorylation in a time-dependent manner (Fig. 5C). Previous studies showed that p65 Ser536phosphorylation increased NF-κB transactivation, and the specific antibody against phosphorylated p65 Ser536 was used to examine p65 phosphorylation (Madrid LV et al., 2001). Treatment of A549 cells with CCL5 for various time intervals resulted in p65 Ser536 phosphorylation (Fig. 5C).
To further investigate whether CCL5-induced p65 Ser536 phosphorylation, and NF-κB activation occurred through the PI3K/Akt pathway, A549 cells were pretreated for 30 min with Ly294002 and Akt inhibitor, which inhibited the CCL5-induced increase in p65 Ser536 phosphorylation as shown in Fig. 6A. In addition, the CCL5-induced increasein κB-luciferase activity was also inhibited by treatment with Ly294002, Akt inhibitor, PDTC and TPCK (Fig. 6B). Co-transfection with p85α, Akt, IKK and IKK mutants also reduced the CCL5-induced κB-luciferase activity (Fig. 6C).Taken together, these
Discussion
By far, lung cancer is the most common cause of cancer-related death in the world (Greenlee RT et al., 2001). Surgery remains the gold standard treatment for locoregional NSCLC, but unfortunately, only 15–20% of these tumors can be radically resected, and overall surgically-treated patient survival is only around 40% after 5 years (Maione P et al., 2004). Even in the early stages, the 5-year survival rate is only 60–65% after complete resection. This high mortality is probably attributable to early metastasis, principally spreading of malignant cells to many tissues including bone, particularly for NSCLC (Mundy GR et
al., 2002). Therefore, early detection of cancer and avoidance of cancer
metastasis demand immediate attention clinically. On the other hand, determining the mechanism of metastasis activity of cancer cells is a fundamentally important issue. To achieve metastasis, cancer cells must evade or co-opt multiple rules and barriers. Several discrete steps are discernible in the biological cascade of metastasis: loss of cellular adhesion, increased motility and invasiveness, entry and survival in circulation, exit into new tissue, and eventual colonization of a distant site (Gupta and Massague, 2006). The mechanism of metastasis is a complicated and multistage process, however our study showed that CCL5 promoted cell migration and the expression of vβ3 integrins in human lung cancer cells. We provide evidence that vβ3 integrin acts as crucial transducers of cell signaling, regulating cell migration and CCL5 act as a critical mediator of the metastasis activity of cancer cells in the tumor microenvironment.
expression, and presumably secreted CCL5/RANTES mediates its biological activities through activation of G protein–coupled receptors, CCR1, CCR3, or CCR5, and binds to glycosaminoglycans (Martin L et
al., 2001). Here we found that A549 cells expressed a higher level of
CCR5 mRNA than CCR1 and CCR3. Therefore, CCR5 is more important than CCR1 and CCR3 in the migration activity of lung cancer. RT-PCR revealed a higher level of expression of CCL5 and CCR5 in A549 and a lower level in H928 cells. In addition, A549 cells were more invasive than H928 and H1299. The results indicated that expression of CCL5/CCR5 axis was associated with an invasive and/or metastatic phenotype of lung cancer cell lines.
Integrins play critical roles in cell migration and adhesion. Integrins link the extracellular matrix to intracellular cytoskeletal structures and signaling molecules and are implicated in the regulation of a number of cellular processes, including adhesion, signaling, motility, survival, gene expression, growth and differentiation (Miranti and Brugge, 2002). Previous studies have shown that CCL5 modulates cell migration and invasion in several cancer cells (Ben-Baruch A, 2006 ; Karnoub AE et al., 2007). However, the expression of integrins by CCL5 in human lung cells is mostly unknown. We found that CCL5 increased vintegrin expression using flow cytometry analysis, which plays an important role during tumor metastasis. Furthermore, CCL5 also increased the cell surface presentation of vbut not ,5 or integrins. In the present study, we used vintegrin antibody to determine the role of vintegrin and found that it inhibited CCL5-induced cancer migration.
cyclic RAD inhibited the enhancement of invasion activity by CCL5, indicating the involvement ofv3 integrin in CCL5-mediated induction of cancer migration
A variety of growth factors stimulate the expression of integrin via signal-transduction pathways that converge to activate NF-κB complex of transcription factors (Boukerche H et al., 2007). The PI3K/Akt pathway is a major cascade mediating activation of the NF-κB signaling pathway in human cancer cells (Pan ZK et al., 1999). Phosphorylation of the p85αsubunit is required for activation of the p110 catalytic subunit of PI3K (Qureshi HY et al., 2007). We found CCL5-enhanced the p85α subunit phosphorylation in human lung cancer cells. Pre-treatment of cells with PI3K inhibitors LY294002 antagonized an increase in migration and integrin expression by CCL5 stimulation. This was further confirmed by the result that the dominant-negative mutant of p85α inhibited the enhancement of migration by CCL5. Moreover, we also found that CCL5 activated Akt Ser473 phosphorylation, while Akt inhibitor and Akt mutant inhibited CCL5-mediated cell migration. Our data indicates that PI3K/Akt could play an important role in the expression of integrin and migration of human lung cancer cells.
Many NF-κB activation pathwayshavebeen revealed, and all of them rely upon sequentially activated kinase cascades (Viatour P et al., 2005). The classical pathway is triggered by various pro-inflammatory cytokines such as IL-1β and TNF-α(Viatour P et al., 2005). These extracellular signals activate the IKK complex which phosphorylates IκBα atSer32 and Ser36 and signals for ubiquitin-related degradation. The
NF-κB-dependent transcription (Viatour P et al., 2005). Besides the phosphorylation and degradation of the IκB signal pathway, an IκB-independent pathway such as p65 phosphorylation for optimal NF-κB activation has been defined(Viatour P et al., 2005). p65 is phosphorylated at Ser536 by a variety of kinases through various signaling pathways, which enhances p65 transactivation potential. TNF-α induces rapid p65 phosphorylation at Ser536 through IKKs, resulting in increased transcriptional activity of p65 (Sakurai H et al., 1999). The results of this study show that the PI3K/Akt pathway contributes to CCL5-induced p65 Ser536 phosphorylation in A549 cells. CCL5-induced IKKα/β, IB phosphorylation and an increase in p65 phosphorylation at Ser536 which began at 15 and 120 min, respectively, while Ly294002 and Akt inhibitor inhibited CCL5-induced p65 phosphorylation at Ser536. CCL5 also enhanced phosphorylation of p85, Akt, IKK, IB and p65 dose-dependently (data not shown). These results indicate that PI3K/Akt may actthrough IKKα/β to increasep65 phosphorylation atSer536 and enhance NF-κB transactivation.
To conclude, we present a novel mechanism of CCL5-directed migration of lung cancer cells via upregulation of vβ3 integrin. CCL5 increases cells migration and integrin expression by activation of PI3K, Akt, IKK-α/β,and NF-κB-dependent pathway (Fig. 6D).
References
Albelda SM, Mette SA, Elder DE, Stewart R, Damjanovich L, Herlyn M, Buck CA. Integrin distribution in malignant melanoma: association of the beta 3 subunit with tumor progression. Cancer Res 1990,50:6757-6764.
Boukerche H, Su Z-z, Emdad L, Sarkar D, Fisher PB. mda-9/Syntenin regulates the metastatic phenotype in human melanoma cells by activating nuclear factor-kappaB. Cancer Res 2007,67:1812-1822. Ben-Baruch A. Inflammation-associated immune suppression in cancer:
the roles played by cytokines, chemokines and additional mediators. Semin Cancer Biol 2006,16:38–52
Brooks PC, Stromblad S, Klemke R, Visscher D, Sarkar FH, Cheresh DA. Antiintegrin alpha v beta 3 blocks human breast cancer growth and angiogenesis in human skin. J Clin Invest 1995,96:1815-1822.
Brooks PC, Montgomery AM, Rosenfeld M, Reisfeld RA, Hu T, Klier G, Cheresh DA. Integrin alpha v beta 3 antagonists promote tumor regression by inducing apoptosis of angiogenic blood vessels. Cell 1994,79:1157-1164.
Basiewicz-Worsztynowicz BM, Karnas-Kalemba WZ. Chemokine RANTES activity and its potential role as a target for diagnosis and therapy. 2004, 57(11-12):653-8
Cooper CR, Chay CH, Pienta KJ. The role of alpha(v)beta(3) in prostate cancer progression. Neoplasia (New York) 2002,4:191-194.
de la Rosa G, Longo N, Rodriguez-Fernandez JL, Puig-Kroger A, Pineda A, Corbi AL, Sanchez-Mateos P. Migration of human blood dendritic cells across endothelial cell monolayers: adhesion molecules and chemokines involved in subset-specific transmigration. J Leukoc Biol 2003,73:639–649
Fong YC, Hsu SF, Wu CL, Li TM, Kao ST, Tsai FJ, Chen WC, Liu SC, Wu CM, Tang CH. Transforming growth factor-beta1 increases cell migration and beta1 integrin up-regulation in human lung cancer cells. Lung Cancer 2008 In Press.
Gupta GP, Massague J. Cancer metastasis: building a framework. Cell 2006,127:679-695.
Giancotti FG, Ruoslahti E. Integrin signaling. Science 1999,285:1028-1032.
Greenlee RT, Hill-Harmon MB, Murray T, Thun M. Cancer statistics, 2001. CA Cancer J Clin 2001,51:15-36.
Gali Soria, Adit Ben-Baruch. The inflammatory chemokines CCL2 and CCL5 in breast cancer. Int J Cancer 2008, 267:271-285
Granelli-Piperno A, Reich E. A study of proteases and proteaseinhibitor complexes in biological fluids. J Exp Med 1978;148:223–34
Horowitz JC, Lee DY, Waghray M, Keshamouni VG, Thomas PE, Zhang H, Cui Z, Thannickal VJ. Activation of the pro-survival phosphatidylinositol 3-kinase/AKT pathway by transforming growth factor-beta1 in mesenchymal cells is mediated by p38 MAPK-dependent induction of an autocrine growth factor. J Biol Chem 2004,279:1359-1367.
cell-derived factor-1 enhances motility and integrin up-regulation through CXCR4, ERK and NF-kappaB-dependent pathway in human lung cancer cells. Biochem Pharmacol 2007,74:1702-12.
Hieken TJ, Farolan M, Ronan SG, Shilkaitis A, Wild L, Das Gupta TK. Beta3 integrin expression in melanoma predicts subsequent metastasis. J Surg Res 1996,63:169-173.
Humphries MJ. Integrin structure. Biochem Soc Trans 2000,28:311-339. Jemal A, Murray T, Ward E, Samuels A, Tiwari RC, Ghafoor A, Feuer
EJ, Thun MJ. Cancer statistics, 2005. CA Cancer J Clin 2005;55:10-30
Karnoub AE, Dash AB, Vo AP, Sullivan A, Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R, Weinberg RA. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature 2007,449:557-63.
Kumar CC. Integrin alpha v beta 3 as a therapeutic target for blocking tumor-induced angiogenesis. Curr Drug Targets 2003,4:123-131. Lai TH, Fong YC, Fu WM, Yang RS, Tang CH. Osteoblasts-derived
BMP-2 enhances the motility of prostate cancer cells via activation of integrins. Prostate. 2008,68:1341-53.
Luboshits G, Shina S, Kaplan O, Engelberg S, Nass D, Lifshitz-Mercer B, Chaitchik S, Keydar I, Ben-Baruch A. Elevated expression of the CC chemokine regulated on activation, normal T cell expressed and secreted (RANTES) in advanced breast carcinoma. Cancer Res 1999,59:4681–4687
inflammatory bowel disease. Inflamm Res 2001,50:491–495
Madrid LV, Mayo MW, Reuther JY, Baldwin AS, Jr. Akt stimulates the transactivation potential of the RelA/p65 Subunit of NF-kappa B through utilization of the Ikappa B kinase and activation of the mitogen-activated protein kinase p38. J Biol Chem 2001,276:18934-18940.
Maione P, Rossi A, Airoma G, Ferrara C, Castaldo V, Gridelli C. The role of targeted therapy in non-small cell lung cancer. Crit Rev Oncol Hematol 2004,51:29-44.
Mundy GR. Metastasis to bone: causes, consequences and therapeutic opportunities. Nat Rev Cancer 2002,2:584-593.
Miranti CK, Brugge JS. Sensing the environment: a historical perspective on integrin signal transduction. Nat Cell Biol 2002,4:E83-90.
Martin L, Blanpain C, Garnier P, Wittamer V, Parmentier M, Vita C. Structural and functional analysis of the RANTES-glycosaminoglycans interactions. Biochemistry 2001;40:6303–18.
Mantovani A, Bottazzi B, Colotta F, Sozzani S, Ruco L. The origin and function of tumour-associated macrophages. Immunol Today 1992;13:265–70.
Matsushima K, Larsen CG, DuBois GC, Oppenheim JJ. Purification and characterization of a novel monocyte chemotactic and activating factor produced by a human myelomonocytic cell line. J Exp Med 1989;169:1485–90.
metastatic behaviour of murine T cell lymphoma. J Leukoc Biol 2002;72:780–9.
Niwa Y, Akamatsu H, Niwa H, Sumi H, Ozaki Y, Abe A. Correlation of tissue and plasma RANTES levels with disease course in patients with breast or cervical cancer. Clin Cancer Res 2001,7:285–289
Pan ZK, Christiansen SC, Ptasznik A, Zuraw BL. Requirement of phosphatidylinositol 3-kinase activity for bradykinin stimulation of NF-kappaB activation in cultured human epithelial cells. J Biol Chem
1999,274:9918-9922.
Pecheur I, Peyruchaud O, Serre C-M, Guglielmi J, Voland C, Bourre F, Margue C, Cohen-Solal M, Buffet A, Kieffer N, Clezardin P. Integrin alpha(v)beta3 expression confers on tumor cells a greater propensity to metastasize to bone. FASEB J 2002,16:1266-1268.
Parkin DM, Bray F, Ferlay J, Pisani P. Estimating the world cancer burden: Globocan 2000. Int J Cancer 2001,94:153-156.
Qureshi HY, Ahmad R, Sylvester J, Zafarullah M. Requirement of phosphatidylinositol 3-kinase/Akt signaling pathway for regulation of tissue inhibitor of metalloproteinases-3 gene expression by TGF-beta in human chondrocytes. Cell Signal 2007,19:1643-1651.
Roth SJ, Carr MW, Springer TA. C–C chemokines, but not the C–X–C chemokines interleukin-8 and interferon-gamma inducible protein-10, stimulate transendothelial chemotaxis of T lymphocytes. Eur J Immunol 1995,25:3482–3488.
Schall TJ, Bacon K, Toy KJ, Goeddel DV. Selective attraction of monocytes and T lymphocytes of the memory phenotype by cytokine
Schall TJ, Jongstra J, Dyer BJ, Jorgensen J, Clayberger C, Davis MM, Krensky AM. A human T cell-specific molecule is a member of a new gene family. J Immunol 1988,141:1018–1025
Sakurai H, Chiba H, Miyoshi H, Sugita T, Toriumi W. IkappaB kinases phosphorylate NF-kappaB p65 subunit on serine 536 in the transactivation domain. J Biol Chem 1999,274:30353-30356.
Tang CH, Tan TW, Fu WM, Yang RS. Involvement of matrix metalloproteinase-9 in stromal cell-derived factor-1/CXCR4 pathway of lung cancer metastasis. Carcinogenesis 2008,29:35-43
Tang CH, Lu DY, Tan TW, Fu WM, Yang RS. Ultrasound induces hypoxia-inducible factor-1 activation and inducible nitric-oxide synthase expression through the integrin/integrin-linked kinase/Akt/mammalian target of rapamycin pathway in osteoblasts. J Biol Chem. 2007,282:25406-15.
Viatour P, Merville M-P, Bours V, Chariot A.: Phosphorylation of NF-kappaB and IkappaB proteins: implications in cancer and inflammation. Trends Biochem Sci 2005;,30:43-52.
Van Damme J, Proost P, Lenaerts J-P, Opdenakker G. Structural and functional identification of two human, tumour-derived monocyte chemotactic proteins (MCP-2 and MCP-3) belonging to the chemokine family. J Exp Med 1992;176:59–65.
Wang JM, Chertov O, Proost P, Li JJ, Menten P, Xu L, et al. Purification and identification of chemokines potentially involved in kidney-specific metastasis by a murine lymphoma variant: induction of migration and NFkappaB activation. Int J Cancer 1998;75:900–7.
facilitate matrix interactions during transendothelial migration of PC3 prostate tumor cells. Prostate 2005,63:65-80.
Woodhouse EC, Chuaqui RF, Liotta LA. General mechanisms of metastasis. Cancer 1997,80:1529-1537.
Yaal-Hahoshen N, Shina S, Leider-Trejo L, Barnea I, Shabtai EL, Azenshtein E, Greenberg I, Keydar I, Ben-Baruch A. The chemokine CCL5 as a potential prognostic factor predicting disease progression in stage II breast cancer patients. Clin Cancer Res 2006,12:4474–4480 Yoshimura T, Robinson EA, Tanaka S, Appella E, Kuratsu JI, Leonard EJ. Purification and amino acid analysis of two human glioma-derived monocyte chemoattractants. J Exp Med1989;169:1449–59.
Figures
Fig. 1 CCL5 induced migration activity of human lung cancer cells
(A) A549 cells were incubated with various concentrations of CCL5, and in vitro migration activity measured with the Transwell after 24 hr showed all supported the A549 cells migration in a dose-dependent way. (B) H928 and H1299 cells were incubated with various concentrations of CCL5, and in vitro migration activity measured with the Transwell after 24 hr. (C) Total RNA were extracted from H928, H1299 and A549 cell lines and subjected to qPCR analysis for CCL5 and CCR5. (D) The migration activity of each cell line measured in vitro with the Transwell after 24 hr showed a significantly higher migration activity in A549 cell lines as compared with H928 or H1299 cell. (E) Total RNA were extracted from H928, H1299 and A549 cells lines and subjected to qPCR analysis for CCL5 and CCR5. (F) The amounts of CCL5 in culture medium (HBE-E6/E7, HBAS-2B, H928, H1299 and A5489) were determined with CCL5 ELISA kit. (F) Total RNA were extracted from A549 cells lines and subjected to qPCR analysis for CCR1, CCR3 and CCR5. Results are expressed as the mean ± S.E. *, p < 0.05 compared with control.
Fig. 2 CCL5-directed migration activity of human lung cancer cells
Fig. 2 CCL5-directed migration activity of human lung cancer cells
involves upregulation ofvβ3 integrins.
(A) A549 cells were incubated with CCL5 (3 ng/ml) for 24 hr, and the cell surface expression of v3, β1,2, 21 and 5 integrin was determined using flow cytometry. (B) A549 cells were incubated with CCL5 (3 ng/ml) for 24 hr, and the mRNA expression of v, 2, β1and integrin was determined using qPCR. (C) A549 cells were pretreated with2, 5 and β3 monoclonal antibody (10 μg/ml)for30 min,followed by stimulation with CCL5 (0.42 nM). The in vitro migration activity measured after 24 hr. (D) A549 cells were pretreated with v3 monoclonal antibody (10 g/ml), cyclic RGD (100 nM) or cyclic RAD (100 nM) for 30 min followed by stimulation with CCL5. The in vitro migration activity measured after 24 hr. (E) Total RNA were extracted from HBE-E6/E7, HBAS-2B, H928, H1299 and A549 cells lines and subjected to qPCR analysis for v and 3 integrin.*, p < 0.05 compared with control; #, p < 0.05 compared with CCL5-treated group.
Fig. 3 PI3K is involved in CCL5-mediated migration and integrin upregulation in human lung cancer cells.
Fig. 3 PI3K is involved in CCL5-mediated migration and integrin upregulation in human lung cancer cells.
(A) A549 cells were incubated with CCL5 (0.42 nM) for indicated time intervals, p-p85 expression was examined by Western blot analysis. (B) A549 cells were pretreated for 30 min with Ly294002 (10 μM)or transfected with DN mutantofp85αfor 24 hr. Then they were followed by stimulation with CCL5 (0.42 nM), and in vitro migration was measured with the Transwell after 24 hr. (C) Cells were pretreated for 30 min with Ly294002 (10 μM), and then stimulation with CCL5 (0.42 nM), and the cell surface v3 integrin was measured by using flow cytometry. Results are expressed as the mean ± S.E. *, p < 0.05 compared with control; #, p < 0.05 compared with CCL5-treated group.
Fig. 4 Akt is involved in CCL5-mediated migration and integrin upregulation in human lung cancer cells.
Fig. 4 Akt is involved in CCL5-mediated migration and integrin upregulation in human lung cancer cells.
(A) A549 cells were incubated with CCL5 (0.42 nM) for indicated time intervals, and p-Akt expression was determined by Western blot analysis. (B) A549 cells were pretreated for 30 min with Akt inhibitor (10 μM)ortransfected with DN mutant of Akt for 24 hr. Then they were followed by stimulation with CCL5 (0.42 nM), and in vitro migration was measured with the Transwell after 24 hr. (C) Cells were pretreated for30 min with Aktinhibitor(10 μM)and then stimulation with CCL5 (0.42 nM), and the cell surface v3 integrin was measured by using flow cytometry. Results are expressed as the mean ± S.E. *, p < 0.05 compared with control; #, p < 0.05 compared with CCL5-treated group.
Fig. 5 CCL5 induces cells migration and integrin upregulation through NF-κB.
Fig. 5 CCL5 induces cells migration and integrin upregulation through NF-κB.
(A) A549 cells werepretreated for30 min with PDTC (10 μM)or TPCK (3 μM)followed by stimulation with CCL5 (0.42 nM), and in vitro migration was measured with the Transwell after 24 hr. (B) A549 cellswerepretreated for30 min with PDTC (10 μM)orTPCK (3 μM) followed by stimulation with CCL5 (0.42 nM) for 24 hr, and the cell surface v3 integrin was measured by flow cytometry. (C) A549 cells were incubated with CCL5 (0.42 nM) for indicated time intervals, and p-IKKα/β,p-IBand p-p65 expression was determined by Western blot analysis. (D) A549 cells were transfected with DN mutantofIKKα or IKKβfor 24 hr and then followed by stimulation with CCL5 (0.42 nM), and in vitro migration was measured with the Transwell after 24 hr. Results are expressed as the mean ± S.E. *, p < 0.05 compared with control; #, p < 0.05 compared with CCL5-treated group.
Fig. 6 PI3K and Akt pathways are involved in CCL5-induced p65 phosphorylation and NF-B activity.
Fig. 6 PI3K and Akt pathways are involved in CCL5-induced p65 phosphorylation and NF-B activity.
(A) A549 cells were pretreated with Ly294002 (10 μM), Akt inhibitor(10 μM)for30 min beforetreatmentwithCCL5 (0.42 nM) for another 60 min, after which p65 Ser536 phosphorylation were determined by immunoblotting with antibodies specific for phospho-p65. (B&C) A549 cells transiently transfected with κB-luciferase plasmid for 24 h were either cotransfected with p85α,Akt,IKKα and IKKβ mutantsor pretreated with Ly294002 (10 μM),Aktinhibitor(10 μM),TPCK (10 μM)orPDTC (3 μM)for 30 min, before incubation with CCL5 (0.42 nM) for 24 h. Luciferase activity was measured, and the results were normalized to the β-galactosidase activity. Results are expressed as the mean ± S.E. *, p < 0.05 compared with control; #, p < 0.05 compared with CCL5-treated group. (D) Schematic presentation of the signaling pathways involved in CCL5-induced migration and integrins expression of lung cancer cells. CCL5 activates PI3K and Akt pathway, which in turn induces IKK phosphorylation, p65 phosphorylation, and NF-B activation, which leads to v3 integrins expression and increases the migration of human lung cancer cells.