Original Article
Appearance of acanthosis nigricans may precede obesity: An
involvement of insulin/IGF receptor signaling pathway
C. H. Wang1,#, W. D. Lin2,#, D. T. Bau3, I. C. Chou1, C. H. Tsai1, , F. J.
Tsai1,2,4
1
Departments of Pediatrics,
2Medical Research, and
3Terry Fox
Cancer Research Laboratory, China Medical University
Hospital, Taichung, Taiwan
4
College of Health Science, Asia University, Taichung, Taiwan
# The authors contribute equally to this study
Correspondence to: Fuu-Jen Tsai, China Medical University Hospital, 2 Yuh-Der Road, Taichung 40402, Taiwan. Tel. +886-4-22052121 ext. 2041, E-mail:
Key Words: Acanthosis nigricans, obesity, insulin, polymorphism Running title: Acanthosis nigricans may precede obesity
Abstract
Obesity is one of the leading causes of preventable death. Complication of child obesity include cardiovascular risks, type 2 diabetes mellitus, impaired glucose tolerance, and acanthosis nigricans (AN). AN is associated with obesity as a manifestation of cutaneous insulin resistance, while the interaction between AN and obesity and detail mechanism for the pre- and co-obese appearance of AN in child are still unrevealed. In this pioneer study, the involvement of insulin/IGF receptor pathway in child pre- and co-obese AN was investigated via studying the association of polymorphisms of INSR, IRS1, IGF1R genes with pre- and co-obese AN. In total, 99 children pre- and co-obese AN patients and 100 healthy controls recruited were genotyped and analyzed by PCR-RFLP method. Significantly different distributions were found in the frequency of the INSR His1085His and IGF1R IVS7-20 genotypes, but not in IRS1 Ala804Ala or IGF1R Thr766Thr genotypes, between the AN and control groups. The T allele of INSR His1085His and the C allele of IGF1R IVS7-20 both conferred a significant (p=0.04 and 2.84E-6, respectively) increased risk of AN. Our results provide not only the evidence of the T allele of INSR His1085His and the C allele of IGF1R IVS7-20 are correlated with the appearance of AN precede or concurrence with obesity but revealed that insulin/IGF receptor pathway may play an important role in this pre- and co-obese AN.
Introduction
Obesity is one of the leading causes of preventable death. As more intake of high caloric content food and sedentary lifestyle, pediatric obesity is becoming more and more prevalence and pose a major impact on public health in the 21st century that may originate from childhood. This universal trend of pediatric obesity is reported not only in the occidental countries, up to 25 percent of children in USA, but also in oriental countries including Taiwan (1). There is no denying that obese children tend to become obese adults especially whose obesity lasts to their adolescence. Complication of obesity include cardiovascular risks, hypertension, dyslipidemia, endothelial dysfunction, type 2 diabetes mellitus and impaired glucose tolerance, acanthosis nigricans (AN), hepatic steatosis, precocious puberty, hypogonadism and polycystic ovary syndrome, obstructive sleep disorder, orthopedic complications, cholelithiasis and pseudotumor cerebri (2–4).
AN, is a hyperpigmented, with velvety cutaneous thickening obviously observed on certain parts of the body, including axilla, sides of neck, the groin, antecubital and poplitdal surface, umbilical area, and in more
severe cases even spread over the whole body and mucosal surface. In the literature, AN is reported to be closely associated with obesity as a manifestation of cutaneous insulin resistance (5). In addition, endocrinopathies, malignancy (most frequently gastric carcinoma in adult), genetic syndromes and the use of drugs may also develop AN as well (6–10).
From the clinical viewpoint, it is taken for granted that obesity often comes before the appearance of AN of cutaneous insulin resistance. Interestingly, from the questionnaires of the child AN patients we met in China Medical University Hospital, a large proportion (about 50%) of their AN syndromes comes before or together with obese, which hereafter will be mentioned as pre- and co-obese AN (PCOAN). This clinical observation may challenge the traditional rationale that elevated insulin concentrations owing to excessive weight gain and subsequent insulin resistance in obese people result in direct and indirect activation of IGF-1 receptors on keratinocytes and fibroblasts, leading to epidermal proliferation and the appearance of AN (11) Insulin and leptin resistance had been proved to be responsible for failure of appetite and excessive
energy intake suppression (12). In addition, defects in insulin binding and post-insulin receptor function, and genetic defects within the insulin receptor gene had been documented in patients with AN (13–16). From our clinical experience and the limited previous reports, we hypothesized that genetic background difference of insulin/IGF receptor and its associated signaling pathway in these PCOAN children should play an important role in the formation of excessive weight gain and AN, no matter obesity or AN occurs first. Insulin resistance itself could precipitate excessive fattiness that stem from failure of prevention of redundant energy intake, facilitating the emergence of AN that resulted from epidermal proliferation at the same time. The overall flowchart of our hypothesis is demonstrated in Figure 1 (Fig.1).
In order to understand the genomic role of insulin/IGF receptor pathway related genes in PCOAN, we have chosen four single nucleotide polymorphisms from three of the genes, INSR His1085His (rs1799817), IRS1 Ala804Ala (rs1801123), IGF1R IVS7-20 (rs2272037), IGF1R Thr766Thr (rs3743262), and investigated their genotypic distribution in a Taiwanese PCOAN child population.
Materials and Methods
Study Population and Sample Collection
Study group’s children and adolescents (ranged 5-18 years old) were recruited from the Department of Pediatrics of China Medical University Hospital in central Taiwan. BMI was calculated as weight (in kilograms) divided by height (in meters) squared. All subjects with a BMI above the 95th percentile for age and gender were classified as obese on the basis of BMI category by the Department of Health of Taiwan. A morning serum after an 8 h fast was obtained for glucose and insulin. Insulin resistance was determined by the homeostasis model assessment for insulin resistance (HOMA-IR) index in the fasting state. Detailed medical and family histories were obtained for all subjects, and physical examinations were performed. All subjects were of normal thyroid and adrenal functions. The subjects in the control group were enrolled from the Department of Family Medicine which were free of obese, AN, and other obese-related diseases. Each person donated the samples with an informed consent and the study was approved by the IRB committee of China Medical University Hospital.
Genotyping Assays. Genomic DNA was prepared from peripheral blood
leucocytes using a QIAamp Blood Mini Kit (Blossom, Taipei, Taiwan) and further processed according to previous studies (17–25). Briefly, the following primers were used for:
INSR C3255T rs1799817: 5’-TTGGGTGAGGGCTTGGGTGGAAG-3’
and 5’-CCTGTGTCCTCTGTCGCTCTGTG-3’;
IRS1 C2412T rs1801123: 5’-CTCCTACTACTCATTGCCAAG-3’ and
5’-CAGACAAGTAGCCAGACTGAT-3’;
IGF1R IVS7-20(C/T) rs2272037:
5’-GACCTCCCATTATAGAAAGTG-3’ and 5’-CCAGTGAGCTTGCGAAGAAG-5’-GACCTCCCATTATAGAAAGTG-3’ and
IGF1R C2298T rs3743262: 5’-TCCACGGTTA AGATTCTTCTG-3’ and
5’-TCCACTAGGTTGTGAGGAAG-3’.
The following cycling conditions were performed: one cycle at 94°C for 5 min; 35 cycles of 94°C for 30 s, 58°C for 30 s, and 72°C for 30 s; and a final extension at 72°C for 10 min. The PCR products, except IGF1R IVS7-20(C/T) rs2272037, were studied after digestion with MspA1I,
HphI and DraIII, restriction enzymes for IRS1 C2412T rs1801123 (cut
rs3743262 (cut from 432 bp T type into 181+251 bp C type) and INSR C3255T rs1799817 (cut from 418 bp T type into 92+326 bp C type), respectively. The PCR product of IGF1R IVS7-20(C/T) rs2272037 was purified using QIAEX II (QIAGEN, Hilden, Germany) and applied to direct sequencing for the SNP type detection. Direct sequencing was conducted using the BigDye 3.1 Terminator cycle sequencing kit (Applied Biosystems, Foster City, CA, U.S.A.) with the ABI 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, U.S.A.).
Statistical Analyses. In our study, only those matches with all SNPs data
(case/control=99/100) were selected into final analyzing. Pearson’s two-sided c2 test or Fisher’s exact test was used to compare the distribution of the genotypes between cases and controls. Data was recognized as significant when the statistical p was less than 0.05.
Results
The clinical characteristics and analysis of recruited 99 PCOAN child patients and 100 healthy controls are shown in Table I. These two groups were similar in gender enrollment. Control group had advanced age, but even so, this group was of lower BMI (kg/m2), lower serum fasting
glucose, lower insulin level and lower HOMA (p<0.005) , meaning that they are of higher insulin sensitivity. In the case that insulin resistance is defined as fasting insulin levels higher than 15 U/ml, none of the control group, but 61 out of the 99 patients in the PCOAN group are insulin resistant. On the other hand, 14 patients had impaired fasting glucose and 4 of them fulfilled the criteria of diabetes (Table I). The meanings of these differences will be further discussed in the Discussion section.
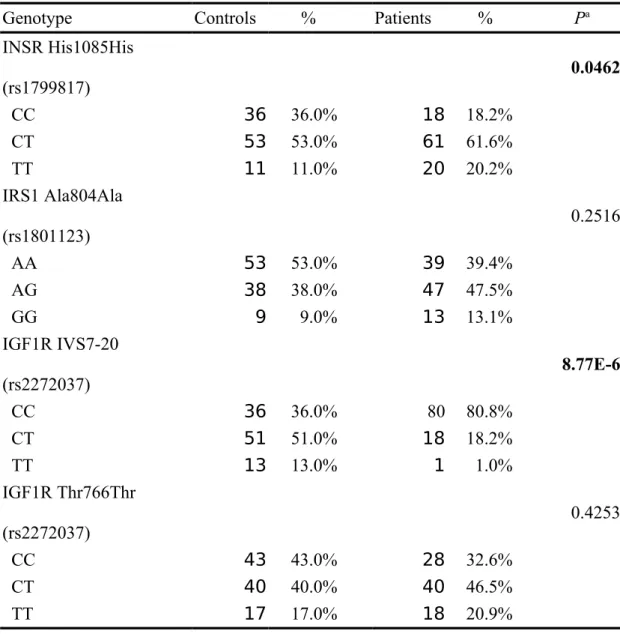
The frequencies of the genotypes for INSR His1085His (rs1799817), IRS1 Ala804Ala (rs1801123), IGF1R IVS7-20 (rs2272037), IGF1R Thr766Thr (rs3743262), between controls and PCOAN patients are shown in Table II. Genotype distributions of various genetic polymorphisms of INSR His1085His and IGF1R IVS7-20 are significantly different between PCOAN patients and control groups (p=0.046 and 8.77E-6, respectively), while those for IRS1 Ala804Ala or IGF1R Thr766Thr were not significant (p>0.05) (Table II). In detail, distributions of INSR His1085His *C homozygote/heterozygote/T homozygote in controls and PCOAN patients are 36.0/53.0/11.0% and 18.2/61.6/20.2%, respectively (Table II). Proportions of IRS1 Ala804Ala *C homozygote/heterozygote/T homozygote in controls and PCOAN
patients are 36.0/51.0/13.0% and 80.8/18.2/1.0%, respectively (Table II). Proportions of IGF1R IVS7-20 *A homozygote/heterozygote/G homozygote in controls and PCOAN patients are 53.0/30.0/9.0% and 39.4/47.5/13.1%, respectively (Table II). Proportions of IGF1R Thr766Thr *C homozygote/heterozygote/T homozygote in controls and PCOAN patients are 43.0/40.0/17.0% and 32.6/46.5/20.9%, respectively (Table II). To sum up, the polymorphism of IGF1R IVS7-20 and INSR His1085His are significantly associated with PCOAN occurrence.
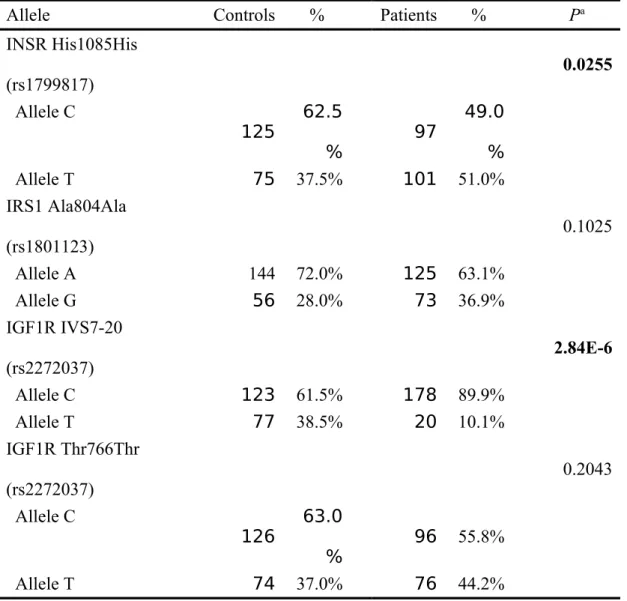
The frequencies of the alleles for the IRS1 Ala804Ala (rs1801123), IGF1R IVS7-20 (rs2272037), IGF1R Thr766Thr (rs3743262), INSR His1085His, between controls and PCOAN patients are shown in Table III. Allele frequency distributions of the INSR His1085His and IGF1R IVS7-20 *C are associated with higher susceptibility for PCOAN occurrence. In detail, distributions of INSR His1085His C/T allele in controls and PCOAN patients were 62.5/37.5% and 49.0/51.0%, respectively (Table III). Distributions of IRS1 Ala804Ala A/G allele in controls and PCOAN patients were 72.0/28.0% and 63.1/36.9%, respectively (Table III). Distributions of IGF1R IVS7-20 C/T allele in controls and PCOAN patients were 61.5/38.5% and 89.9/10.1%,
respectively (Table III). Distributions of IGF1R Thr766Thr C/T allele in controls and PCOAN patients were 63.0/37.0% and 55.8/44.2%, respectively (Table III).
Discussion
AN has been reported to be strongly influenced by ethnicity, and is common in children and adolescents, especially in populations with high rates of adult diabetes (26,27). However, two recent reports have challenged those describing AN as a reliable predictor of hyperinsulinaemia (28,29). Hirschler et al. reported the BMI of subjects with AN to be significantly greater than those of subjects without AN, while neither fasting immunoreactive insulin (IRI) nor the homeostasis model assessment insulin resistance index (HOMA-IR) differed in Hispanic subjects (30). In a Japanese study, there was a significant difference between the AN-positive and AN-negative groups in duration of obesity, in addition to age and percent overweight (31). Furthermore, we proposed a clinical observation of large proportion of pre- and co-obese AN patients that AN may occur before and concurrent the appearance of obesity, and investigated its possible mechanism via a pioneer genetic approach.
According to the results shown in Table 1, we have some explanations and annotations about the data presented and the stories behind them
(Table 1). In our study, both groups had similar gender distribution that prevents gender bios because female was reported to have higher peripheral insulin sensitivity than male (32). The PCOAN group was of younger age, higher serum fasting glucose and insulin level than the control group. The blunted insulin sensitivity in PCOAN group supported our hypothesis that PCOAN may stem from indigenous insulin/IGF receptor pathway genetic difference since insulin sensitivity tends to decline with age (33). In our cohort, 30 percent of PCOAN patients showed no insulin resistance, implying that other factors such as EGFR, FGFR and leptin should play a more important role in these non-insulin resistance patients (34). On the other hand, the use of fasting serum insulin level or HOMA-IR as indicators of insulin resistance may be inappropriate if glucose clamp test was viewed as a gold standard. However, it is difficult and impossible to perform clamp test in our obese children both in parents’ will and technique aspect (35). The PCOAN group was of higher HOMA-IR index than the control group, which meant this group was of higher insulin resistance. No consensus was agreed on the cut-off value of HOMA-IR of obese children, but a value higher than 3 was generally considered as insulin resistance (36,37).
Previous genetic studies about AN have focused largely on the -adrenergic receptor (B3ADR) which discuss the associations of AN with obesity, cardiovascular diseases, and type 2 diabetes (38–48). In this paper, we firstly investigated another pathway which is also closely related to obesity--- the insulin/IGF receptor pathway, and its genetic association with PCOAN. Among the four SNPs we investigated in this pioneer research, we found that variant genotypes of INSR His1085His and IGF1R IVS7-20 were significantly associated with a higher susceptibility of PCOAN (Tables II and III). The results suggested that both insulin and IGF receptor subpathways are involved in PCOAN, and the involvements of their downstream genes, together with the crosstalk between them are all worth further investigations.
From the viewpoint of molecule cellular level, insulin interacted with not only insulin receptor but IGF receptor, activating their downstream effectors and even crosstalking with each other (49). Along with afore-mentioned insulin resistance that might resulted in failure of suppression of excessive energy intake with ensuing obesity, these concept fits our results showing that there is a significant association of insulin/IGF
receptor pathway with PCOAN. It is well known that most obesity may originate from disturbed interaction between genetics and environment that could not be explained by a single factor, our study provided a specific genetic factor, insulin/IGF receptor pathway, played a certain part of this complex pathogenesis of obesity under the scrutiny of patients’ phenotype and genotype association.
In the future, further molecular studies are needed to elucidate the complex relationship between these polymorphisms and PCOAN. In addition, more genetic population studies about AN, like the association studies, can strengthen the power to elucidate the correlation among AN and other diseases, such as obesity and diabetes mellitus. Our preliminary data provide not only the evidence of the T allele of INSR His1085His and the C allele of IGF1R IVS7-20 are correlated with the appearance of AN precede or concurrence with obesity but revealed that insulin/IGF receptor pathway may play an important role in Taiwanese pre- and co-obese AN.
This study is supported by grants partly from the China Medical University Hospital (CMU-97-328). We specially appreciate every patient and colleague for their cooperation and contribution.
Conflicts of Interests
The authors declare that there is no conflict of interest.
References
1. Friedman JM. Obesity: cases and control of excess body fat. Nature. 2009;459:340–2.
2. Krebs NF, Himes JH, Jacobson D, Nicklas TA, Guilday P, Styne D. Assessment of child and adolescent overweight and obesity.
Pediatrics. 2007;120:S193–228.
3. Sturm R. The effects of obesity, smoking, and drinking in medical problems and costs. Health Aff. 2002;21:245–53.
4. Pomietto M, Docter AD, Van Borkulo N, Alfonsi L, Krieger J, Liu LL. Small steps to health: building, substainable partnerships in pediatric obesity. Pediatrics. 2009;123:S308–16.
5. Guran T, Turan S, Akcay T, Bereket A. Significance of acanthosis nigricans in childhood obesity. J Paediatr Child Health. 2008;44:338– 41.
6. Schwartz RA. Acanthosis nigricans. J. Am. Acad. Dermatol. 1994;31:1–19.
7. Schwartz RA. Acanthosis nigricans. In: Demis DJ, editor. Clinical dermatology (unit 12-26). 18th ed. Philadelphia: JB Lippincott; 1999; 1–11.
8. Tasjian D, Jarratt M. Familial acanthosis nigricans. Arch Dermatol. 1984;120:1351–4.
9. Skiljevic DS, Nikolic MM, Jakovljevic A, Dobrosavljevic DD.
2001;18:213–6.
10.Birns J, Badawi RA, Chase AR, Watson T. Acanthosis nigricans associated with acute myeloid leukemia. Eur J Int Med. 2004;15:473. 11.Higgins SP, Freemark M, Prose NS. Acanthosis nigricans: a practical
approach to evaluation and management. Dermatology Online Journal. 2008;14:2.
12.Barsh GS, Schwartz MW. Genetic approaches to studying energy balance: perception and integration. Nat Rev Genet. 2002;3(8):589-600.
13.Flier JS, Eastman RC, Minaker KL, et al. Acanthosis nigricans in obese women with hyperandrogenism. Characterization of an insulinresistant state distinct from the type A and B syndromes. Diabetes 1985;34:101-7.
14.Leme CE, Wajchenberg BL, Lerario AC, et al. Acanthosis nigricans, hirsutism, insulin resistance and insulin receptor defect. Clin
Endocrinol (Oxf) 1982;17:43–9.
15.Rique S, Nogues C, Ibanez L, et al. Identification of three novel mutations in the insulin receptor gene in type A insulin resistant patients. Clin Genet. 2000;57:67–9.
16.Moller DE, Cohen O, Yamaguchi Y, et al. Prevalence of mutations in the insulin receptor gene in subjects with features of the type A
syndrome of insulin resistance. Diabetes 1994;43:247–55. 17.Bau DT, Tseng HC, Wang CH, et al. Oral cancer and genetic
polymorphism of DNA double-strand break gene Ku70 in Taiwan. Oral Oncol. 2008;44:1047–51.
18.Chiu CF, Tsai MH, Tseng HC, et al. A novel single nucleotide polymorphism in XRCC4 gene is associated with oral cancer
susceptibility in Taiwanese patients. Oral Oncol. 2008;44:898–902. 19.Chiu CF, Tsai MH, Tseng HC, et al. A novel single nucleotide
polymorphism in ERCC6 gene is associated with oral cancer susceptibility in Taiwanese patients. Oral Oncol. 2008;44:582–6. 20.Chiu CF, Wang CH, Wang CL, et al. A novel single nucleotide
polymorphism in XRCC4 gene is associated with gastric cancer susceptibility in Taiwan. Ann Surg Oncol. 2008;15:514–8.
21.Bau DT, Wu HC, Chiu CF, et al. Association of XPD polymorphisms with prostate cancer in Taiwanese patients. Anticancer Res.
2008;27:2893–6.
polymorphism in XRCC4 gene is associated with breast cancer
susceptibility in Taiwanese patients. Anticancer Res. 2008;28:267–70. 23.Yang MD, Hsu YM, Kuo YS, et al. Significant Association of Ku80
Single Nucleotide Polymorphisms with Colorectal Cancer
Susceptibility in Central Taiwan. Anticancer Res. 2009;29:2239–42. 24.Chang CH, Chang CL, Tsai CW, et al. Significant Association of an
XRCC4 Single Nucleotide Polymorphism with Bladder Cancer Susceptibility in Taiwan. Anticancer Res. 2009;29(5):1777–82.
25.Hsu CF, Tseng HC, Chiu CF, et al. Association between DNA Double Strand Break Gene Ku80 Polymorphisms and Oral Cancer
Susceptibility in Taiwan. Oral Oncol.
doi:10.1016/j.oraloncology.2008.12.002, 2008.
26.American Diabetes Association. Type 2 diabetes in children and adolescents. Diabetes Care. 2000;23:381–9
27.Stuart CA, Gilkison CR, Smith MM, Bosma AM, Bruce K, Nagamani M. Acanthosis nigricans as a risk factor for noninsulin dependent diabetes mellitus. Clin Pediatr. 1998;37:73–9.
28.Hirschler V, Aranda C, Oneto A, Gonzalez C, Jadzinsky M. Is
acanthosis nigricans a marker of insulin resistance in obese children? Diabetes Care. 2002;25:2353.
29.Nguyen TT, Keil MF, Russell DL, Pathomvanich A, Uwaifo GI, Sebring NG, et al. Relation of acanthosis nigricans to
hyperinsulinemia and insulin sensitivity in overweight African American and white children. J Pediatr. 2001;138:474–480. 30.Hirschler V, Aranda C, Oneto A, Gonzalez C, Jadzinsky M. Is
acanthosis nigricans a marker of insulin resistance in obese children? Diabetes Care. 2002;25:2353.
31.Miura N, Ikezaki A, Iwama S, Matsuoka H, Ito K, Sugihara S. Genetic factors and clinical significance of acanthosis nigricans in obese
Japanese children and adolescents. Acta Pediatrica. 2006;95:170–5. 32.Borissova AM, Tankova T, Kirilov G, et al. Gender-dependent effect
of ageing on peripheral insulin action. Int J Clin Pract. 2005;59:422–6. 33.Petersen KF, Befroy D, Dufour S, et al. Mitochondrial dysfunction in
the elderly: possible role in insulin resistance. Science. 2003;300:1140–2.
34.Hermanns L, Hermanns JF, Pierard GE. Juvenile acanthosis nigricans and insulin resistance. Pediatr Dermatol. 2000; 19: 2–14.
method for quantifying insulin secretion and resistance Am J Physiol Gastrointest Liver Physiol 1979;237:G214–23.
36.Tresaco B, Bueno G, Pineda I, Moreno LA, Garagorri JM, Bueno M.
Homeostatic model assessment (HOMA) index cut-off values to identify the metabolic syndrome in children. J Physiol Biochem.
2005;61(2):381–8.
37.Madeira IR, Carvalho CN, Gazolla FM, de Matos HJ, Borges MA,
Bordallo MA. Cut-off point for Homeostatic Model Assessment for
Insulin Resistance (HOMA-IR) index established from Receiver Operating Characteristic (ROC) curve in the detection of metabolic syndrome in overweight pre-pubertal children. Arq Bras Endocrinol
Metabol. 2008;52(9):1466–73.
38.Walston J, Silver K, Bogardus C, Knowler WC, Celi FS, Austin S, et al. Time of onset of non-insulin-dependent diabetes mellitus and genetic variation in the beta 3-adrenergic-receptor gene. N Engl J Med. 1995;333:343–7.
39.Widen E, Lehto M, Kanninen T, Walston J, Shuldiner AR, Groop LC. Association of a polymorphism in the b3-adrenergic-receptor gene with features of the insulin resistance syndrome in Finns. N Eng J Med. 1995;333:348–51.
40.Clement K, Vaisse C, Manning BSJ, et al. Genetic variation in the b3-adrenergic receptor and an increased capacity to gain weight in
patients with morbid obesity. N Engl J Med. 1995;333:352–4. 41.Kadowaki H, Yasuda K, Iwamoto K, et al. A mutation in the
b3-adrenergic receptor gene is associated with obesity and
hyperinsulinemia in Japanese subjects. Biochem Biophys Res Comm. 1995;215:555–60.
42.Yoshida T, Sakane N, Umekawa T, Sakai M, Takanashi T, Kondo M. Mutation of b3-adrenergic-receptor gene and response to treatment of obesity. Lancet. 1995;346:1433–4.
43.Fujisawa T, Ikegami H, Yamato E, et al. Association of Trp64Arg mutation of the b3-adrenergic receptor with NIDDM and body weight gain. Diabetologia. 1996;39: 349–52.
44.Arner P. The b3-adrenergic-receptor a cause and cure of obesity. N Engl J Med. 1995;333:382–3.
45.Pulkkinen A, Kareinen A, Saarinen L, Heikkinen S, Lehto S, Laakso M. The codon 64 polymorphism of the beta3-adrenergic-receptor gene is not associated with coronary heart disease or insulin resistance in
nondiabetic subjects and noninsulin-dependent diabetic patients. Metabolism. 1999;48:853–6.
46.Nagase T, Aoki A, Yamamoto M, et al. Lack of association between the Trp64Arg mutation in the b3-adrenergic receptor gene and obesity in Japanese men: a longitudinal analysis. J Clin Endocrinol Metab. 1997;82:1284–7.
47.Large V, Hellstrom L, Reynisdottir S, et al. Human beta-2
adrenoceptor gene polymorphisms are highly frequent in obesity and associate with altered adipocyte beta-2 adrenoceptor function. J Clin Invest. 1997;100:3005–13.
48.Ishiyama-Shigemoto S, Yamada K, Yuan X, Ichikawa F, Nonaka K. Association of polymorphisms in the b2-adrenergic receptor gene with obesity, hypertriglyceridaemia, and diabetes mellitus. Diabetologia. 1999;42:98–101.
49.Cullen MT, Brice E, Kahn CR. Critical nodes in signalling pathways: insights into insulin action. Nature Reviews Molecular Cell Biology. 2006;7:85–96.
Figure legend
Postulated mechanisms for the pathogenesis of pre and co-obese acanthosis nigricans (Modified from Steven P Higgins, Michael Freemark, Neil S Prose. Acanthosis nigricans: A practical approach to evaluation and management. Dermatology Online Journal 14 (9): 2, 2008).
Table I. The clinical and biochemical features of pre-obesity and co-obesity AN
patients (PCOAN) and control groups
PCOAN group Control group P
Age 10.8 (4-18) 24 (19-40) < 0.0001 Sex (male/female) 55/44 50/50 0.435 BMI (kg/m2) 30.7 (22.4-44.2) 20.1 (18-22) < 0.0001 Fasting glucose (mg/dL) 101.1 (77-327) 81.5 (64-96) < 0.0001 Fasting insulin (U/mL) 21.1 (3.9-77.9) 10.3 (6.3-12.4) < 0.0001 HOMA-IR 5.4 (3.7-21.3) 1.8 (0.8-2.6) < 0.0001
Table II. Distributions of IRS1, INSR and IGF1R genotypes among pre-obesity and
co-obesity AN (PCOAN) patients and control groups.
Genotype Controls % Patients % Pa
INSR His1085His (rs1799817) 0.0462 CC 36 36.0% 18 18.2% CT 53 53.0% 61 61.6% TT 11 11.0% 20 20.2% IRS1 Ala804Ala (rs1801123) 0.2516 AA 53 53.0% 39 39.4% AG 38 38.0% 47 47.5% GG 9 9.0% 13 13.1% IGF1R IVS7-20 (rs2272037) 8.77E-6 CC 36 36.0% 80 80.8% CT 51 51.0% 18 18.2% TT 13 13.0% 1 1.0% IGF1R Thr766Thr (rs2272037) 0.4253 CC 43 43.0% 28 32.6% CT 40 40.0% 40 46.5% TT 17 17.0% 18 20.9%
Table III. Distributions of IRS1, INSR and IGF1R alleles among pre-obesity and co-obesity AN (PCOAN) patients and control groups.
Allele Controls % Patients % Pa
INSR His1085His (rs1799817) 0.0255 Allele C 125 62.5 % 97 49.0 % Allele T 75 37.5% 101 51.0% IRS1 Ala804Ala (rs1801123) 0.1025 Allele A 144 72.0% 125 63.1% Allele G 56 28.0% 73 36.9% IGF1R IVS7-20 (rs2272037) 2.84E-6 Allele C 123 61.5% 178 89.9% Allele T 77 38.5% 20 10.1% IGF1R Thr766Thr (rs2272037) 0.2043 Allele C 126 63.0 % 96 55.8% Allele T 74 37.0% 76 44.2%