棲蘭檜木林不同林地類型之野生動物群聚
陳一銘1,2) 何紋靈1) 翁瑞鴻1) 葛兆年1)
摘 要
棲蘭檜木林是台灣代表性的山地雲霧林之一,在生物多樣性保育中具有重要的地位,然而有關其 野生動物群聚的研究仍然不足。我們於2010~2013年間在棲蘭檜木林區選擇原始林(無人為干擾)、整 理林 (枯倒木移除)與人工林(皆伐後造林或天然更新)等三種林地類型進行動物豐量之比較研究,以自 動相機進行調查。總共完成310次獨立之自動相機取樣,計10,231工作天,攝得2710筆哺乳類影像資 料,含山羌(Muntiacus reevesi)、台灣野山羊(Capricornis swinhoe)、台灣獼猴(Macaca cyclopis)、鼬 獾(Melogale moschata)、野豬(Sus scrofa)、食蟹 (Herpestes urva)、黃鼠狼(Mustela sibirica)、黃喉 貂(Martes flavigula)、白面鼯鼠(Petaurista alborufus)、赤腹松鼠(Callosciurus erythraeus)、長吻松鼠 (Dremomys pernyi)與條紋松鼠(Tamiops maritimus)等12種以及無法辨識的地棲性鼠科(Muridae)動物,
其中食蟹 與黃喉貂為本地區新紀錄。最優勢的三種動物為山羌、野山羊與台灣獼猴,共佔所有動物 影像的 93.1%,其相對組成在三種林地類型間差異顯著。比較各物種的出現頻度(OI值,影像數/1000 h),只有野山羊的OI值在三種林地類型間具有顯著差異,而以人工林顯著高於原始林。在同樣的林地 類型中,山羌的 OI值數據變異較大,於原始林的樣區間、人工林的樣區間都有極顯著差異;野山羊的 OI值數據變異小,無論在何種林地類型的樣區間皆無顯著差異,推測可能與野山羊的攀爬能力較強有 關。主成分分析 (PCA)因素負荷量圖顯示原始林與整理林區隔不明顯,而人工林樣區則彼此較接近。
我們認為整理林歷經長期復原,目前動物群聚與原始林差異不大,而皆伐後的更新造林地所受的影響 則比較長遠。
關鍵詞:棲蘭山、檜木林、枯倒木整理、野生動物群聚、自動相機。
陳一銘、何紋靈、翁瑞鴻、葛兆年。 2015。棲蘭檜木林不同林地類型之野生動物群聚。台灣林業科學 30(1):75-88。
1)林 業 試 驗 所 森 林 保 護 組 ,10066台北市南海路53號 Division of Forest Protection, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
2)通訊作者 Corresponding author, e-mail:Yimingo@yahoo.com.tw
2014年8月送審 2015年1月通過 Received August 2014, Accepted January 2015.
Research paper
Mammal Assemblages of Different Forest Types in the Cilan Cypress Forest
Yi-Ming Chen,1,2) Wun-Ling He,1) Ruei-Hong Wong,1) Chao-Nien Kuo1)
【Summary】
The Cypress Forests (Chamaecyparis formosensis and C. obtusa var. formosana) at Cilan are a symbolic mountain cloud forest ecosystem in Taiwan, and play an important role in biodiversity conservation; however studies of mammal assemblages of the area are still insufficient. We con- ducted a comparative study on mammal abundances in the Cilan area during 2010~2013. Three forest types were selected: virgin forests (undisturbed), managed forests (from which snags and fallen logs had been removed), and plantations (reforested after having been clear-cut). The cam- era-trapping method was applied at 4 different sample sites in each forest type. By establishing 310 independent camera-trapping events, with an effort of 10,231 camera-days, 12 species of mammals were photo-captured as follows: Reeve’s Muntjac (Muntiacus reevesi), Formosan Serow (Capri- cornis swinhoe), Formosan Macaque (Macaca cyclopis), Ferret Badger (Melogale moschata), Wild Boar (Sus scrofa), Crab-eating Mongoose (Herpestis urva), Siberian Weasel (Mustila sibirica), Yel- low-throated Marten (Martes flavigula), White-faced Flying-squirrel (Pitaurista alborufus), Red- bellied Squirrel (Callosciurus erythraeus), Long-nosed Squirrel (Dremomys pernyi), and Striped Squirrel (Tamiops maritimus), in addition to unidentified members of the Muridae. Records of the Crab-eating Mongoose and Yellow-throated Marten are new to the Cilan area. The top 3 most recorded species were Reeve’s Muntjac, Formosan Serow, and Formosan Macaque, comprising a major portion (91.3%) of the captured images. Relative compositions of these 3 dominant mam- mals significantly differed among the 3 forest types. Based on the occurrence index (OI, number of images/1000 camera-hours) of the animals, only the Formosan Serow showed distinct differences among the 3 forest types. Its OI value was higher in plantations than in virgin forests. The OI data set of Reeve’s Muntjac showed more variation within the same forest type, with distinct differ- ences among all sample sites in both of the virgin and plantation forests. On the other hand, the OI of the Formosan Serow was less variable, which might have resulted from their superior climbing ability. A principle component analysis (PCA) plot revealed that there was no segregation between the virgin forest sites and managed forest sites, while the 4 plantation sites were closer. We con- cluded that there were no obvious differences in the mammal assemblages between virgin forests and managed forests, which have undergone succession for several decades. However, plantations regenerated from clear-cut stands exhibited more long-term influences.
Key words: Cilan, cypress forest, cloud forest, wildlife assemblage, photo-trapping.
Chen YM, He WL, Wong RH, Kuo CN. 2015. Mammal assemblages of different forest types in the
Cilan cypress forest. Taiwan J For Sci 30(1):75-88.
緒 言
棲 蘭 林 區 位 處 山 地 雲 霧 盛 行 帶 , 氣 候 穩 定而重濕,因而發展出以台灣特有的檜木為優 勢,面積最大、最具代表性的山地雲霧林,區 內紅檜 (Chamaecyparis formosensis)和台灣扁 柏(Chamaecyparis obtusa var. formosana)的蓄 積 量 相 當 大 , 是 非 常 重 要 的 天 然 資 源 (Chang 1963, Kuo and Chan 1993, Wang 2000)。台灣 的雲霧林具有相當高的生物特殊性,Chen and Mao (2004)引述相關報告(Wang 2000)表示,霧 林帶的種子植物特有率明顯高於台灣全區之植 物特有率,故而霧林帶於台灣具有應予保護之 價值,顯見其在保育之重要性。目前本區已劃 歸棲蘭重要野生動物棲息環境之一部分,並包 含鴛鴦湖自然保留區在內,因此棲蘭山檜木林 無疑也是極為重要的野生動物棲息環境,過去 本地區與週邊曾進行過野生動物相關之調查(LL Lee 2000, JH Lee 2002, Chen and Mao 2004, 2007)已為本區野生哺乳動物的組成描述建立基 礎,然而歷次研究成果間仍有差異,同時有關 野生動物相對豐量組成與比較等量化之研究仍 然不足。其中本研究所在的森林保育處棲蘭工 作站100線林道後段與160、170林道被認為是棲 蘭、北橫地區中、大型野生動物多樣性較高的 地點(Lee 2000)。
除 了 早 期 的 伐 木 , 本 林 區 分 別 於 1 9 8 6 ~ 1993、1994~1999年分別進行過二期的枯立倒 木整理作業,自檜木老齡林中以架空集材方式 搬出枯死、風倒木標售利用,並於整理區內林 木稀疏之處實施橫坡帶狀整地、中耕培土,以 自生苗保留、天然下種及人工栽植等方式進行 更新(Lin 1999)。然而台灣已於1993年宣告全面 停伐天然林,基於前述雲霧林的特殊性與保育 重要性,致第二期的枯立倒木整理作業乃引發 森林經營與自然保育衝突的論戰(Chen 1999)。
棲 蘭 檜 木 林 枯 立 倒 木 整 理 作 業 已 於 1999年中 止,然其衍生有關保育生物學的議題仍然值得 探討驗證。
森林因人為撫育程度的強弱、年代長短,
可能改變森林棲地結構的複雜性以及植物物種 的多樣性,這些都可能是影響動物群聚的因子
(Chey et al. 1997, Yang 2003)。伐木集材期間 林下植被結構的破壞,是影響鳥類及哺乳類群 聚的主因,但隨著林下植被的回復,撫育作業 對 鳥 類 及 哺 乳 類 群 聚 的 影 響 始 也 會 減 緩 (Tsai 2001, Hsieh 2003)。若是對林地行強度伐採等干 擾,則當地植被的大量破壞會對某些依賴森林 的鳥種族群造成長時間的負面影響(Sekercioglu 2002)。較大型的野生動物常被選為可代表環境 棲地完整的護傘種(umbrella species)或旗艦種 (flagship species, Caro and O’Doherty 1999),由 於也是重要的狩獵物(game animal),在保育上也 較受到重視,然而營林作業對較大型野生動物 之影響在台灣很少有相關研究成果。因此本研 究即希望能透過檢測棲蘭林區較大型野生動物 的種類、相對豐量、群聚結構等,來了解棲蘭 山檜木林地在不同的經營模式下作為野生動物 棲地的功能如何?尤其在經過長時間復原的原 始檜木林與檜木整理林之間其差異是否存在?
相較於小型而相對較普遍的物種,傳統上 大型野生動物的調查有其技術上的困難之處(Lin et al. 1998)。然而近年來由於紅外線感應照相 設備的長足發展,使得研究人員得以突破過去 陸生哺乳動物調查資料獲取的困難(Pei 1998),
並且讓資料更客觀、更容易量化。此外棲蘭 100 線等林道由於交通可及性較高、干擾相對較少 等優勢,也比較適合進行努力量需求較高的研 究工作。其中Chen and Mao (2007)曾於棲蘭160 林道之檜木林針對相鄰的枯倒木整理區與非整 理區,除了對植群特性加以描述分類外,也以 自動照相機進行野生動物豐富度的比較,發現 兩區差異不顯著,但在小型哺乳動物捕捉則以 整理林顯著優於原始林。不過此研究之自動相 機調查部分僅採用物種整體出現頻度,且樣區 侷限於棲蘭160林道6 km附近(本研究R2樣區)。
另外Yuan et al. (2004)也從鳥類群聚的特性來探
討160林道沿線這兩種檜木林棲地中鳥類功能群
(guild)的差異。我們除擴大研究範圍,增加取
樣努力以求更仔細的描述棲蘭林區的較大型動
物群聚組成特性外,並將100線林道14 km以後
的的柳杉及檜木造林地一併納入調查範圍,希
望藉拉大生態跨幅來增加解讀資料的可能性,
獲得更具體的結論。
材 料 與 方 法
一、樣區環境概述
實驗地範圍位於棲蘭100線林道14~28 km,
接續170、160兩條支線林道,行政區域跨宜蘭、
新竹兩縣,屬雪山山脈向東北延伸之主稜,為 蘭陽溪與淡水河系的主要分水嶺,平均高度約 為海拔1700~2200 m之間。本地區除了因位處 霧林帶而有豐沛而均勻的降雨外,同時也是東 北季風的迎風帶,平均年雨量達 3477 mm,均 溫為11~15℃ (Chen and Mao 2004)。本地的原 生植群以檜木林為主,一般稜線與上坡位以針 葉樹為優勢,溪谷與下坡位則多針闊葉混生林 (Chen and Mao 2007)。今100線林道沿線多為皆 伐作業後植造或天然下種、補植形成的柳杉或檜 木人工林;而 160與170林道沿線則有較多的枯 倒木整理林以及小部分殘存的原始檜木林,林 道盡頭之後則大致為未經干擾的原始檜木林。
二、樣區配置
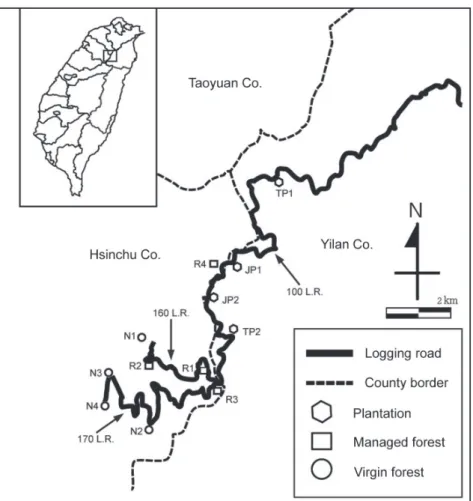
為求通盤了解棲蘭林區的地棲野生動物群 聚,並比較過去不同的經營作業對野生動物群 聚造成的影響,本研究將試驗地依照人為干擾 (營林撫育作業)之程度區分為少有干擾的原始檜 木老齡林(原始林)、曾經過枯倒木整理作業的 老齡檜木林(整理林)、檜木及柳杉造林地(人工 林)等3種林地類型。由於大部分整理林改良作 業已歷經多年演替,從外觀上並不易與原始林 區分,我們乃依據森林保育處提供的台帳配合 1/5000相片基本圖進行各種林地類型劃界,並 從航照影像大略判讀植被完整性以及等高線判 識地形來規劃可能的樣區,最後經現場踏勘來 決定樣區配置,並於樣區內劃設樣線。結果共 選出原始林樣區4處、整理林樣區4處、人工林 樣區 4處(Fig. 1, Table 1),茲分述如下:
(一) 原始林樣區
以160林道終端(N1)、170林道5 km開闊
山谷(N2)、170林道9 km蕃社跡登山小徑(N3) 與 170林道終端(N4)等4處為原始檜木林調查樣 區,上述樣區的地形與植被特性都不相同,詳 見 Table 1。原始林大致人為干擾很小,其中 N1、N2在調查樣線的起始處包含一小段廢棄林 道, N2、N4樣區則曾發現盜伐殘跡。
(二) 整理林樣區
以 160林道1.5 km (R1)、160林道6 km「檜 木的故鄉」(R2)、170林道起點(R3)與22.6 km 小支稜 (R4)為整理林樣區。其中R1、R2和R3分 別在1987、1989與1984年度進行枯立倒木整理 作業。目前樹冠層組成仍為當初保留的巨大檜 木,高出與次層樹冠甚多;而天然下種及人為更 新後的次生檜木明顯比原始林多,生長整齊。
(三) 人工林樣區
以 100線林道主線14 km (TP1)及25.5 km (TP2)的檜木人工林,與21 km (JP1)及24 km (JP2)的柳杉人工林作為人工林樣區,這兩種人 工林在外觀結構上並無明顯不同。由於林木樹 齡相近、樹高及胸徑差異不大,且植株間距整 齊均一,因此人工林樹冠層綿延相接,林下較 陰暗而沒有明顯的次冠層與灌木層,地被覆滿 蘚苔類植物。有時林地有殘枝、倒木橫陳,可 能是過去原木伐採或疏伐作業所致。
三、樣線設置與自動相機取樣
本研究以紅外線自動感應相機進行野生動 物影像捕獲法(photo-trapping, camera trapping) 調查,以了解棲息的野生動物種類並評估其相 對豐量 (relative abundance)。動物影像捕獲法 假設拍攝到某種類之影像次數越多,即代表該 種 類 野 生 動 物 之 相 對 豐 量 越 高 , 由 於 可 以 在 野外自動工作,而廣為野生動物研究採用(Pei 1998)。調查期間為2010年9月至2013年8月,
方式為沿著樣線進行取樣,樣線設於樣區森林 內,以開路或利用現有山徑往大略方向延伸,
除通行的便利性以外,也要避免過度偏好相同
的環境條件。由於每個樣區的範圍大小與地形
條件相異,因此視野外操作的可行性,各樣區
內設置的樣線數量與配置並不相同。設置樣線 時 每 20 m標記為一個樣段,作為相機取樣單 元,最終各樣區分別規劃成 30~36個樣段不等 (Table 1)。
相機架設樣點限定於特定樣段左右10 m範 圍內,選擇野生動物可能出入頻繁之處架設,調 查期間每月前往試驗地回收前月設置之相機(若 遇颱風或道路坍方,則順延至隔月),並以至少 間隔1個樣段的方式循序輪替設置新相機進行取 樣,調查期間各樣段僅取樣1次,以兼顧取樣的 方便並維持樣本之獨立性。此外由於樣線延伸必 然會經過較多的微棲地類型,因此也可以降低 主觀的取樣偏差。取回之相機除讀取資料及保 養外,並以手動測試其自動拍攝功能,確保相 機作用正常以供下次調查更換之用。原則上每
個樣區以相機3部進行取樣,但由於相機普遍有 故障情形,因此並非每個樣點都能成功獲取樣 本資料,最終每個樣區都完成至少1年的調查並 獲取至少 18份樣本資料以進行進一步的分析。
本研究採用數位式紅外線感應相機 (Scout guard SG550,廠牌Boly-Media),相機設定參 數主要為:啟動時間(trigger time)約1.2 s;感應 靈敏度(sensitivity)設為中等;拍攝方式為每次 錄影4~6 s;每一次錄影事件之延遲時間(delay) 為60 s。錄影在動物種類辨識上相當有利,同時 也可觀察到部分的行為表現,而設定延遲時間 則可以有效減少空拍量(例如枝葉搖晃)。相機架 設高度通常在 1.2 m以下,視現場情況設為不同 的俯仰角,有效的偵測中心距離通常在1~3 m之 間,以避免過大或過小的拍攝範圍。
Fig. 1. Road system, research sites, and the location of Cilan.
四、資料處理與分析
影 像 資 料 皆 於 相 機 攜 回 實 驗 室 後 進 行 判 讀,辨識紀錄動物種類、出現時間等資料。雖 然 台 灣 自 動 相 機 調 查 慣 常 將 1小時內出現無法 辨視之同種個體合併為同一筆錄(Chiang et al.
2007),但由於本研究的相機設定點絕大部分為 獸徑,根據判圖的經驗,同一隻動物個體重複 出現的情形通常為連續記錄,若非如此,則並 沒有明確的理由可以認定1小時內重複出現的紀 錄缺乏獨立性,因此本研究採直接判定的方式 處理類似情形。根據整體影像頻度判斷後,我 們採間隔小於10 min之紀錄才進行篩選,根據 影像中動物的行為及特徵判視「可能」為同樣 個體者登記為同一筆記錄(有效影像),否則即視 為不同個體之紀錄。本文以有效影像所占百分 比來描述動物群聚組成之特性,以卡方獨立性 檢驗測試前 3優勢種的有效影像數來比較林地類 型間之差異。在相對豐量的比較上,則以影像 的出現頻度 (單位時間出現量)來進行標準化,
目 前 台 灣 慣 用 出 現 頻 度 簡 稱OI值(occurrence index),其定義是以1000 h為單位時間,亦即:
OI = 特定動物影像出現數/相機工作小時×1000 (Pei 1997)。本研究用每個樣點同一部相機連 續時間的拍攝結果單獨計算各種動物的出現OI 值,並視為獨立的取樣。單一樣點的相機收回 後發現因故障或電力不足致使拍攝天數不足 10 日者排除不列入出現頻度的分析 (Chiang et al.
2007)。由於野生動物的行為特性與影像捕獲法 之取樣性質,各種動物在單一採樣點的出現頻 度變相當大,尤其數量較稀少的動物出現率為 零的情形很多,通常不服從常態分配 (Pei and Chiang 2004)。在經驗中認為這與野生動物傾 向使用固定路徑的習性有關,會因此造成相同 個體重複出現(Chen 2007)。本研究以樣區為重 複探討不同林地類型的動物出現頻度差異性,
基於前述考量,將各樣區所有樣點的 OI值樣本 求取平均後作為觀測值以避免數據偏態,並採 單因子變異數分析 (ANOVA)檢定後再以Tukey’s Table 1. Environmental traits related to all of the sites of the 3 forest types at Cilan
Forest type Site Locality Terrain Dominant woody plants No. effort Sampling (day) Virgin N1 160 R.
2)end Slope, ridge Chamaecyparis
1), Schima superba, 23 701
Rhododendron formosanum.
N2 170 R. 5 km Valley Chamaecyparis, Pinus taiwanensis 20 697 N3 170 R. 9 km Prim. ridge Chamaecyparis, Rhododendron 23 705
formosanum.
N4 170 R. end Abandoned R. Chamaecyparis, Cunninghamia 23 691 konishii
Managed R1 160 R. 1.5 km Sec. ridge, slope Chamaecyparis 18 510 R2 160 R. 6 km Mild slope Chamaecyparis 19 566 R3 170 R. 0.5 km Mild slope Chamaecyparis 29 950 R4 100 R. 22.6 km Ridge Chamaecyparis, Rhododendron 30 1749
formosanum
Plantation TP1 100 R. 14 km Mild slope Chamaecyparis 34 990 TP2 100 R. 25.5 km Mild slope Chamaecyparis 25 759 JP1 100 R. 21 km Mild slope Cryptomeria japonica 36 1026 JP2 100 R. 24 km Mild slope Cryptomeria japonica 30 887
Sum 310 10,231
1)
The genus Chamaecyparis includes C. obtusa var. formosana and C. formosensis.
2)
R., logging road.
honest significant difference (HSD)進行事後檢 定 ; 若 物 種 在 OI值合併平均後仍有多數零值 時,則仍採無母數Kruskal-Wallis變異數分析,
事後成對比較則以Mann-Whitney分析進行成對 比較,並以組數為分母調降顯著水準進行校正 (Bonferroni corrections)。若是對特定林地類型 中不同之樣區進行差異比較,則以採樣點為重 複,故此時採用無母數 Kruskal-Wallis變異數分 析。此外為了解各樣區間動物群聚的特性與彼 此相關情形,排除僅有一次出現紀錄的黃喉貂 之後,以物種的出現頻度進行主成份分析 (PCA) 運算,並繪製因素負荷量圖。以上分析工作皆 以統計程式R-3.1.1 (R Core Team 2014)運算 處理。
結 果
一、棲蘭山林區哺乳動物群聚組成
本研究總共回收 409台次自動相機,扣除 99次相機故障、設置失敗、取樣天數不足10日 或被偷,總共得到310個獨立之自動相機取樣,
計10,231工作天(Table 1)。攝得之影像扣除重 複與無法辨識者,共得到2934筆有效之影像,
其中2712筆為哺乳類、222筆為鳥類,平均每 部相機每天可攝得 0.29筆有內容之影像。本調 查記錄哺乳動物有:山羌(Muntiacus reevesi)、
台灣野山羊 (Capricornis swinhoe)、台灣獼猴 (Macaca cyclopis)、鼬獾(Melogale moschata)、
野豬 (Sus scrofa)、食蟹 (Herpestis urva)、黃 鼠狼(Mustela sibirica)、白面鼯鼠(Petaurista alborufus)、黃喉貂(Martes flavigula)、赤 腹 松 鼠 (Callosciurus erythraeus)、長吻松 鼠 (Dremomys pernyi)與條紋松鼠(Tamiops maritimus)等12種以及無法辨識的地棲性鼠科 (Muridae)動物。其中以山羌的有效影像占所有 影像的73.1%為最高,野山羊占11.8%、獼猴占 8.2%次之,其他動物影像所占的百分比都很低 (Fig. 2)。除齧齒動物外各林地類型出現的物種 數為原始林 6種、整理林8種、人工林6種,其 中人工林未記錄到野豬、黃喉貂,原始林則未 記錄到食蟹 (Table 2)。主要物種在各林地類
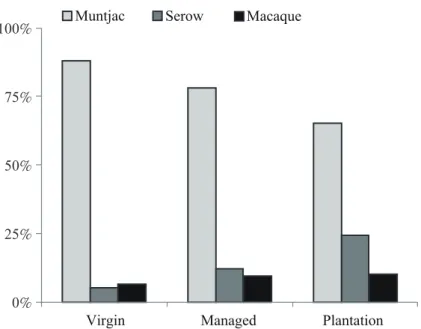
型中的百分比組成各有不同,3種林地類型都以 山羌的影像所佔百分比最高。在整理林以及人 工林,野山羊的比例高於獼猴,但是在原始林 台灣獼猴的比例卻高於野山羊。此外山羌所占 比例依原始林、整理林、人工林的順序遞減,
但在野山羊則反呈遞增的趨勢(Fig. 3)。選取山 羌、野山羊、獼猴等3種優勢動物之有效影像進 行卡方獨立性檢驗,結果顯示這3種動物的相對 組成在3種林地類型間具有極顯著的差異(Chi- squared = 151.0, 4 d.f., p < 0.001)。
二、不同林地類型間的比較
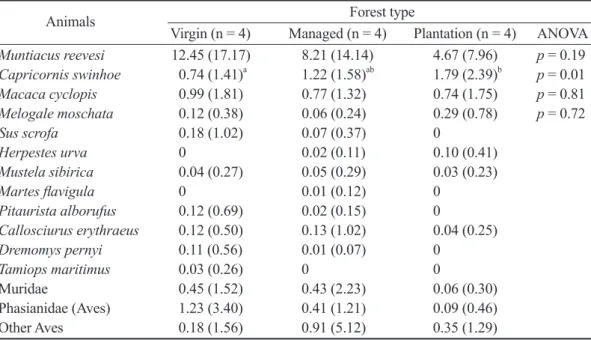
3種林地類型中物種出現頻度(OI值)分布情 形如 Table 2。進一步對出現頻度較高的較大型 物種分別進行單因子變異數分析,其中野山羊 的出現頻度由高至低分別為人工林、整理林以 及 原 始 林 , 且 在 3種林地類型之間具有顯著差 異,Tukey’s HSD事後檢定顯示其中以原始林與 人工林之間具有顯著差異。山羌的出現頻度高 低與野山羊相反,由高至低則依序為原始林、
整理林以及人工林,然而單因子變異數分析結 果3種林地類型間的差異不顯著。台灣獼猴、鼬 獾的出現頻度在 3種林地類型間皆無顯著差異。
台灣野豬在人工林中完全沒有出現紀錄,而在 其他兩種林地類型中出現頻度也不高,分析結 果同樣沒有顯著差異(Kruskal-Wallis H = 0.53, 1 d.f., p = 0.57);食蟹 、黃鼠狼及黃喉貂則因出 現樣點太少而未進行分析。以簡單相關分析配 對檢定物種間的出現頻度相關情形,發現山羌 與山豬之間存有極顯著的中度正相關(r
2= 0.52, 10 d.f., p = 0.008),其他各物種間均無相關性。
三、相同林地類型之樣區間的比較
以 OI值較高的物種進行無母數Kruskal- Wallis變異數分析,結果如Table 3。山羌在原 始林的4樣區間出現頻度有極顯著差異;人工林 的4樣區間出現頻度也有極顯著差異;整理林的 4樣區間則無差異。台灣獼猴的出現頻度,在 整理林的4樣區間、人工林的4樣區間都有顯著 差異,在原始林地中則沒有差異。關於鼬獾,
只 對 原 始 林 與 人 工 林 兩 種 類 型 進 行 檢 定 , 結
Table 2. The average occurrence index (OI)
1)of photo-trapped animals in 3 forest types at Cilan during 2010~2013
Animals Forest type
Virgin (n = 4) Managed (n = 4) Plantation (n = 4) ANOVA Muntiacus reevesi 12.45 (17.17) 8.21 (14.14) 4.67 (7.96) p = 0.19 Capricornis swinhoe 0.74 (1.41)
a1.22 (1.58)
ab1.79 (2.39)
bp = 0.01 Macaca cyclopis 0.99 (1.81) 0.77 (1.32) 0.74 (1.75) p = 0.81 Melogale moschata 0.12 (0.38) 0.06 (0.24) 0.29 (0.78) p = 0.72
Sus scrofa 0.18 (1.02) 0.07 (0.37) 0
Herpestes urva 0 0.02 (0.11) 0.10 (0.41)
Mustela sibirica 0.04 (0.27) 0.05 (0.29) 0.03 (0.23)
Martes flavigula 0 0.01 (0.12) 0
Pitaurista alborufus 0.12 (0.69) 0.02 (0.15) 0
Callosciurus erythraeus 0.12 (0.50) 0.13 (1.02) 0.04 (0.25)
Dremomys pernyi 0.11 (0.56) 0.01 (0.07) 0
Tamiops maritimus 0.03 (0.26) 0 0
Muridae 0.45 (1.52) 0.43 (2.23) 0.06 (0.30)
Phasianidae (Aves) 1.23 (3.40) 0.41 (1.21) 0.09 (0.46)
Other Aves 0.18 (1.56) 0.91 (5.12) 0.35 (1.29)
1)
The OI value is the number of images/1000 working hours. Values are presented as the mean and (standard deviation).
2)
Values in a row with the same superscript letter indicate no significant difference at the 5% signifi- cance level as determined by a post-hoc Tukey’s HSD test.
Fig. 2. Proportions of mammal images captured in Cilan.
果均無顯著差異。至於野山羊,無論在何種林 地類型,樣區間之出現頻度較為均勻而無顯著 差異。
四、PCA因素負荷量繪圖
排除僅有單次出現紀錄之黃喉貂,以 7種中
大型物種於各樣區內的出現頻度為介量進行主
成份分析(PCA)。其前3軸之特徵值大於1,共 可解釋70.8%之變異量。其中PC1及PC2可解釋 54.6%之變異量,所繪製之PCA因素負荷量散布 圖可以看出人工林的4個樣區分布於右下方,以 較多的野山羊、鼬獾為特徵。整理林樣區與天 然林樣區則相當分散,其中整理林樣區R4與天 然林樣區 N3較偏向右下側,從樣區特性來看,
R4樣區位於22.6 km地理位置最接近造林地樣區 (Fig. 1),而N3樣區則為稜線上的登山小徑,地 形較崎嶇,此與其他兩個天然林樣區不同。從 整體散布型態判讀,我們認為原始林與整理林 的動物組成並沒有明顯的區隔(Fig. 4)。
討 論
過 去 棲 蘭 林 區 曾 經 執 行 過 動 物 相 相 關 研 究計畫 (LL Lee 2000, JH Lee 2002, Chen and Mao 2007)的調查的範圍不一,有些含括司馬
庫斯、北橫沿線與松蘿湖等地,根據文獻內容 篩選出棲蘭林區之動物紀錄資料後加以比較,
可以發現在前人研究中有 4種動物沒有出現在我 們的調查成果,分別為水鹿、穿山甲、白鼻心 以及台灣黑熊;而本調查則有食蟹 以及黃喉 貂沒有出現在前人調查成果中。這樣的差異,
除了因為某些動物的稀有性所致,與調查方法 也有很大的關係。例如水鹿在我們的 JP2樣區 (24Km)中曾經有排遺出現,不過並未被相機拍 到。從文獻中我們可以知道水鹿在雪山山系的 數量一直都不高(Wang and Wang 1993, Huang 2004, Lee 2007, Su 2008, Tsai and Pei 2008, Lin 2009a, b),猜測目前在棲蘭林區的數量可能相 當低。穿山甲主要分布於低海拔(Chao 1989),
我們過去於北部山區對穿山甲的洞穴普查發現 以海拔800 m以下出現較多,最高紀錄為羅培山 海拔約 1400 m左右(unpublished data),而棲蘭 林區穿山甲紀錄大約在海拔 1700 m,應屬偶然 Table 3. The average occurrence index (OI)
1)of 4 photo-trapped animals from sites in different forest types at Cilan during 2010~2013
Forest Type site Muntjac Serow Macaque Ferret Badger
Virgin N1 9.51 (12.83)
a2)0.27 (0.52) 0.86 (1.16) 0.05 (0.21) N2 14.87 (18.64)
ab0.93 (1.20) 0.35 (0.57) 0 N3 5.38 (4.32)
a0.99 (1.40) 0.87 (1.48) 0.22 (0.39) N4 20.37 (12.78)
b0.80 (0.88) 1.79 (1.94) 0.19 (0.34)
K-W ANOVA p = 0.002 p = 0.198 P = 0.159 p = 0.180
Managed R1 15.83 (16.36) 1.77 (1.26) 0.79 (1.04)
b0
R2 5.38 (4.85) 1.39 (1.63) 0.14 (0.32)
a0
R3 5.94 (4.85) 1.11 (1.22) 0.36 (0.53)
a0
R4 7.60 (8.53) 0.90 (1.37) 1.55 (2.60)
b0.18 (0.50)
K-W ANOVA p = 0.283 p = 0.555 p < 0.001 ---
Plantation TP1 0.18 (0.41)
a2.08 (1.42) 0.12 (0.29)
a0.41 (0.51) TP2 9.09 (8.23)
b1.86 (1.69) 1.16 (1.47)
b0.52 (0.86) JP1 3.49 (2.83)
b1.20 (1.06) 0.61 (1.25)
ab0.19 (0.49) JP2 7.49 (10.94)
b2.01 (3.31) 1.22 (1.99)
ab0.06 (0.25)
K-W ANOVA p < 0.001 p = 0.403 p = 0.014 p = 0.090
1)
The OI value is the number of images/1000 working hours. Values are presented as the mean and (standard deviation).
2)
Values with the same superscript letter in a column indicate no significant difference at the 0.8% (5%,
6 paired comparisons) significance level, as determined by the post-hoc Mann-Whitney U-test, after
a Kruskal-Wallis (K-W) ANOVA.
Fig. 3. Proportions of 3 dominant mammals’ images from different forest types in Cilan.
Fig. 4. The principle component analysis (PCA) factor loading plot of all sites in Cilan based on the occurance index (OI) of 7 mammal species.
Virgin forest: N1~N4 = 1~4; Managed forest: R1~R4 = 5~8; Plantation: Tp1 = 9; Tp2 = 10;
Jp1 = 11; Jp2=12.
Mj, Muntjac; Wb, Wild Boar; Mc, Macaque; Ws, Weasel; Bg, Ferret Badger; Sr, Serow;
Mg, Crab-eating Mongoose.
出現的個體。白鼻心的海拔分布可達海拔 2000 m (Cheng and Wang 1993),從前人的調查結 果來看,本種在棲蘭應屬不普遍,本研究未能 偵測到白鼻心,可能是樣區配置還不夠均勻之 故。台灣黑熊在雪山以北山區的數量稀少(Wang and Wang 1993, Huang 2012),因此並不容易以 自動相機調查,反而以其爪痕可存留數年較易 於偵測 (Huang 2012),這也是2002年(Lee 2002) 調查中棲蘭山林區黑熊紀錄的型態。
在前人研究並未記錄的物種中,食蟹 在 本研究中出現於 R4、JP1及JP2等3個樣點,平 均 OI值為0.04,並不算太低,不過這3個樣點都 在 21~24 km之間,是否意味食蟹 在棲蘭林區 分布較為侷限?這可能需要針對食蟹 的偏好 棲地做較詳細的調查。黃喉貂在本調查中僅於 整理林 R2樣區,即160林道6 km棧道區出現過 1次,是所有紀錄動物中出現率最低的一種。
該 筆 紀 錄 攝 得 時 間 為 2010年12月22日06時30 分,根據林區工作人員的敘述,也曾在同一個 月於 170林道目擊兩隻黃喉貂。查閱過去有關 鄰近雪山山系動物相調查幾乎沒有黃喉貂出現 (Wang and Wang 1993, Huang 2004, Lee 2007, Su 2008, Tsai and Pei 2008, Lin 2009a, b),僅有 大鹿林道 2筆紀錄,但報告中並未載明記錄方式 (Lee 2007)。根據早期與近年對棲蘭林區工作人 員的訪查 (unpublished data),我們推測棲蘭山 林區黃喉貂曾發生區域滅絕至少 25年。黃喉貂 為大型掠食性動物,可能扮演制約食草動物的 重要生態角色,目前其是否已在此建立穩定的 族群?未來有沒有可能會再度消失或持續順雪 山東北稜往明池、松羅湖甚至低海拔的哈盆保 留區方向擴展?我們認為值得持續進行監測與 關注。
本研究以主要物種的相對豐量來探討牠們 對不同林地類型的偏好或適應,結果清楚的顯 示棲蘭山檜木林區的野山羊在造林地的相對豐 量高於原始林,且 OI平均值從原始林、整理林 到人工林依序遞增;同時我們也很難忽略山羌 的OI平均值在原始林、整理林及人工林依序遞 減 , 且 原 始 林 數 值 為 人 工 林 2.5倍以上,剛好 與野山羊趨勢相反的現象。由於山羌的平均OI
值離散程度甚大,分析結果並沒有得到顯著性 (Table 3),但我們認為這可能受限於樣區數目 不足,未來值得再深入探討改進。從山羌、野 山羊、獼猴這 3種優勢動物的組成差異以及兩種 草食獸的相對豐量表現,我們可以確認棲蘭檜 木林區的野生動物群聚明顯的受到林地經營方 式的影響。台灣中部的研究指出天然闊葉林的 脊椎動物多樣性與數量都明顯高於造林地(Hsieh et al. 2006);在北美太平洋西北區大尺度的研 究中,在天然更新及人工更新的幼齡林與老生 林 (old growth)之間,小型哺乳類社會結構相 似,但老生林支持的生物量明顯較高(Andrew et al. 1995)。然而在加拿大西岸英屬哥倫比亞 地區的研究則顯示,大部分小型哺乳類之數量 在皆伐或擇伐更新之幼齡林都明顯高於老生林 (Sullivan et al. 2000)。這些現象與我們的結果 說 明 了 森 林 經 營 對 野 生 動 物 族 群 有 直 接 的 影 響,但是其效應視分類群、物種而有不同。以 草食獸而言,族群量往往與光照有關,例如在 林冠層過於鬱閉的溫帶紅松林地,疏伐則有利 於食草生長而使草食獸的族群增長(Bender et al.
1997)。然而本區造林地樹冠鬱閉程度相當高,
因此我們認為影響野山羊棲地選擇應該還有其 他的因子。
在 相 同 的 林 地 型 態 中 , 山 羌 的 OI值在樣
區間變異顯著,而野山羊則否(Table 3),我們
認為與野山羊可在崎嶇的地形上行動無礙的能
力有關。根據觀察,目前許多造林地可能因過
去伐木作業對林床造成破壞,加上皆伐後林地
暴露、殘根腐爛分解後土壤流失,使得岩石裸
露,造成崎嶇的地貌,至今無法復原。而從拍
攝 的 影 像 中 可 以 見 到 長 鬃 野 山 羊 在 障 礙 物 中
間跳躍攀爬,反之,山羌、山豬則沒有在這種
環境中活動的能力。野山羊在造林地或許能因
競爭優勢而獲得更多的資源,但是目前看來山
羌與野山羊在各樣區的出現頻度並沒有負相關
(r
2= 0.09, 10 d.f., p = 0.34),因此對於野山羊
偏好或適存於造林地之原因,還需要針對棲地
組成結構、資源利用等進行更全面的研究。至
於台灣獼猴在整理林以及造林地中分別都有樣
區間的差異,由於獼猴主要為成群活動,加上
其為樹棲性,因此我們尚不能對這種現象提出 推論。
棲 蘭 林 區 的 枯 倒 木 整 理 林 經 過 數 十 年 的 生息,至少在主要的喬木層已經恢復至接近原 始林的狀態(Chen and Mao 2007),Yuan et al.
(2004)於160林道兩側的鳥類群聚調查結果認 為,原始林與整理林之間鳥種數並無差異,但 群聚、功能群的結構並不相同。而在拉拉山區 中海拔的天然林與林相改良區的鳥類群聚調也 有相似的結果(Fang 1996),但也有研究顯示疏 伐對小型哺乳類族群沒有影響 (Christian et al.
1996, Tsai 2001)。以目前來說,我們發現原始 林與整理林的主要哺乳動物種類相對豐度都沒 有顯著差異,這應該是因為歷經長期的演替復 原 的 結 果 。 而 雖 然 前3排名的動物相對組成在 林地型態間明顯具有差異,但是PCA因素負荷 量圖則顯示,原始林與整理林哺乳動物群聚的 區分並不明顯。由於物種種數不多,如果要從 PCA因素負荷量圖中得到更清楚的結果,可能 須要增加稀少種的資料精確度,也需更大量的 取樣。總之,我們認為過去的枯倒木整理作業 必然對野生動物產生衝擊,但歷經長期復原,
目前動物相與原始林差異不大,至於皆伐後的 更新造林地所受的影響則比較長遠。
本研究以同一樣區所有樣點的平均值作為 樣本,以降低自動相機調查很容易出現的數據 異質性。這樣的設計使我們在野山羊的比較性 研究獲得滿意的成果,而其他較稀有的物種也 得到一定的改善。然而受限於野外工作執行困 難,為合理分配樣區避免產生偽重複,反而使 得樣本數較為不足,這個情形在山羌的 OI值分 析上就特別明顯。未來在實驗設計上建議可以 較少的樣點來平均取樣偏差,如此則樣區的面 積可以縮小,有利於設計較多的樣本數,以提 高統計檢定品質。
致 謝
本研究承蒙行政院退除役官兵輔導委員會 森林保育處、棲蘭工作站給予諸多行政上的支 援,國立東華大學張世杰教授給與物資及事故
連繫上的協助,鄭浩鈞、謝家航、鄭惟仁、許 詩涵、葉文琪、呂芳芊等諸位先生協助樣區規 劃、野外調查與資料收集建檔;芮久玟女士協 助英文撰寫校訂;以及兩位審稿人細心斧正,
特此一併申謝。
引 用 文 獻