行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
結構相似多酚類化合物對 EGF 誘導訊活化之影響
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:▉個別型計畫 □整合型計畫 計畫編號:NSC90-2314-B-038-041-
執行期間:90 年 08 月 01 日至 91 年 07 月 31 日
計畫主持人:沈杏娟
共同主持人: 本成果報告包括以下應繳交之附件:
□ 赴國外出差或研習心得報告一份
□ 赴大陸地區出差或研習心得報告一份
□ 出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:台北醫學大學醫學系皮膚學科
中 華 民 國 91 年 10 月 8 日
行政院國家科學委員會專題研究計畫成果報告
計畫編號:NSC 90-2314-B-038-041 執行期限:90 年 8 月 1 日至 91 年 7 月 31 日
主持人:沈杏娟 台北醫學大學醫學系皮膚學科
中文摘要
上皮成長因子(EGF)在調控細胞成長與增 生上扮演重要角色,其誘導細胞增生是經 由誘導細胞膜上受器之磷酸化,進而活化 其訊息傳導路徑。在皮膚惡性腫瘤如上皮 細胞癌、乾癬病灶處等多種皮膚疾病上都 發 現 有 EGF 受 器 大 量 表 現 或 細 胞 內 tyrosine 磷 酸 化 被 不 正 常 活 化 的 情 形 發 生。因此,對 EGF 誘導之訊息活化有專一 性抑制作用之化合物,當可作為未來預防 或治療與 EGF 調控異常有關之皮膚疾病 用藥。
多酚類化合物為天然存在的植物成分中主 要化合物之一,先前的文獻已證實天然多 酚類化合物具備多種特殊的生物活性,例 如抗氧化、預防腫瘤生成等。根據預試驗 結果顯示特殊種類之 flavonoid 對 TPA 誘 導之血癌、腸癌細胞增生有明顯的抑制作 用。為了更進一步瞭解其詳細的作用機轉 與方式,本計劃選擇過去在 EGF 研究中最 常被使用的 A431 細胞作為計劃之細胞模 型細胞, 並且由先前篩選所得較具潛力的 類黃素中選用一系列結構相似的 flavonoid 化合物 2’-OH flavanone, 3’-OH flavanone, 4’-OH flavanone, 5’-OH flavanone 6’-OH flavanone, 7’-OH flavanone, 4’-OCH3 flavanone, 6’-OCH3 flavanone, 7-OCH3 flavanone, 5-OCH3 flavanone, narigenin (4’, 5, 7-trihydroxyflavanone), naringin (4’, 5, 7-trihydroxyflavanone 7-rhamnoglucoside), taxifolin (3, 3’, 4’, 5,
7-pentahydroxyflavanone), baicalin, kaempferol, quercetin, morin 及 myricetin.
來探討其在 A431 細胞中對 EGF 誘導之訊 息活化之影響。
關鍵詞:上皮成長因子(EGF) 、上皮成長 因子受體(EGFR)、A431、類黃 素(Flavonoid)
Abstr act
Epidermal growth factor receptor (EGFR)-mediated tyrosine phosphorylation may be a primary indicator of signal transduction regulating cell growth and proliferation. Enhanced tyrosine kinase activity due to aberrant or overexpression of receptor or non-receptor tyrosine kinases has been implicated in a variety of skin diseases including different type of skin cancer and psoriasis. Therefore, agents inhibited effectively on EGF mediated phosphorylation and signal transduction might be a good candidate for the prevention of cancer formation.
Flavonoids are among the best candidates for mediating the protective effect of diets rich in fruits and vegetable with respect to cancer formation. Our unpublished data appeared that flavonoids were able to inhibit TPA-induced cellular proliferation in leukemia and colon cancer. To gain additional information about their growth effects on tumors and their cellular action
mechanisms, a series of structure related polyphenols including 2’-OH flavanone, 3’-OH flavanone, 4’-OH flavanone, 5’-OH flavanone 6’-OH flavanone, 7’-OH flavanone, 4’-OCH3 flavanone, 6’-OCH3 flavanone, 7-OCH3 flavanone, 5-OCH3 flavanone, narigenin (4’, 5, 7-trihydroxyflavanone), naringin (4’, 5, 7-trihydroxyflavanone 7-rhamnoglucoside), taxifolin (3, 3’, 4’, 5, 7-pentahydroxyflavanone), baicalin, kaempferol, quercetin, morin and myricetin.were added to cultures of epidermal carcinoma cells A431 treated with epidermal growth factor (EGF).
Keywor ds:Epidermal growth factor (EGF), Epidermal growth factor receptor (EGFR),flavonoid
前言
Epidermal growth factor (EGF) is a potent mitogen for a variety of mammalian epithelial tissues and cells. There is evidence to indicate that the initial interaction of EGF with target cells occurs on specific membrane receptors, i.e. EGF receptors. In normal human epidermis, EGF receptors are localized primarily on mitotically active basal keratinocytes and diminish in number as the cells differentiate. But, what does EGF do in skin? The best studied effects of EGF are its ability to increase proliferation, differentiation, and repair of epithelial and nonepithelial tissues.EGF increases the migration of both fibrolasts and keratinocytes in culture. Besides, Some of skin diseases are assoicated with over-expression of EGF or EGFR disfunction. In skin diseases in which kertinocytes abnormally differentiate, the distribution and number of EGFR change.
For example, in psoriasis vulgaris, the
number of EGF is increased twofold to fourfold in active lesions because the EGFR persistently are expressed in the abnormally differentiating stratum spinosum and corneum. Similarly in epithelial cancer at other body sites such as squamous cell carcinomas of the lung, epidermoid carcinoma of vulva.
Tyrosine kinases play an important role in the regulation of proliferation and differentiation. Enhanced protein tyrosine kinase activity due to overexpression of receptor or protein tyrosine kinase activity leads to a continuous signaling resulting in an uncontrolled cell proliferation 〔1-3〕.
Growth factor receptor tyrosine kinases activated by the binding of their specific ligands undergo auto-phosphorylation and phosphorylate various cellular substrates to carry out signal tranduction in the cells 〔4, 5 〕. The receptors with tyrosine kinases activity have an extracellular ligand-binding domain and an intracellular tyrosine kinase domain responsible for tranducing the mitogenic signal in cells. Several transmembrane tyrosine kinase receptors have been found including epidermal growth factor (EGF) receptor, platelet derived growth factor (PDGF) receptor, and colony stimulatory factor-I (CSF-I) receptor. The overexpression of these growth factor receptors with activation of associated tyrosine kinase activity has bee found detected in human cancers such as breast and liver cancers 〔 6-7 〕 . These results indicated that activation of tyrosine kinases associated growth factor receptors played an important role in the process of carcinogenesis.
The mature EGFR is an Mr 170,000 transmembrane glycoprotein that is overexpressed in a number of human malignancies, and epidermal growth factor (EGF) receptor signal pathway is recognized as an important pathway in the development of some human tumors〔8 〕 Amplification or overexpression of the EGF receptor in certain tumor types is related to cell growth and tumorigenicity 〔9 , 10 〕. EGF and TGF-α are natural ligands for EGFR that initiate cellular responses by binding to specific receptors on the surfaces of the target cells. Like several other growth factors, the binding of EGF to its cell surface receptor triggers a cascade of intracellular events, including activation of a tyrosine kinase activity intrinsic to the EGFR. These events lead to increased DNA synthesis and cell proliferation in most of cells. A431 cells have high levels of EGFR, but mitogenic response to EGF does not correlate with their increased EGF binding capacity. Previous studies demonstrated that low concentrations of EGF (0.01 nM) stimulate cell proliferation, while high concentration of EGF (10 nM) inhibit cell proliferation in A431 cells 〔11 〕. Therefore, the relationship between biological effects of EGF and tyrosine kinase activity is complex.
Reactive oxygen species (ROS) are generally considered cytotoxic, because of the oxidative damage they can cause to cellular components. However, at low concentrations, ROS may function as physiological mediators of cellular responses. For example, H2O2 mimics the stimulatory effects of insulin on glucose transport and lipid synthesis in adipocytes
〔1 2, 13 〕. Recently, the production of ROS has been detected in a variety of cells stimulated with cytokines or peptide growth factors such as IL-1, TNF-α, TGF-β and PDGF 〔14, 15,16〕. Bae et al reported that EGF-induced EGFR phosphorylation was mediated by H2O2 formation 〔17 〕.
These data indicated that ROS was involved in EGF induced signal transduction.
Previous paper proposed that compounds blocked EGF mediated signal pathway have potential to be antiproliferative agents in tumors dependent on EGF for growth 〔18 〕. Therefore, development of effective blocking agents on EGF mediated cellular response is the major purpose of the present study. In order to develop the effective compounds on the inhibition of EGF-mediated cellular proliferation, there are 6 major experimental items involved in the present study.
1. Study the inhibitory effects of flavonoids on EGF-induced proliferation in A431 cells and the cytotoxicity of flavonoids by MTT assay, trypan exclusion and
3H-thymidine incorporation.
2. Study the effects of flavonoids on the binding of EGF to its EGFR by receptor binding assay.
3. Study the inhibitory activities of flavonoids on EGFR phosphorylation induced by EGF in A431 cells by western blotting using anti-phosphotyrosine antibody.
4. Previous report indicated that EGF-mediated signal transduction was through H2O2 production. Study the
antioxidative activity of indicated flavonoids on EGF induced H2O2 production in A431 cells was involved in the present study.
5. Activation of MAPK cascade had been demonstrated to be downstream of EGFR phosphorylation. Therefore, investigating the effects of flavonoids on EGF-induced MAPK (ERK1/ERK2) phosphorylation will be involved in the present study.
6. SAR (Structure Activity Relationship) of these flavanones will be discussed and proposed here based on the results of their inhibitory intensities on EGF-induced responses.
The results of the present study provided more molecular evidences in development of effective inhibitors on EGF induced signal transduction for further study.
結果與討論
EGF 誘導 A431 增生
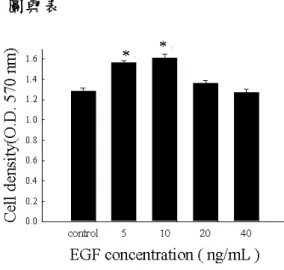
利用 MTT assay 進行本實驗,結果顯示以 10 ng/ml 的 EGF 處理 2.5 × 105個的 A431 細胞,在 4 小時後能產生最佳的促進增生 效果。(Fig.1、Fig. 2)
EGF 誘導 EGFR 磷酸化, ERK 磷酸化表 現之時間依循性
MAPK 瀑布效應( Mitogen activated protein kinase cascade )調控多種細胞的生理、生化 反應,特別是在細胞的增生作用中扮演相 當重要的角色,更是調控其他多種相關基 因表現及訊息傳遞的重要途徑,且在癌細 胞的增生過程中,更常呈高度磷酸化的表 現 〔19 〕。由西方墨點試驗 (Fig.3)結果 顯示,EGF 在 10 分鐘便可促使 EGFR 和 ERK 蛋白質磷酸化。之前的文獻指出,
EGFR 的磷酸化是 tyrosine kinase 的位置被
磷酸化,而 PY–20 的抗體可辨識 tyrosine kinase 的位置有被磷酸化的蛋白質的表 現;因此我們利用 PY-20 在 180 kDa 的位 置來確認 EGFR 的確是 tyrosine kinase 的 位置被磷酸化而可以被 PY-20 的抗體偵 測到;除此我們亦利用 active form EGFR 的 抗 體 專 一 性的 偵 測 180 kDa 位置的 EGFR 蛋白質表現量。
EGF 對 A431 細胞中 J NK 及 p38 的影響 由先前的研究得知,MAPK cascade 有三 種 subunits 分別為 MAPK,JNK 與 p38。
本實驗欲探討 JNK 與 p38 是否參 EGF 所 誘導的細胞增生作用。由西方墨點法試驗 結果顯示 JNK 與 p38 不會受到 EGF 的影 響而產生磷酸化的變化(Fig.4)。因此,JNK 和 p38 並不是 EGF 所誘導的細胞增生的主 要路徑。
PD98059 抑制由 EGF 所誘導細胞增生之 劑量依循性
PD98059 為 專 一 性 phosphorylated MAPK/ERKs 的抑制 劑 〔20 〕。 以 MTT assay 觀察 PD98059 對 A431 細胞生長的影 響(Fig.5)。由結果顯示,PD98059 在 20 、 40、80 及 160µM 等不同濃度的投予下,
能對 EGF 所誘導的 A431 細胞增生產生劑 量依循性的抑制效果。也就是說,利用 MEK 專一性抑制劑的處理,能抑制由 EGF 所誘導的細胞增生作用,因此證明 EGF 所誘導的細胞增生可能主要是透過 ERK 這個路徑。
EGF 誘導 COX-2 表現之時間依循性 Cyclooxygenase-2 (COX-2) 此一致發炎物 質在近年的研究中被發現可能與腫瘤的形 成有關,特別是在腸癌的發生上有相當之 關聯性 〔21 〕。那麼 COX-2、EGF 及 A431
細胞三者之間是否有相關聯?利用西方墨 點法來探討這三者之間的影響。結果顯 示,在加入 EGF 80 分鐘之後,COX-2 蛋白質被大量的活化, COX-1 蛋白質則 不受到影響。因此,在 EGF 所誘導的訊息 傳遞中,COX-2 可能參與其中,而 COX-1 則較無關係(Fig.6)。
EGF 誘導之 MAPK 磷酸化與 COX-2 活 化的關聯性
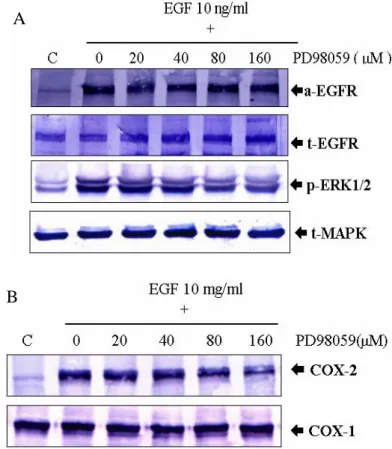
由前面實驗已經知道 ERK 和 COX-2 與 EGF 所誘導的細胞增生間有著極密切的 相關性,但這二種蛋白質是如何影響由 EGF 所誘導的細胞增生呢?本實驗中,利 用 20、40、80 或 160µM 等 不同濃度的 PD98059 對 A431 細胞進行前 處理 30 分鐘,再以 EGF 10 ng/ml 處理 10 分鐘及 160 分鐘,最後以西方墨點法進行分析。
EGF 10 ng/ml 處理 10 分鐘組觀察 ERK 的變化,處理 160 分鐘組觀察 COX-2 的變 化(Fig.7A, Fig.7B)。由 Figure.7 的實驗證 據顯示,在 20、40、 80 及 160µM 等不同 濃度的 PD98059 投予下,對 EGF 所誘導 的 phosphorylated MAPK/ERKs 蛋白質表 現產生劑量依循性的抑制作用,有趣的是 在 160 分鐘之時同時抑制了 COX-2 蛋白 質的表現,但對 EGFR 和 PY-20 蛋白質的 表 現 並 未 有 太 大 的 影 響 ; 所 以 我 們 推 測:COX-2 的基因表現位於 ERK 的下游。
由此結果可以證明 EGF 誘導細胞增生的 訊 息 傳 遞 順 序 為 : EGF 結 合 到 其 受 器 ( EGFR )上,促使受器( EGFR )磷酸化,接 著再透過 MAPK/ERKs 的磷酸化,進而調 控下游 COX-2 蛋白質的表現以達到促進 細胞增生的表現。
COX-2 活化與 PGE2生成對 EGF 誘導細 胞增生的重要性
NSAIDs(non-steroids anti-inflammatory
drugs)非類固醇抗發炎藥物在臨床上常用 於治療前列腺疾病,其可能的機制主要是 藉由抑制 COX-2 酵素的活性進而達到抑 制前列腺素生成的目的。本實驗中,將預 處理無血清培養液的 A431 細胞加入不同 濃度梯度的 NSAIDs:Aspirin, Diclofenac 與 Indomethacin 30 分 鐘 之 後 再 加 入 EGF 10 ng/ml , 24 小時後進行 MTT assay。結果顯示 NSAIDs 類的藥物確實有 效 的 抑 制 由 EGF 所 誘 導 的 細 胞 增 生
(Fig.8)。
PGE2生成對 EGF 誘導細胞增生的重要性
Prostaglandin E2 (PGE2)是 arachidonic acid (花生四烯酸)經 COX-2 酵素反應的代謝產
物,目前的研究發現 PGE2 具有促進腫瘤
形成的特性,特別是在 COX-2 的表現及腸
癌的發生上有相當之關連性〔22 〕。故我們
也想了解 PD98059 和 NSAIDs 對 EGF 所 誘導產生之細胞增生抑制是否也會經由外 加 PGE2 的方式來達到回復? 如(Fig. 9)
所示,外加 PGE2 1µg/ml 的確可以回復 PD98059 和 NSAIDs 抑制 EGF 所誘導之 細胞增生作用。因此,我們推測 EGF 所
誘導的細胞增生過程中,PGE2 扮演舉足
輕重的角色。但 PGE2 究竟是透過何種機
轉或路徑來達到細胞回復的目的呢?由之 前的證據顯示,EGF 所誘導的細胞增生是 透過 ERK 這個路徑,所以我們推測外加
PGE2 使得原本被抑制的細胞可以繼續走
向增生有可能也是透過相同的路徑;因 此,我們想利用西方墨點法來確認在添加 PGE2之後,ERK 甚至 COX-2 的蛋白質表 現是否會回 復(Fig.10 、Fig.11)?結果顯 示,處理 PD98059 160µM 可以明顯抑制 ERK 和 COX-2 蛋白質的表現,若額外處 理 PGE2 1µg/ml 可以使原來受抑制 COX-2 的蛋白質得到回復的表現但對於 ERK 則
無明顯差異;此證據除了顯示 PGE2 在整 個 EGF 所誘導的細胞增生中的角色外,更
進一步確定 PGE2 會使原本受到抑制增生
的 A431 細胞恢復生長能力是透過 COX-2 這個路徑。
Flavonoids 對 EGF 誘導 MAPK 磷酸化, EGFR 及 COX-2 表現之抑制作用
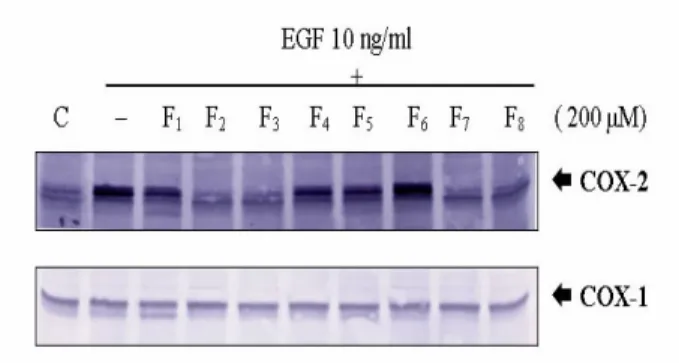
以上的實驗證據顯示,EGF 能分別在 10 分 鐘 及 160 分 鐘 誘 導 phosphorylated MAPK/ERKs、EGFR、COX-2 的蛋白質表 現,因此本實驗利用西方墨點法探討八種 flavonoids ( F1: flavanone, F2: 2'-OH flavanone F3:4'-OH flavanone, F4:6-OH flavanone, F5:7-OH flavanone, F6:taxifolin, F7:wogonin, F8:fisetin。)對 EGFR 磷酸 化的影響。處理 200 M flavonoids 並分 別觀察其在 10 分鐘對 EGF 10 ng/ml 所誘 導 phosphorylated MAPK/ERKs、EGFR 蛋 白質表現,及 160 分鐘對 EGF 10 ng/mL 所誘導的 COX-2 蛋白質表現的抑制能力 (Fig.12)。結果顯示,此八種結構相似的 flavonoids 在 200µM 和 EGF 10 ng/ml 同時 投 予 於 serum-depleted A431 細 胞 時 , 4'-OH flavanone ( F3 )及 fisetin( F8 )能對 EGF 所誘導的 EGFR 和 ERK 蛋白質表現 產生明顯的抑制作用,另外, F2 ( 2'-OH flavanone )、F3 ( 4'-OH flavanone ) 和 F7
( wogonin )、F8 ( fisetin )對 EGF 所誘導的 COX-2 蛋白質表現有最明顯的抑制作用 (Fig.13)。
Flavonoids 對 EGF 誘導 ERK 磷酸化及 COX-2 表現之劑量依循性抑制作用
由 Figure.12 和 13 的結果顯示,200µM 4'-OH flavanone 和 fisetin 對 EGF 10 ng/ml 所誘導的 ERK 磷酸化及 COX-2 蛋白表現 有明顯抑制作用;故針對此二種化合物以
西方墨點法就磷酸化的 ERK 和 COX-2 蛋 白表現進行其濃度依循性抑制與否的評估 (Fig.14、Fig.15、Fig.16、Fig.17),4'-OH flavanone 和 fisetin 此二個化合物在抑制 EGF 誘導的上述細胞反應呈現一劑量依 循性抑制現象。
PGE2在 flavonoids 所抑制 A431 細胞增生 中的角色
先前的結果已經指出 Flavonoids 類的化合 物可以抑制 EGF 所誘導之細胞增生作
用,但是否與 PGE2 確有關聯性?故我們
先 預 處 理 200 µM 4'-OH flavanone 和 fisetin,再外加 PGE2之後,發現 A431 細 胞受到抑制的情形可以得到回復;故推測 flavonoids 在抑制 A431 細胞增生的過程中 或 許 和 抑 制 PGE2 的 生 成 也 有 相 關 性 (Fig.18、Fig.19)。
Flavanoids 對 EGF 誘導細胞群落增生的 抑制能力探討
由 以 上 的 實 驗 結 果 觀 察 發 現 , 4'-OH flavaone 及 fisetin 結構相似之 flavonoids 化合物能針對 EGF 誘導的腫瘤促進作用 有明顯的抑制效果;而為了提供更直接的 證據來證明此二個化合物的抑制能力,利 用軟瓊脂實驗(soft agar assay)來對此二 個 flovanoids 化合物的抗腫瘤促進能力進 行更進一步的機制探討(Fig.20)。對照組的 A431 cells 於正常的培養條件下(DMEM medium, 10% fetal calf serum and 1%
antibiotics)依正常的型態生長,並沒有明 顯的細胞群落產生;而以 EGF 10ng/ml 處 理的 A431 cells 於相同的培養環境下,於 二個星期後開始有細胞群落產生,同時處 理 PGE2 1µg/ml 的 A431 細胞細胞群落的 產生更為明顯。一個月後觀察 EGF 10 ng/ml 處理的 A431 cells 的細胞群落產生
極為明顯;而當 4'-OH flavaone 及 fisetin 各以 200µM 的濃度與 EGF 10 ng/ml 同時 投予 1 個月後,EGF 誘導細胞群落的產生 明顯的被抑制,且細胞型態上與對照組細
胞維持相同。而且若額外加 PGE2 會使的
原本受到抑制的 A431 細胞群落再度增加
證實 PGE2 的存在的確對這整個的細胞增
生很重要。由以下顯微鏡的觀察計數細胞 群落結果顯示,此二個 flavonoids 化合物 能有效且無明顯毒性的抑制 EGF 誘導的 細胞群落產生及細胞型態之改變。
Flavonoids 對 EGF 誘導產生之 PGE2之 抑制探討
Arachidonic acids 會經由 cyclooxygenase 的 活 化 代 謝 後 產 生 prostaglandins , 而 prostaglandins 在近年的研究中被認為與 腫瘤的形成有關,特別是在 COX-2 的表 現及腸癌腫瘤的發生上有相當的關聯性。
故實驗中以 PGE2 enzyme immunoassay ( EIA ) kit 針對 PD98059、NSAIDs、4'-OH flavanone 和 fisetin 對 EGF 所誘導產生之 PGE2 探 討 其 抑 制 能 力 。 分 別 收 集 PD98059 80、160µM、Aspirin 2、4 mM、
Diclofenac 100、200µM、4'-OH flavanone 和 fisetin 100、200µM 和 EGF 10 ng/ml 同 時投與在 serum-starvated 的 A431 的培養 液中,利用 EIA kit 進行分析,並利用 ELISA reader 於 430 nm 偵測定量 PGE2的 含量變化。由實驗結果發現 EGF10 ng/ml 能誘導 A431 細胞產生較對照組高出 4 倍 的 PGE2含量,而我們所處理的 compound 均能對此 EGF 所誘導產生之 PGE2 有明 顯的抑制作用(Fig.21)。
圖與表
Figure 1. Effect of EGF induced A431 cell prolifer ation. EGF at the dose of 10 ng/ml is an optimal dose to induce A431 cell proliferation . Cell proliferation was detection by MTT assay ( O.D.570 nm ). p values were calculated by student's t test. Results are representative of three independent experiments. * Statistically significant induction in EGF 10 ng/ml compared with control. ( P <0.05 )
Figure 2. Time-dependent of EGF induced A431 cells prolifer ation. EGF 10 ng/ml is stimulate A431 cell proliferation at the time of 24 hr obviously induce A431 cells proliferation.
Cell proliferation was detection by MTT assay.
p values were calculated by student's t test.
Results are representative of three independent
experiments.** Statistically significant induction in EGF 10 ng/ml compared with control.( P <0.01 )
Figure 3. Induction of EGFR and MAPK/ERKs phosphoylation in EGF stimulated A431 cells. A431 cells were stimulated with EGF 10 ng/ml for different time periods( 10 to 160 min ) and the cell lysate ( 50 µg )was resolved on a 8% SDS-PAGE, transferred to a PVDF membrane, followed by Western blotting with antiphosphorylated EGFR and MAPK/ERK1/2 or anti-EGFR and ERK1/2 and α-tubulin.
FFigiguurree 4.4. PP3838-- aanndd JJNKNK-m-medediaiatteded sisiggnnaall ttrraannsdsduuctctioionn mamayy nnotot bbee ininvovollvveedd iinn EEGGFF-i-inndduucecedd pprrololiiffeerraattivivee eeffffeeccttss iinn AA443311 c
ceellllss.. A431 cells treated with EGF 10 ng/ml for the different periods ( 10 to 160 min ).Cells were then harvested, lysed and subjected to Western blot analysis with antibodies that recognize phosphorylated JNK and p38 and anti-JNK and p38.
Figure 5. MAPK specific inhibitor PD98059 dose-dependently inhibited EGF-induced A431 cell prolifer ation. A431 cells were pretreated with PD98059 for different dose ( 20 to 160 µM) for 30 min and then treated with EGF 10 ng/ml for 24 hr. p values were calculated by student's t test. Cells were then harvested and subjected by MTT assay. **
Statistically significant attenuation in PD98059 40 ng/ml compared with EGF 10 ng/ml treated cells. ( P <0.01 )
Figure 6. Effects of EGF-induced COX-2 expression. A431 cells were seeded at a density of 2.5× 105 cells/well in 60 mm plates. The next day, cells were starvated for serum-free medium for 24 hr. After 24 hr, A431 cells were re-treated with fresh serum-free medium containing EGF 10 ng/ml for 160 min and Western blot analysis of the expression COX-2 and COX-1. Proteins recognized by each antibody are indicated on the right hand side. The cells were harvested at different time points. 50 µg protein were extracted and analyzed in each lane.
Figure 7. MAPK specific inhibitior PD98059 inhibited EGF induced MAPK phosphor ylation associated with suppressing COX-2 induction. A431 cells were treated with different dose ( 20 to 160 µM ) for 30 min prior to stimulation for 10 min with EGF 10 ng/mL for phosphorylated EGFR and ERK1/2 ( A ) or stimulation for 160 min for COX-2 ( B ). The
A
B
activation state of EGFR and ERK1/2 was determined by SDS-PAGE separation of 50 µg of whole cell lysate followed by Western blot with anti-phospho-EGFR and ERK1/2 and COX-2 antibodies.
Figure 8. NSAIDs inhibited EGF-induced PGE2 production. A431 cells were pretreated NSAID ( including Aspirin 2 to 4 mM, diclofenac 50 to 100 µM and indomethacin 100 to 200 µM ) for 30 min and treated with EGF 10 ng/ml for 24 hr. Cell viability was detected by MTT assay. p values were calculated by student's t test. Cells were then harvested and subjected by MTT assay. ** Statistically significant attenuation in NSAID compared with EGF 10 ng/ml treated cells. ( P <0.01 )
Figure 9. Rever sal by the extr a PGE2of the suppressive effect of PD98059 and NSAIDs on EGF-induced prolifer ation.
Serum-starvated A431 were treated with ( A )PD98059 ( 160 µM ) , ( B ) NSAID ( Aspirin and diclofenac ) for 30 min and then treated with EGF 10 ng/ml and PGE2 1 µg/ml.
Cell density was measured by MTT assay.
( A ) .Asterisk ( * ) denotes a significant differences with P < 0.01 between the group with EGF 10 ng/ml treatment. # denotes a significant differences with P < 0.01 between the PD98059 group with or without PGE2 1 µg/mL treatment. ( B ). Asterisk ( * ) denotes a significant differences with P < 0.01 between the aspirin group with or without PGE2 1 µg/ml treatment. # denotes a significant differences with P < 0.05 between the diclofenac group with or without PGE2 1 µg/ml treatment.
B
Figure 10. The suppressive effect of PD98059 on EGF-induced ERK expression can not rever sed by PGE2. Serum-starvated A431 cells were pretreated PD98059 80 and 160 µM for 30 min then treated with EGF (10 ng/ml) and PGE2 1 µg/ml for 10 min and the expression of protein was detected by Western blot with anti-phospho-ERK1/2 and anti-ERK1/2 antibodies.
Figure 11. The suppressive effect of PD98059 on EGF-induced COX-2 expression can be rever sed by PGE2. Serum-starvated A431 cells were pretreated PD98059 80 and 160 µM for 30 min then treated with EGF ( 10 ng/ml ) and PGE2 1 µg/ml for 160 min to detect COX-2 fand COX-1 protein expression by Western blot.
Figure 12. Inhibitor y effects of flavonoids on EGF induced phor ylated MAPK/ERKs expression. Cells were treated with EGF 10
ng/ml with indicated compound ( 200 µM ) for 10 min and the protein expression were detected by 8 % SDS-PAGE separation of 50 µg of whole cell lysate followed by Western blot with anti-phospho-EGFR and ERK1/2 antibodies.
F1:Flavonone、F2:2'-OH flavanone、F3:4'-OH flavanone F4:6-OH flavanone 、 F5:7-OH flavanone 、 F6:taxifolin 、 F7:wogonin and F8: fisetin.
Figure 13. 2' -OH flavanone 4' -OH flavanone and fisetin showed the significant inhibition on EGF-induced COX-2 expression. Cells were treated with EGF 10 ng/ml with indicated compound ( 200 µM ) for 10 min and the protein expression were detected by 8 % SDS-PAGE separation of 50 µg of whole cell lysate followed by Western blot with anti-COX-2 and COX-1 antibodies.
Figure 14. 4' -OH flavanone showed the dose-dependent inhibition on EGF-induced MAPK and EGFR phosphor ylation. A431 cells were pretreated with 4'-OH flavanone ( 25
to 200 µM ) for 30 min and then treated with EGF 10 ng/ml for 10 min and the phosphorylation ERK1/2 protein expression was detected by Western blot.
Figure 15. 4' -OH flavanone showed the dose-dependent inhibition on EGF-induced COX-2 phosphor ylation. A431 cells were pretreated with 4'-OH flavanone ( 25 to 200 µM ) for 30 min and then treated with EGF 10 ng/ml for 160 min and COX-2 and COX-1 protein expression was detected by Western blot
.
Figure 16. Fisetin showed the dose-dependent inhibition on EGF-induced MAPK and EGFR phosphor ylation. A431 cells were pretreated with fisetin ( 25 to 200 µM )for 30 min and then treated with EGF 10 ng/ml for 10 min and the EGFR and ERK protein expression was detected.
Figure 17. Fisetin showed the dose-dependent inhibition on EGF-induced COX-2 phosphor ylation. A431 cells were treated fisetin ( 25 to 200 µM ) for 30 min and then treated with EGF 10 ng/ml for 160 min and the COX-2 and COX-1 protein expression was detected. The activation state of COX-2 was determined by SDS-PAGE 8 % separation of 50 µg of whole cell lysates followed by Western with anti-COX-2 and COX-1.
Figure 18. Rever sal by the extr a PGE2 of the suppressive effect of 4' -OH flavanone on EGF-induced prolifer ation.
Serum-starvated A431 were treated with 4'-OH flavanone (25 to 200 µM ) for 30 min and then treated with EGF 10 ng/ml and PGE2 1 µg/ml.
Cell density was measured by MTT assay.
Asterisk ( * ) denotes a significant differences with P < 0.01 between the group with EGF 10
ng/ml treatment. # denotes a significant differences with P < 0.01 between the group with or without PGE2 1 µg/ml treatment. Data were described as mean ± SEM of at least three independent experiments.
Figure 19. Rever sal by the extr a PGE2 of the suppressive effect of fisetin on EGF-induced prolifer ation. Serum-starvated A431 were treated with fisetin (25 to 200 µM ) for 30 min and then treated with EGF 10 ng/ml and PGE2 1 µg/ml. Cell density was measured by MTT assay. Asterisk ( * ) denotes a significant differences with P < 0.01 between the group with EGF 10 ng/ml treatment. # denotes a significant differences with P < 0.01 between the group with or without PGE2 1 µg/ml treatment. Data were described as mean ± SEM of at least three independent experiments.
* P< 0.05, ** P< 0.01 Data were described as mean ± SEM of at least three independent experiments.( * P< 0.05, ** P< 0.01 versus EGF 10 ng/ml, ## versus fisetin 200 µM )
Figure 20. Significant attenuation of EGF-induced PGE2 production by indicated compounds. Serum-starvated A431 were treated with PD98059 80 to 160 µM、Aspirin 2 to 4 mM、Diclofenac 100 to 200 µM、4'-OH flavanone and fisetin 100 to 200 µM. After 24 hrs, medium were detection by 430 nm. Asterisk ( * ) denotes a significant differences with P <
0.01 between the group with EGF 10 ng/ml treatment. ## denotes a significant differences with P < 0.01 between with control.
Figure 21. Supression of EGF-induced colonies by 4' -OH flavanone and fisetin. A431 cells were treated with EGF 10 ng/ml with or without indicated compounds for 2 weeks. Data were described as mean ± SEM of at least three
independent experiments. Asterisk ( * ) denotes a significant differences with P < 0.01 between the group with control treatment. # denotes a significant differences with P < 0.05 between fisetin 200 µM with or without PGE2 1 µg/ml.
參考文獻
1. Aaronson SA. (1991) Growth factors and Cancer. Science 254: 1146-1151, 1991.
2. Sporn MB, Roberts AB. Autocrine growth factors and cancer. Nature 313: 745-747, 1985.
3. Ahmad A, Gali H, Javed S, Agarwal R.
Skin Cancer chemopreventive effects of a flavonid antioxidant silymarin are mediated via impairment of receptor tyrosine kinase signaling and perturbation in cell cycle progression. Biochem Biophys Res Commun 248: 294-301, 1998.
4. Meuillet EJ, Mania-Farnell B, George E et al., modulation of EGF receptor activity by changes in the GM3 content in a human epidermoid carcinoma cells A431. Exp Cell Res 256:74-82, 2000.
5. Sako Y, Minoghchi S, Yanagida T. Single molecule imaging of EGFR signaling on the surface of living cells. Nature Cell Biol 2: 168-172, 2000.
6. Abramovitch R, Marikovsky M, Meir G, Neeman M, Stimulation of tumor growth by wound –derived growth factors. Brit J Cancer 79: 1392-8, 1999.
7. Reilly RM, Kiarash R, Sandhu H, Lee YW.
A comparison of EGF and Mab 528 labeled with 111In for imaging human breast cancer. J Nuclear Med 41:903-11, 2000.
8. Gullick WJ. Prevallence of aberrant expression of the epidermal growth factor
receptor in human cancers. Br Med Bull 47:
87-98, 1991.
9. Carpenter G, King L, Jr., Cohen S. Rapid enhancement of protein phosphorylation in A431 cell membrane preparations by epidermal growth factor. J Biol Chem 254:
4884-4891, 1979.
10. Gill GN, Buss JE, Lazar CS, Lifshitz A, Cooper JA. Role of epidermal growth factor stimulated protein kinase in control of proliferation of A431 cells. J Cell Biochem 19: 249-257, 1982.
11. Gulli LF, Palmer KC, Chen YQ, Reddy KB.
Epidermal growth factor-induced apoptosis in A431 cells can be reversed by reducing the tyrosine kinase activity. Cell Growth &
Differ 7: 171-178, 1996.
12. Mukherjee SP, Lane RH, Lynn WS.
Endogenous hydrogen peroxide and peroxidative metabolism in adipocytes in response to insulin and sulfhydryl reagents.
Biochem Pharmacol 27: 2589-2594, 1979.
13. May JM, de Haen C. Insulin stimulated intracellular hydrogen peroxide production in rat epididymal fat cells. J Biol Chem 254:
9017-9021, 1979.
14. Lo YYC, Wong JMS, Cruz TF. Reactive oxygen species mediate cytokine activation of c-Jun NH2-termal kinases. J Biol Chem 270: 11727-11730, 1995.
15. Krieger-Brauer HI, Kather H. Antagnoistic effects of different members of the fibroblast and platelet-derived growth factor families on adipose conversion and NADPH-dependent H2O2 generation in 3T3 L1 cells. Biochem J 307: 549-556, 1995.
16. Chen Q, Olashaw N, Wu J. Participation of
reactive oxygen species in the lysophosphatidic acid-stimulated mitogen activated protein kinase kinase activation pathway. J Biol Chem 270: 28499-28502, 1995.
17. Bae YS, Kang SW, Seo MS et al., Epidermal growth factor (EGF) induced generation of hydrogen peroxide. J Biol Chem 272: 217-221, 1997.
18. Copp BR, Fairchild CR, Cornell L., Casazza AM et al., Naamidine A is an antagonist of the epidermal growth factor receptor and an in vivo active antitumor agent. J Med Chem 41: 3909-3911, 1998.
19. Hossain MZ, Jagdale AB, Ao P and Boynton AL. Mitogen-activated protein kinase and phosphorylation of connexin43 are not sufficient for the distribution of gap junctional communication by platelet-derived growth factor and tetradecanoylphorbol acetate. J.Cell Physiol.179:87-96, 1999
20. Choi C, Xu X, Oh JW, Lee SJ, Gillespie GY, Park H, Jo H. and Benveniste EN.
Fas-induced expression of chemokines in human glioma cells: involvement of extracellular signal-regulated kinase 1/2 and p38 mitogen-activated protein kinase.
Cancer Res. 6,3084-91,2001
21. Hongmiao S. Jinyi Shao, M., Kay Washimgton and Raymond N. J.Biol.Chem.
276,18075-18081,2001.
22. Wenzel U, Kuntz S, Brendel MD and Daniel H. Dietary flavone is a potent apoptosis inducer in human colon carcinoma cells. Cancer Res. 60:
4645-4653, 2000.