行政院國家科學委員會專題研究計畫 成果報告
斑馬魚 syndecan-3 基因之表現與功能鑑定
計畫類別: 個別型計畫
計畫編號: NSC94-2311-B-038-003-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 臺北醫學大學生化學科
計畫主持人: 周志銘
計畫參與人員: 蔡豐州、林尚萱
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫 ■ 成 果 報 告
□期中進度報告
(計畫名稱)
計畫類別:■ 個別型計畫 □ 整合型計畫 計畫編號:NSC 94-2311-B-038-003
執行期間: 94 年 8 月 1 日至 95 年 7 月 31 日
計畫主持人:周志銘 共同主持人:
計畫參與人員: 蔡豐州、林尚萱
成果報告類型(依經費核定清單規定繳交):■精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列 管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:台北醫學大學
中 華 民 國 95 年 10 月 30 日
計畫中文摘要(五百字以內)
關鍵字:肝素結合蛋白神經促進因子、Syndecan-3
Syndecan-3 (N-syndecan) 是哺乳類 proteoglycans 家族,四個成員之一,在哺乳類的相 關研究中顯示此基因主要會表現在神經系統,特別是在胚胎發育時期。而Syndecan-3 蛋白的 組成包含了一個穿膜區域(transmembrane domain),一個34 個胺基酸的穿膜蛋白羧基端
(C-terminal)分佈在細胞質,而且在細胞外側的部分包含許多glycosaminoglycan 結合的位 置,而此外側區域會和許多生長因子和細胞外的間質蛋白結合,扮演重要的生物功能。目前 認為這一類的蛋白可能扮演例如 FGF 受體或 integrins 的協同受體(coreceptor )角色,可 能在胚胎發育神經系統形成的過程中細胞附著 (cell adhesion)、神經樹突生長的引導 (neurite guidance) 和 細 胞 的 轉 移 (cell migration) 有 關 。 同 時 也 有 研 究 指 出 此 基 因 會 和 海 馬 廻 (hippocampus) 中突觸的多變性 (synaptic plasticity) 調節有關。
肝素結合蛋白神經促進因子(Heparin binding neurotrophic factor / Pleiotrophin ; HBNF/PTN) 是首先利用肝鹽親和性管柱進行與其結合的蛋白純化並利用高鹽所分離的肝鹽 結合蛋白,這種與肝鹽或肝鹽類似的醣類高度結合的蛋白目前研究發現這類的蛋白在生物功 能上具有相當重要的地位。Syndecan 3 蛋白外所結合的肝素硫酸鹽會和纖維母細胞生長因子 2 (fibroblast growth factor-2 ; FGF-2) 和 肝 素 結 合 生 長 因 子 pleiotrophin (heparin-binding growth-associated molecule;HB-GAM) 結合。而在體外培養的神經細胞研究中發現不論是改 變Syndecan 3 蛋白外肝鹽結合多醣類或利用Syndecan 3 蛋白的多元抗體加入培養的神經細 胞內都會抑制由肝素結合蛋白神經促進因子(HBNF/PTN-induced neurite outgrowth) 所促進的 神經纖維生長。目前有很多的研究也指出和Syndecan 3 蛋白外肝鹽結合的肝鹽類小分子都具 有抑制由肝素結合蛋白神經促進因子 (HBNF/PTN-induced neurite outgrowth) 所促進的神經 纖維生長。而先前的研究亦指出syndecan-3 可能扮演的功能是肝素結合蛋白神經促進因子 (HBNF/PTN) 的接受體,但是相關的研究還不是很明確,所以本計畫擬利用實驗室已建立的 系統進一步的探討syndecan-3 和肝素結合蛋白神經促進因子在斑馬魚胚胎發育過程中的交互 作用為何。
在先前本實驗室的研究 (Chang et al.,2004) ,利用具神經組織專一性表現的啟動子(HuC promoter) 來調控綠色螢光蛋白(green fluorescence protein, GFP),標定斑馬魚胚胎發育過程 中的神經組織,成功的建立肝素結合蛋白神經促進因子(HBNF)所誘導促進斑馬魚神經纖 維生長的活體分析系統。本研究將探討syndecan-3 (zSyn3) 基因在斑馬魚胚胎發育過程中的功 能。主要研究的目標如下:
1.將 HBNF 和完整 (zSyn-3)或羧基端缺失的dS3 (cytoplasmical truncated form of zSyn-3; dS3) 基因共轉染 (cotransfection) 入 PC12 細胞中,利用螢光標定進行細胞內此兩基因分佈的 研究,並藉由分別轉染肝素結合蛋白促進因子 (HBNF) 和共轉染HBNF 和完整或羧基端 缺失的zSyn-3 基因,對於誘導 PC12 細胞神經纖維生長的促進能力來探討配合體─接受體 (ligand-receptor) 之間的交互作用。
2.利用morpholino oligonucleotide (MO) 的技術,藉由將zSyn-3 MO 和 HBNF/ HuC-GFP 表現 質體共同以顯微注射的方式來研究,由肝素結合蛋白神經促進因子所促進的神經纖維生長
(HBNF-induce of neurite outgrowth)是否受到斑馬魚syndecan-3 基因表現降低而影響神經 生長促進能力。
3.在先前的研究我們發現在斑馬魚的胚胎發育過程中,肝素結合蛋白神經促進因子(HBNF)
可誘導並促進斑馬魚神經纖維生長。利用上述建立的活體分析系統,分別利用CMV、
glia-specific GFAP promoter 和neuron-specific HuC promoter 不同特性的啟動子來調控 zSyn-3 基因,並同時利用顯微注射的技術將具神經纖維生長促進因子的HBNF 基因分別 和不同啟動子調控的 zSyn-3 基因共同注射 (coinjection) 入斑馬魚的胚胎中,根據斑馬魚
神經纖維生長促進的能力來探討 HBNF 和 zSyn-3 兩個基因的交互反應。
4.同時也可以利用用CMV、glia-specific GFAP promoter 和 neuron-specific HuC promoter 不同特性的啟動子來調控羧基端缺失的dS3 (cytoplasmical truncated form of zSyn-3; dS3) 基 因,來探討 zSyn-3 在胚胎發育時扮演的功能。
計畫英文摘要(五百字以內)
Keywords: Syndecan-3 (N-syndecan), Heparin binding neurotrophic factor (HBNF),
Syndecan-3 (N-syndecan) is one of the four mammalian syndecans and it is mainly expressed in the nervous system, especially during development. Syndecan-3 has one transmembrane domain, a short cytoplasmic tail of 34 amino acids, and an extracellular domain that carries heparan sulfate chains. Syndecans have been suggested to function as coreceptors with other signaling receptors, such as FGF receptors and integrins. It has been suggested to function in cell adhesion, neurite guidance, and cell migration during development of the nervous system. Syndecan-3 has also been implicated in the regulation of synaptic plasticity in the hippocampus. Syndecan-3 is expressed in an activity-dependent manner in the CA1 pyramidal neurons, and application of exogenous syndecan inhibits the induction of longterm potentiation (LTP). Heparin binding neurotrophic factor/ Pleiotrophin (HBNF/PTN) was first isolated as a heparin-binding protein that was eluted from a heparin affinity column with high salt concentration. Such a high affinity-binding property suggests that heparin or heparin-type carbohydrates may play important roles in the biological function of HBNF/PTN. Syndecan 3 heparan sulphate chains bind to fibroblast growth factor- 2 (FGF-2) and heparin-binding growth-associated molecule (HB-GAM; also known as pleiotrophin).
Both the heparan sulfate side chains of syndecan-3 and polyclonal anti-syndecan-3 inhibit HBNF/PTN-induced neurite outgrowth in the cultured neurons. In addition to syndecan-3 heparan sulfate, the low molecular weight heparin displays more potent inhibition of HBNF/PTN-induced neurite outgrowth. Indeed, further studies demonstrate that a transmembrane heparan sulfate proteoglycan, syndecan-3, acts as a receptor for HBNF/PTN.
In our previous study, we established an in vivo neurite outgrowth assay in zebrafish embryos that provided a direct observation of HBNF-induced neurite outgrowth from GFP-labeled neurons during zebrafish development. Therefore, in this proposal, we will analysis zSyn-3 function during zebrafish development. Specific aims are listed below:
(1) Co-transfection of HBNF and wild-type or cytoplasmical truncated form of zSyn-3 (dS3) into PC12 cells, cellular co-localization and ligand-receptor interaction will be studied by comparison of ability to induce neurite outgrowth with transfection HBNF only or co-transfection of HBNF and zSyn-3 (wt or dS3) will also be investigated.
(2) In the experiment of loss-of-function, morpholino oligonucleotide (MO) technology will be applied. Through coinjection of zSyn-3 MO and HBNF/HuC-GFP, the effect of knockdown syndecan-3 expression on HBNF-induce of neurite outgrowth will be investigated.
(3) Previously, we demonstrated that HBNF could induce neurite outgrowth in zebrafish embryos. The HBNF-induced neurite outgrowth will be further studied by coinjection of zSyn-3 under the control of different promoters, such as CMV, glia-specific GFAP promoter and neuron-specific HuC promoter.
(4) We will expression of cytoplasmical truncated form of syndecan-3 (dS3) under the control of different promoters, such as CMV, glia-specific GFAP promoter and neuron-specific HuC promoter to analysis syndecan-3 (zSyn-3) function during zebrafish development.
Introduction:
The heparan sulfate proteoglycans (HSPGs) play the important roles in a variety of biologic processes such as early embryonic patterning, morphogenesis, and disease.( Forsberg and Kjellen, 2001; Nybakken and Perrimon, 2002; Selleck, 2001). The biological function of HSPGs is often achieved through modulation of growth factor–mediated signaling. In Drosophila studies have shown that loss-of-function mutations in genes encoding membrane-anchored HSPGs, the glypicans dally and dally-like, and in genes encoding enzymes involved in biosynthesis of HSPGs compromise signaling pathways mediated by growth factors such as Wingless (Wg), Decapentaplegic (Dpp), Fibroblast growth factor (FGF), and Hedgehog (Hh).( Nybakken and Perrimon, 2002; Lin and Perrimon, 1999; Tsuda et al., 1999; Baeg, et al., 2001). In vitro studies have also demonstrated the requirement of syndecan-4 in FGF-2–induced biologic response (Horowitz, et al., 2002) and dally-like in the activation of a Hedgehog - mediated signaling pathway ( Lum, et al., 2003).
Syndecan family
The syndecan family of transmembrane proteins comprises a major class of cell surface heparan sulfate proteoglycans (HSPGs). Currently, four members have been identified in mammals:
syndecan-1, syndecan-2 (fibroglycan), syndecan-3 (N-syndecan), and syndecan-4 (ryudecan)(Bernfield et al., 1992; David, 1993; Carey, 1997). The extracellular functions of syndecans are primarily mediated by their heparan sulfate (HS) or glycosaminoglycan (GAG) side chains, which have affinity for a wide variety of secreted molecules and extracellular matrix (ECM) components.(Bernfield et al., 1992; David, 1993; Schlessinger et al., 1995; Couchman and Woods, 1996; Carey, 1997;). HSPG are known to regulate cell adhesion, cell migration, differentiation, and growth factor signaling (Bernfield, 1999; Sanderson, 2001; Yamaguchi, 2001). They are abundant on most cell surfaces and are involved in a wide range of cell–cell and cell–matrix interactions.
In the nervous system, HSPGs are involved in neurogenesis, neurite guidance, synaptogenesis, and synaptic plasticity (Yamaguchi, 2001). The syndecan-1 was expressed in the developing rat neural tube (Nakanishi et al.,1997). The syndecan-4 was expressed in the neuroepithelium and its expression was much lower than syndecan-1. Syndecan-2 and syndecan-3 show contrasting expression profiles during development and segregated distribution within neurons. Syndecan-3 is more highly expressed in developing brain and concentrated in axons.
Syndecan-2 is more strongly expressed in mature brain and localized in synapses (Hsueh and Sheng, 1999; Yamaguchi, 2001). Syndecan-3 is expressed at higher levels when compared with syndecan-2 and is found on neuronal processes throughout the neural tube. The syndecan-3 have shown that this HSPG is most prominently expressed in differentiated regions in the developing mouse and chick brain (Carey et al., 1992; Gould et al., 1995; Kinnunen et al., 1998; Hsueh and Sheng, 1999). But, not much is known about which syndecans are specifically expressed in neurons, how they are distributed at the subcellular level, and how their expression patterns are
regulated during development of the nervous system.
Syndecan-3 (N-syndecan) is one of the four mammalian syndecans and it is mainly expressed in the nervous system, especially during development (Carey et al., 1997). Syndecan-3 has one transmembrane domain, a short cytoplasmic tail of 34 amino acids, and an extracellular domain that carries heparan sulfate chains (Carey et al., 1997). Syndecan-3 heparan sulfate chains bind to fibroblast growth factor-2 (FGF-2) (Chernousov and Carey, 1993) and heparin-binding growth-associated molecule (HB-GAM) (Raulo et al., 1994). Syndecans have been suggested to function as coreceptors with other signaling receptors, such as FGF receptors and integrins. It has been suggested to function in cell adhesion, neurite guidance, and cell migration during development of the nervous system (Raulo et al., 1994). Syndecan-3 has also been implicated in the regulation of synaptic plasticity in the hippocampus (Lauri et al., 1999). Syndecan-3 is expressed in an activity-dependent manner in the CA1 pyramidal neurons, and application of exogenous syndecan inhibits the induction of longterm potentiation (LTP) (Lauri et al., 1999).
Heparin binding neurotrophic factor/Pleiotrophin (HBNF/PTN)
Heparin binding neurotrophic factor/Pleiotrophin (HBNF/PTN) HBNF/PTN was first isolated as a heparin-binding protein that was eluted from a heparin affinity column with high salt concentration (Rauvala, 1989). Such a high affinity-binding property suggests that heparin or heparin-type carbohydrates may play important roles in the biological function of HBNF/PTN.
Indeed, further studies demonstrate that a transmembrane heparan sulfate proteoglycan, N-syndecan (syndecan-3), acts as a receptor for HBNF/PTN (Merenmies and Rauvala,1990).
Heparin-binding neurotrophic factor or neurite-promoting factor (HBNF) was first co-purified with bovine acidic fibroblast growth factor from brain tissues (Rauvala, 1989). It is a secretory heparin-binding protein with highly basic and cysteine-rich amino acid residues. In addition to neurite outgrowth promotion, HBNF also has a variety of biological activities, such as stimulating cell growth, acting as an angiogenesis factor, and containing oncogenic activity (Deuel, et al, 2002). Therefore, it is also known as pleiotrophin (PTN) (Li, et al., 1990) and heparin-binding growth-associated molecule (HB-GAM) (Merenmies and Rauvala, 1990). It has been proposed that HBNF/PTN is evolutionally conserved according to the published sequences from various species (Bohlen and Kovesdi, 1991). In our previous study, we established an in vivo neurite outgrowth assay in zebrafish embryos that provided a direct observation of HBNF-induced neurite outgrowth from GFP-labeled neurons during zebrafish development (Chang, et al., 2004).
Zebrafish
Zebrafish is a good model organism for the study of vertebrate development (Penberthy et al., 2002; Rubinstein, 2003). The embryos develop outside the mother and are optically transparent, allowing direct observation of their embryonic development that takes only 48 hours for completion at 280C. We have cloned several zebrafish tissue-specific promoters including pancreatic-, neuron-, and muscle-specific promoters. Their tissue specificies of gene expression were confirmed by expression of GFP in zebrafish embryos. Therefore, these tissue-specific promoters could be used to drive GFP or RFP expression in zebrafish embryos. In general, it is common to investigate the function of known or novel genes by gain-of-function and loss-of-function in zebrafish. To achieve gain-of-function, genes of interest are driven by tissue-specific promoters and injected into one-cell
zebrafish embryos (Gong et al., 2001). Alternatively, the expression constructs under the control of either ubiquitous or tissue-specific promoter were co-injected with tissue-specific promoter/GFP construct. On the other hand, to achieve loss-of-function, genes of interest are knock-downed by injection of morpholino antisense-oligonucleotides (MAO) or co-injection of MAO with tissue-specific promoter/GFP construct (Nasevicius and Ekker, 2000; Urtishak et al., 2003). The suitable transgenic GFP/RFP zebrafishes also can be used to inject MAO or expression constructs, respectively.
However, the complete zebrafish syndecan-3 transcript has not been reported. In this study, we will clone the full-length cDNA of zebrafish syndecan-3 mRNA, express the zsyn-3 gene and determine its genomic structure. The biological activity of zSyn-3 will assay in PC12 cells as well as further analyze the interaction of HBNF/PTN and zSyn-3 in zebrafish embryo.
In our previous study, we established an in vivo neurite outgrowth assay in zebrafish embryos that provided a direct observation of HBNF-induced neurite outgrowth from GFP-labeled neurons during zebrafish development. Therefore, in this proposal, we will analysis zSyn-3 function during zebrafish development. Specific aims are listed below:
(1) Co-transfection of HBNF and wild-type or cytoplasmic truncated form of zSyn-3 (dS3) into PC12 cells, cellular co-localization and ligand-receptor interaction will be studied by comparison of ability to induce neurite outgrowth with transfection HBNF only or co-transfection of HBNF and zSyn-3 (wt or dS3) will also be investigated.
(2) In the experiment of loss-of-function, morpholino oligonucleotide (MO) technology will be applied. Through coinjection of zSyn-3 MO and HBNF/HuC-GFP, the effect of knock-down syndecan-3 expression on HBNF-induce of neurite outgrowth will be investigated.
(3) Previously, we demonstrated that HBNF could induce neurite outgrowth in zebrafish embryos. The HBNF-induced neurite outgrowth will be further studied by coinjection of zSyn-3 under the control of different promoters, such as CMV, glia-specific GFAP promoter and neuron-specific HuC promoter.
(4) We will expression of cytoplasmic truncated form of syndecan-3 (dS3) under the control of different promoters, such as CMV, glia-specific GFAP promoter and neuron-specific HuC promoter to analysis syndecan-3 (zSyn-3) function during zebrafish development.
References:
Baeg GH, Lin X, Khare N, Baumgartner S, Perrimon N. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development. 2001;128:87-94.
Bernfield M, Go¨ tte M, Woo Park P, Reizes O, Fitzgerald ML, Lincecum J, and Zako M (1999).
Functions of cell-surface heparin sulfate proteoglycans. Annu. Rev. Biochem. (68): 729–777.
Bernfield M, Hinkes MT, and Gallo RL (1993) Developmental expression ofthe syndecans:
possible function and regulation. Dev Suppl (1993):205–212.
Bernfield M, Kokenyesi R, Kato M, Hinkes MT, Spring J, Gallo RL, and Lose EJ (1992) Biology of the syndecans: a family of transmembrane heparansulfate proteoglycans. Annu Rev Cell Biol (8):365–393.
Bohlen P, and Kovesdi I (1991) HBNF and MK, members of a novel gene family of heparin binding proteins with potential roles in embryogenesis and brain function, Prog. Growth Factor Res. (3): 143–157.
Carey D (1997) Syndecans: multifunctional cell-surface co-receptors.Biochem J 27:1–16.
Carey DJ, Conner K, Asundi VK, O’Mahoney DJ, Stahl RC, Showalter L, Cizmeci-Smith G, Hartman J, and
Rothblum LI (1997) cDNA cloning, genomic organization, and in vivo expression of rat N-syndecan. J Biol Chem (272): 2873–2879.
Carey DJ, Evans DM, Stahl RC, Asundi VK, Conner KJ, Garbes P, and Cizmeci- Smith G (1992) Molecular cloning and characterization of N-syndecan, a novel transmembrane heparan sulfate proteoglycan. J Cell Biol (117):191–201.
Chang MH, Huang CJ, Hwang SP, Lu IC, Lin CM, Kuo TF, Chou CM (2004).Zebrafish heparin-binding neurotrophic factor enhances neurite outgrowth during its development. Biochem Biophys Res Commun.(321):502-509.
Chernousov M.A., and Carey, D.J. (1993). N-syndecan (syndecan 3)from neonatal rat brain binds basic fibroblast growth factor. J. Biol.Chem.(268):16810–16814.
Couchman JR, and Woods A (1996) Syndecans, signaling, and cell adhesion.J Cell Biochem (61):578 –584.
David G (1993) Integral membrane heparan sulfate proteoglycans.FASEB J (7):1023–1030.
Deuel TF, Zhang N, Yeh HJ, Silos-Santiago I, and Wang ZY (2002) Pleiotrophin: a cytokine with diverse functions and a novel signaling pathway, Arch. Biochem. Biophys. (397): 162–171.
Forsberg E, Kjellen L. Heparan sulfate: lessons from knockout mice. J Clin Invest. 2001;108:175- 180.
Gong Z, Ju B, and Wan H, (2001) Green fluorescent protein (GFP) transgenic fish and their applications, Genetica (111): 213–225.
Horowitz A, Tkachenko E, Simons M. Fibroblast growth factor-specific modulation of cellular response by syndecan-4. J Cell Biol. 2002;157:715- 725.
Hsueh YP, Sheng M (1999) Requirement of N-terminal cysteines of PSD-95 for PSD-95 multimerization and ternary complex formation, butnot for binding to potassium channel Kv1.4. J Biol Chem (174):532–536.
Kinnunen T, Kaksonen M, Saarinen J, Kalkkinen N, Peng HB, and Rauvala H. (1998).Cortactin–Src kinase signaling pathway is involved in N-syndecan- dependent neurite outgrowth. J. Biol. Chem. (273):
10702–10708.
Lauri SE, Kaukinen S, Kinnunen T, Ylinen A, Imai S, Kaila K, Taira T, Rauvala H (1999) Reg1ulatory role and molecular interactions of acell-surface heparan sulfate proteoglycan (N-syndecan) in hippocampallong-term potentiation. J Neurosci (19):1226 –1235.
Li YS, Milner, Chauhan PG, Watson AK, Hoffman MA, Kodner RM, , Milbrandt CM, and Deuel T.F. (1990) Cloning and expression of a developmentally regulated protein that induces mitogenic and neurite outgrowth activity, Science (250):1690–1694.
Lin X, Perrimon N. Dally cooperates with Drosophila Frizzled 2 to transduce Wingless signalling. Nature.
1999;400:281-284.
Lum L, Yao S, Mozer B, et al. Identification of Hedgehog pathway components by RNAi in Drosophila cultured cells. Science. 2003;299:2039- 2045.
Merenmies, J, Rauvala, H (1990) Molecular cloning of the a 18-kDa growth- associated protein of developing brain,
J. Biol. Chem. (265):16721–16724.
Nakanishi T, Kadomatsu K, Okamoto T, Ichihara-Tanaka K, Kojima T, Saito H, Tomoda Y, Muramatsu T.
(1997) Expression of syndecan-1 and -3 during embryogenesis of the central nervous system in relation to binding with midkine. J Biochem (Tokyo) (121):197–205.
Nasevicius A, and Ekker SC.(2000) Effective targeted gene 'knockdown' in zebrafish. Nat Genet.
(26):216-20.
Nybakken K, Perrimon N. Heparan sulfate proteoglycan modulation of developmental signaling in Drosophila. Biochim Biophys Acta. 2002;1573: 280-291.
Park, HC, Kim, C.H., Bae, Y.K., Yeo, S.Y., Kim, S.H., Hong, S.K., Shin, J., Yoo, K.W., Hibi, M., Hirano, T., Miki, N., Chitnis, A.B., Huh, T.L., Analysis of upstream elements in the HuC promoter leads to the establishment of transgenic zebrafish with fluorescent neurons, Dev. Biol. 227 (2000) 279–293.
Penberthy WT, Shafizadeh E, and Lin S. (2002) The zebrafish as a model for human disease. Front Biosci.(7):1439-53.
Raulo, E., Chernousov, M.A., Carey, D.J., Nolo, R., Rauvala, H., Isolation of a neuronal cell surface receptor of heparin binding growth-associated molecule HB-GAM, J. Biol. Chem. 269 (1994) 12999–13004.
Rauvala, H. (1989) An 18-kd heparin-binding protein of developing brain that is distinct from fibroblast growth factors, EMBO J. (8): 2933–2941.
Rubinstein AL. (2003) Zebrafish: from disease modeling to drug discovery. Curr Opin Drug Discov Devel.
(6):218-23.
Sanderson, 2001; Sanderson, R. D. (2001). Heparan sulfate proteoglycans in invasion and metastasis. Semin.
Cell Dev. Biol. (12): 89–98.
Schlessinger J, Lax I, Lemmon M (1995) Regulation of growth factor activation by proteoglycans: what is the role of the low affinity receptors? Cell (83):357–360
Selleck SB. Genetic dissection of proteoglycan function in Drosophila and C. elegans. Semin Cell Dev Biol.
2001;12:127-134.
Thisse C, Thisse B, Schilling TF and Postlethwait JH. Structure of the zebrafish snail1 gene and its expression in wild-type, spadetail and no tail mutant embryos., Development. 119 (1993) 1203-1215.
Tsuda M, Kamimura K, Nakato H, et al. The cellsurface proteoglycan Dally regulates Wingless signalling in Drosophila. Nature. 1999;400:276- 280.
Urtishak KA, Choob M, Tian X, Sternheim N, Talbot WS, Wickstrom E, and Farber SA. (2003) Targeted gene knockdown in zebrafish using negatively charged peptide nucleic acid mimics. Dev Dyn.
(228):405-13.
Westerfield, M., The Zebrafish Book, third ed., University of Oregan Press, Eugene, OR, 1995.
Yamaguchi, Y. (2001). Heparan sulfate proteoglycans in the nervous system: Their diverse roles in neurogenesis, axon guidance, and synaptogenesis. Semin. Cell Dev. Biol. (12): 99–106.
Result
Expression profiles of zebrafish syndecan-3(zSyn-3) mRNA in adult tissues
In order to determine the expression pattern of zebrafish zSyn3 mRNA by RT-PCR, zebrafish embryos at various tissues from adult zebrafish were collected and used to isolate total RNA for cDNA preparation. Our data showed that the zSyn3 mRNA was expressed in brain, eye, heart, intestine and kidney (Fig.1). The expression of β-actin transcript was also determined as well and served as internal control (Fig.1).
Construction of expression clones from zebrafish cDNA
Syndecan-3 has one transmembrane domain, a short cytoplasmic tail of 34 amino acids, and an extracellular domain that carries heparan sulfate chains. We have cloned the full-length zSyn3 cDNA from zebrafish. RT-PCR analysis revealed that zebrafish zSyn3 transcript was highly expressed in adult brain, eye and intestine tissues while less in other tissues.
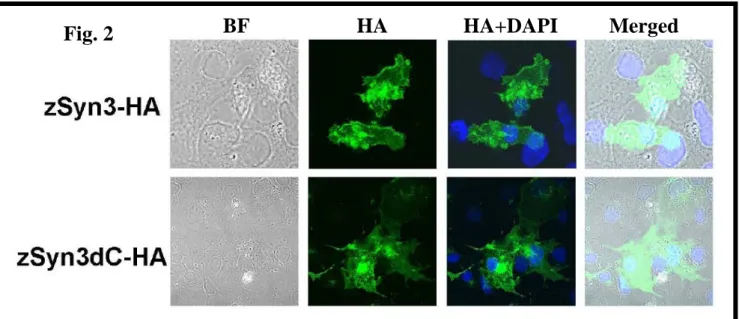
Phenotype of cells transiently transfected with Syn3
In order to study the function of the syndecan-3 protein, transient transfection experiments were performed in COS-1 cells (Fig. 2). Cells were transfected with the full-length syndecan-3 (zSyn3-HA) or with the deletion of the entire cytoplasmic domain (zSyn3dC-HA), fixed 48 h post-transfection and immunolabeled with monoclonal anti-HA antibody. Analyzed by confocal microscopy, strong membrane staining was observed in the syndecan-3 transfected cells, while the deletion of the entire cytoplasmic domain abolished the membrane staining. This result indicates that syndecan-3 located to the membrane.
Fig. 1
Fig. 2 BF HA HA+DAPI Merged
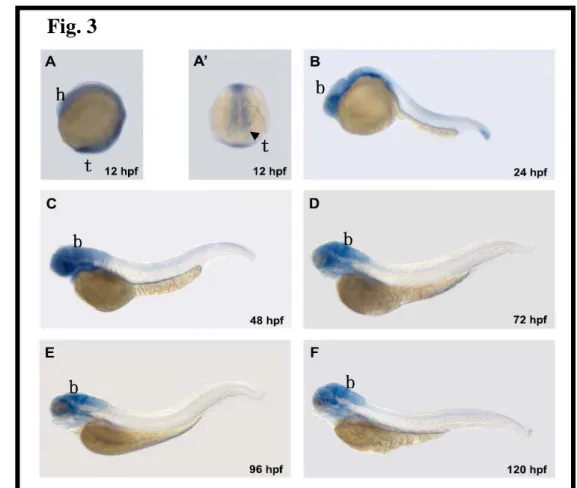
Differential expression of zSyn3 mRNA during embryogenesis
In order to elucidate the role of zebrafish Syn3 in the development of the nervous system, we examined the temporal and spatial patterns of zsyn3 expression using whole-mount in situ hybridization. In 12, 24, 48, 72, 96, and 120 hpf embryos (Fig. 3, panels A to F), zsyn3 mRNA was expressed predominantly in the developing neural structures including eye, forebrain, midbrain, and hindbrain, which is consistent with our previous data that zebrafish syn3 expressed in adult brain (Fig. 3).
Effects of zSyn3 knockdown during zebrafish development
To reveal the function of zSyn3, we prepared antisense morpholino oligonucleotides (MOs) against the translational initiation site (zSyn3-MO) of the zSyn3 gene. Injection of zSyn3-MO inhibited the translation of zSyn3 protein (Fig. 4). The injection zSyn3-MO, led to reduction or truncation of the posterior body; these effects were more severe than control (Fig.4).
Fig. 3
h
t
t
b b
b
b
b